
黄淮麦区(南片)小麦籽粒过氧化物酶活性分析及其等位基因检测
2021-12-01 09:06张星宇宋佳静宋全昊赵立尚朱统泉朱保磊陈建辉宋晓朋
麦类作物学报 2021年9期
陈 杰,张星宇,白 冬,宋佳静,宋全昊,金 艳,赵立尚,朱统泉,朱保磊,陈建辉,宋晓朋
(1.河南省驻马店市农业科学院,河南驻马店 463000;2.河南农业大学农学院,河南郑州 450002;3.河南省信阳市农业科学院,河南信阳 464000;4.河南省对外科技交流中心,河南郑州 450003)
小麦面粉的色泽是影响小麦加工品质的重要因素之一,在制作馒头、包子、面条等传统面制食品过程中,对面粉的白度有较高的要求[1]。为了满足消费者对面粉白度的要求,生产商通常会在面粉中加入过氧化苯甲酰等增白剂以提高面粉的白度。但长期食用含增白剂的食品对身体健康会产生不利的影响[2]。因此,通过小麦品质色泽改良来提升面粉及其面制食品的白度对保障我国食品安全具有重要意义。Hemalatha等[3]和Gelinas等[4]研究表明,小麦籽粒中过氧化物酶(peroxidase,POD)对面粉及其面制品的色泽具有增白作用。因此,研究小麦籽粒中的POD活性及其基因型有利于小麦品质色泽的改良。
小麦籽粒的POD活性属于数量性状,受生态环境、栽培方式、遗传等多重因素的影响,但遗传因素的效应更大[5-6]。小麦第3和第7同源染色体组上存在调控POD活性的主效基因位点[7-10]。魏景欣等[9]以豆麦/石4185重组自交系群体的214个株系为材料,利用90K SNP芯片对小麦籽粒POD活性进行全基因组关联分析,结果在3AL染色体上检测到主效QTL位点,可以解释5.3%~9.2%的表型变异。时 佳等[10]以黄淮麦区151份及北部冬麦区82份小麦品种(系)为材料,利用90K SNP 芯片对小麦籽粒POD活性进行全基因组关联分析,在7DS染色体上检测到主效QTL位点,可以解释18.0%~21.4%的表型变异。Wei等[11]利用同源克隆的方法克隆出位于3AL染色体上的TaPod-A1基因,并开发设计了两对共显性互补功能标记POD-3A1和POD-3A2,用于鉴定等位基因TaPod-A1a和TaPod-A1b。Geng等[12]利用同源克隆的方法克隆出位于7DS染色体上的TaPod-D1基因,并开发设计了两对共显性互补功能标记POD-7D1和POD-7D6,用来鉴定等位基因TaPod-D1a和TaPod-D1b。上述分子功能标记已经在新疆冬小麦和春小麦里面进行了应用[13-15]。
黄淮麦区(南片)包含陕西省关中平原地区、河南省信阳以北地区以及安徽省和江苏省淮河以北地区,是我国重要的小麦主产区之一。改良本麦区小麦的色泽对提升我国小麦的加工品质有重要意义。与黄色素含量、多酚氧化酶(polyphenol oxidase,PPO)活性和POD活性相关的基因均能影响小麦的色泽品质,有关黄色素含量和PPO基因在本麦区品种(系)中的分布已有了深入的研究,但是有关POD基因在本麦区的研究鲜有报道。因此,本研究以黄淮麦区(南片)新育成的94份小麦品种(系)为试验材料,利用分光光度计对试验材料的POD活性进行测定,同时利用功能标记POD-3A1、POD-3A2、POD-7D1和POD-7D6对试验材料TaPod-A1和TaPod-D1位点的等位基因进行检测,研究TaPOD-A1和TaPOD-D1等位基因在本麦区新育成小麦品系中的分布情况及其与POD活性之间的关系,以期为黄淮麦区(南片)小麦品质色泽改良提供参考信息。
1 材料与方法
1.1 供试材料
供试材料为2020-2021年度国家黄淮麦区(南片)区域试验的全部参试品种(系),共94份(表1)。其中来自陕西的有12份,河南的有52份,安徽的有16份,江苏的有10份,北京的有4份。这些材料基本代表了黄淮麦区(南片)当前的小麦育种和生产需求现状。

表1 试验材料的POD活性及其基因等位变异类型Table 1 POD activity and allele of POD gene in wheat cultivars
1.2 POD活性测定
利用分光光度计测定供试材料的POD活性,具体步骤参照魏景欣等[9]的方法进行。
1.3 等位基因检测
供试材料籽粒基因组DNA的提取参照陈 杰等[16]的方法。用POD-3A1和POD-3A2两对共显性互补标记对供试材料3A染色体上TaPod-A1位点的等位基因进行检测,如果扩增出291 bp的片段,则为TaPod-A1a等位基因;如果扩增出766 bp的片段,则为TaPod-A1b等位基因;用POD-7D1和POD-7D6两对共显性互补标记对供试材料7D染色体上TaPod-D1位点的等位基因进行检测,如果扩增出540 bp的片段,则为TaPod-D1a等位基因;如果扩增出640 bp的片段,则为TaPod-D1b等位基因。所选分子标记的具体信息详见表2。PCR反应体系和扩增程序均参照陈 杰等[16]的方法进行。PCR扩增产物经琼脂糖凝胶电泳分离后,用Alpha凝胶系统扫描成像。
1.4 数据分析
用Microsoft Excel 2003计算分布频率,用SPSS 18.0软件进行方差分析。
2 结果与分析
2.1 供试材料的POD活性
利用分光光度计对94份供试材料进行POD活性测定。POD活性总体平均值为668.6 U·g-1·min-1,标准差为110.5,变幅为431.3~954.8 U·g-1·min-1。POD活性小于600 U·g-1·min-1的材料有28份,占比29.8%;POD活性为600~700 U·g-1·min-1的材料有35份,占比37.2%;POD活性大于700 U·g-1·min-1的材料有31份,占比33.0%。其中,有4份材料(郑麦163、德研1658、益科麦17118和阜麦13)的POD活性小于500 U·g-1·min-1;有4份材料(西农1366、职院171、平安12号和普冰03)的POD活性高于900 U·g-1·min-1。说明黄淮麦区(南片)大部分新育成小麦材料的POD活性为中等水平。结合表1进一步分析可知,来自陕西、河南、安徽、江苏的供试材料均以中等POD活性的材料分布为主。
2.2 TaPod-A1和 TaPod-D1位点等位基因的鉴定结果
利用POD-3A1和POD-3A2两对共显性互补标记对94份供试材料的TaPod-A1位点进行等位基因检测。结果显示,徐麦15019、泛麦27、郑研麦182等46份材料可扩增出291 bp的目的条带,说明这些材料含有TaPod-A1a等位基因,分布频率为48.9%;西农172、富麦916、憨丰3468等48份材料能扩增出766 bp的目的条带,说明这些材料含有TaPod-A1b等位基因,分布频率为51.1%。部分材料的扩增结果见图1。

a:用标记POD-3A1扩增的结果;b:用标记POD-3A2扩增的结果;M:DNA marker;1:西农172;2:富麦916;3:徐麦15019;4:泛麦27;5:憨丰3468;6:职院171;7:郑研麦182;8:郑大181;9:许科10号;10:安科1605;11:百农5819;12:德研1658。图2同。a:Detection results amplified by POD-3Al;b:Detection results amplified by POD-3A2;M:DNA marker;1:Xinong 172;2:Fumai 916;3:Xumai 15019;4:Fanmai 27;5:Hanfeng 3468;6:Zhiyuan 171;7:Zhengyanmai 182;8:Zhengda 181;9:Xuke 10;10:Anke 1605;11:Bainong 5819;12:Deyan 1658.The same in figure 2.图1 POD-3A1和POD-3A2标记对部分材料的检测结果Fig.1 Detection results of some test materials amplified by POD-3Al and POD-3A2
用POD-7D1和POD-7D6两对共显性互补标记对94份供试材料的TaPod-D1位点进行等位基因检测,结果显示,94份材料均能扩增出640 bp的目的片段,部分材料的扩增结果见图2,说明所有材料均含有TaPod-D1b等位基因。表明供试材料TaPod-D1位点的多态性较为单一。

图2 POD-7D6标记对部分材料的检测结果Fig.2 Detection results of some test materials amplified by POD-7D6
2.3 TaPod-A1和 TaPod-D1位点等位基因与POD活性的关系
从表3可知,在TaPod-A1位点,共检测到TaPod-A1a和TaPod-A1b两种等位基因,携带TaPod-A1a等位基因材料的POD活性平均值为614.4 U·g-1·min-1,携带TaPod-A1b等位基因材料的POD活性平均值为720.6 U·g-1·min-1,且两者之间的差异达显著水平,这表明等位基因TaPod-A1a和TaPod-A1b分别与低POD、高POD活性相关。在TaPod-D1位点,只检测到TaPod-D1b等位基因,携带此等位基因材料的POD活性平均值为668.6 U·g-1·min-1,变幅为431.3~954.8 U·g-1·min-1。
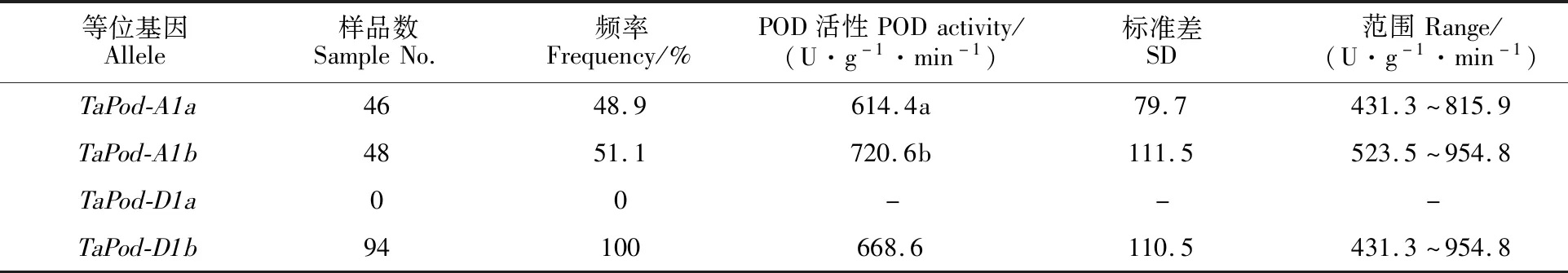
表3 TaPod-A1和 TaPod-D1位点等位基因对POD活性的影响Table 3 Effect of TaPod-A1 and TaPod-D1 loci on POD activity
2.4 TaPod-A1和 TaPod-D1位点等位基因在不同省份供试材料中的分布频率
从表4可知,与低POD活性相关的TaPod-A1a等位基因在黄淮麦区(南片)各省份供试材料的分布频率高低依次为安徽(62.5%)>河南(51.9%)>陕西(41.7%)>江苏(30.0%)>北京 (25.0%);与高POD活性相关的TaPod-A1b等位基因在各省份供试材料的频率高低则相反。说明TaPod-A1b优异等位基因在黄淮麦区(南片)不同省份间的分布频率差异很大,在北京、陕西和江苏的分布频率均高于TaPod-A1a等位基因,在安徽和河南的分布频率均低于TaPod-A1a等位基因。

表4 TaPod-A1 和 TaPod-D1位点等位基因在不同省份材料中的分布频率Table 4 Distribution frequency of TaPod-A1 and TaPod-D1 loci in materials from different provinces
3 讨 论
单子龙等[17]利用KASP标记对153个河北小麦品种的TaPod-A1位点进行等位基因检测,发现优异等位基因TaPod-A1b的分布频率为 25.49%。耿洪伟等[13]利用标记POD-3A1和POD-3A2对129份新疆小麦品种的TaPod-A1位点进行等位基因检测,发现TaPod-A1b等位基因的分布频率为39.5%。本研究结果表明,TaPod-A1b等位基因在黄淮麦区(南片)新育成的94份小麦品种(系)中的分布频率为51.1%,在黄淮麦区(南片)组成省份之间的分布情况为江苏(70.0%)>陕西(58.3%)>河南(48.1%)>安徽 (37.5%)。从以上数据可以看出,TaPod-A1b等位基因在我国不同省份间的分布比例差异很大,在江苏和陕西的分布频率均高于50%,在河南、安徽的分布频率均低于50%。因此,今后应当对TaPod-A1位点上的TaPod-A1b等位基因施加选择压力,淘汰综合农艺性状差且携带TaPod-A1a等位基因的材料,选择综合农艺性状好且携带TaPod-A1b等位基因的材料,以促进高POD活性小麦品种的选育。
谢 磊等[14]和王丽丽等[15]利用标记POD-7D1和POD-7D6分别对新疆小麦TaPod-D1位点等位基因进行检测,发现TaPod-D1a等位基因的分布频率分别为42.9%和53.1%。然而,本研究在黄淮麦区(南片)新育成的94份小麦品种(系)的TaPod-D1位点上没有检测到TaPod-D1a等位基因。出现这种状况的原因可能有两个,一是黄淮麦区小麦品种遗传背景较近,品种趋于同质化[18];二是本麦区的育种人员致力于产量和抗性性状的改良,对品种的POD活性选择没有引起足够重视。下一步可以跨麦区扩大材料交流数量,进行TaPod-D1位点等位基因的鉴定与发掘,并通过不同等位基因和表型的关联分析,明确各等位基因与表型之间的关系,为POD活性基因分子辅助育种提供理论依据。
本研究通过对94份供试小麦材料POD活性的表型和基因型分析可知,TaPod-A1位点上TaPod-A1a和TaPod-A1b等位基因之间的活性差异达到显著水平。这与Wei等[11]和王丽丽等[15]的研究结果一致,验证了等位基因TaPod-A1b与高POD活性相关,等位基因TaPod-A1a与低POD活性相关,同时也验证了标记POD-3A1和POD-3A2作为分子辅助育种工具的有效性。另外,本研究通过表型和基因型筛选出来的西农1366、职院171、普冰03和平安12号不仅POD活性高于900 U·g-1·min-1,而且均携带与高POD活性相关的TaPod-A1b等位基因,可作为选育高POD活性品种的亲本资源,在组配杂交组合中进行应用。
猜你喜欢
河南农业·综合版(2022年2期)2022-03-18
河南农业(2022年2期)2022-03-14
作物学报(2022年3期)2022-01-22
河南农业·综合版(2021年7期)2021-08-23
智慧健康(2021年17期)2021-07-30
河南农业(2021年7期)2021-07-30
校园英语·月末(2020年6期)2020-09-14
新课程·下旬(2018年9期)2018-11-14
安徽农学通报(2016年20期)2016-12-26
青少年科技博览(中学版)(2015年10期)2015-01-11
