
小麦水分利用效率对CO2浓度响应的模型研究
2021-12-01 09:06康华靖王复标叶子飘
麦类作物学报 2021年9期
康华靖,楼 珏,王复标,安 婷,叶子飘
(1.温州市农业科学研究院,浙江温州 325006;2.浙南作物育种重点实验室,浙江温州 325006;3.井冈山大学 数理学院生物物理研究所,江西吉安 343009)
水对陆生植物的热量调节和能量代谢具有极其重要意义[1-2]。水分利用效率(water use efficiency,WUE)是指植物消耗单位水量生产出的同化量[3-4]。在不同的学科和范畴,对水分利用效率的理解和定义不尽相同[5]。在叶片尺度上,水分利用效率主要指内禀水分利用效率(intrinsic water-use efficiency,WUEi)和瞬时水分利用效率(instantaneous water-use efficiency,WUEinst),前者被定义为叶片净光合速率(net photosynthetic rate,Pn)与气孔导度(stomatal conductance,Gs)之比[6-9],用于评估基因对植物的影响[7];后者被定义为叶片的Pn与蒸腾速率(transpiration rate,Tr)之比[6],主要用于估算环境因素对植物的影响[7]。
在全球CO2浓度快速升高的背景下,植物对CO2浓度升高的响应早已成为研究热点。同样,植物叶片的WUEi和WUEinst对大气CO2浓度(atmospheric CO2concentration,Ca)的响应曲线的研究也逐渐被重视[10-12]。有学者定性地研究植物叶片的WUEinst对Ca的响应[13-14];也有学者用二次方程[15-16]、指数方程[17]、对数函数[10]定量描述植物叶片的WUEinst对Ca的响应。然而,由以上这些模型得到的拟合值与观测值之间存在较大的偏差,且用这些模型预测的WUEi和WUEinst随CO2浓度的增加而一直增加;另外,用指数方程或对数函数不能得到与最大内禀水分利用效率(WUEi-max)或最大瞬时水分利用效率(WUEinst-max)相对应的饱和CO2浓度。此外,王建林等[18]构建的模型虽能同时描述WUE对光和CO2的响应关系,但由于该模型较为复杂,涉及的参数较多,导致其应用相对困难;同时,由于该模型也是一个没有极点的函数,同样也不能用该模型得到与WUEi-max或WUEinst-max相对应的饱和CO2浓度。由此可知,目前用于拟合植物叶片WUEi和WUEinst对CO2响应曲线的模型存在难以克服的缺陷,尚无一个普适性模型能够准确描述植物叶片的WUEi和WUEinst对CO2响应曲线。而未来气候变暖和CO2浓度升高等全球气候变化正在通过影响C积累量和耗水量来显著影响植物的WUE[19-22]。因此,构建一个植物叶片WUE对CO2响应的普适性模型对定量描述植物叶片在当前和未来气候条件下WUE对CO2响应的变化规律尤为必要。
为此,本研究首先在植物光合作用对CO2的响应新模型的基础上构建WUEi和WUEinst对CO2的响应模型;其次,测量典型的C3作物——小麦(TriticumaestivumL.)在光合有效辐射强度(PAR)为2 000 μmol·m-2·s-1时的CO2响应曲线,然后运用新构建的WUEi和WUEinst对CO2(包括大气CO2浓度(Ca)和胞间CO2浓度(intercellular CO2concentration,Ci))的响应模型进行拟合,比较拟合结果与观测数据之间的差异,以判断新构建的模型是否合理;同时,也比较分析WUEi和WUEinst对Ca和Ci的响应是否存在差异,以期为准确定量研究WUE对CO2响应提供一个有效的数据工具。
1 模型构建
1.1 WUEi对CO2的响应模型
Pn对CO2的响应可由叶子飘[23]构建的模型进行描述:
(1)
式中,α为Pn对CO2的响应曲线的初始斜率;β和γ分别为与CO2浓度无关的2个系数;Rp是植物叶片的光下呼吸速率;C为CO2浓度,它既可以表示大气CO2浓度(Ca),也可以表示为胞间CO2浓度(Ci)。因此,式(1)既可以描述植物叶片的Pn对Ca的响应曲线(response curve of photosynthesis to atmospheric CO2concentration,Pn-Ca),也可以描述Pn对Ci的响应曲线(response curve of photosynthesis to intercellular CO2concentration,Pn-Ci)。由式(1)拟合可以得到植物叶片的最大羧化速率(maximum carboxylation rate,Pnmax)、饱和CO2浓度(saturation CO2concentration,Csat)、CO2补偿点(CO2compensation point,Γ)和Rp等。
由式(1)并结合WUEi=Pn/Gs可得到WUEi对CO2的响应模型:
(2)
式中的αi为WUEi对CO2响应曲线的初始斜率;βi和γi分别为与CO2浓度无关的2个系数;Ki=Rp/Gs,为CO2浓度等于0 μmol·mol-1时,植物叶片的Rp与Gs之比值。同样地,式(2)既可以描述植物叶片的WUEi对Ca的响应曲线(response curve of WUEito atmospheric CO2concentration,WUEi-Ca),也可以用于拟合WUEi对Ci的响应曲线(response curve of WUEito intercellular CO2concentration,WUEi-Ci)。
由于式(2)是一个有极点的函数,因此可以由式(2)得到最大WUEi(maximum WUEi,WUEi-max)及与其相对应的饱和CO2浓度(saturated CO2concentration,Cint-sat):
(3)
把(3)式代入(2)式,那么WUEi-max可以由下式得到:
(4)
1.2 WUEinst对CO2的响应模型
由式(1)并结合WUEinst=Pn/Tr可得到WUEinst对CO2的响应模型:
(5)
式中的αinst为WUEinst对CO2的响应曲线的初始斜率;βinst和γinst分别为与CO2浓度无关的2个系数;Kinst=Rp/Tr,为CO2浓度等于0 μmol·mol-1时植物叶片的Rp与Tr之比值,该值被定义为零点瞬时水分利用效率。同样地,式(5)既可以描述植物叶片的WUEinst对Ca的响应曲线(response curve of WUEinstto atmospheric CO2concentration,WUEinst-Ca),也可以用于研究WUEinst对Ci的响应曲线(response curve of WUEinstto intercellular CO2concentration,WUEinst-Ci)。
类似地,可以由式(5)得到最大WUEinst(maximum WUEinst,WUEinst-max)及其相对应的饱和CO2浓度(saturation CO2concentration,Cinst-sat):
(6)
把式(6)代入式(5),那么WUEinst-max可以由下式得到:
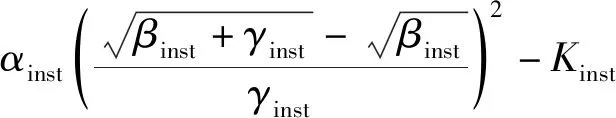
(7)
1.3 材料培养及数据测量
本试验在中国科学院禹城综合试验站进行。该试验站位于山东省德州市下辖的禹城市境内。禹城市地处山东省西北部,境内皆平原,地势南高北低,西高东低;属暖温带大陆季风气候,四季分明,干湿季节明显,光照充足。小麦供试品种为济麦22。2012年10月播种,大田常规管理。数据测量于2013年5月14-16日的晴天进行。此时小麦处于盛花期。随机选5~7株长势较为一致的健壮植株,以其旗叶作为测量对象。此时旗叶已完全展开,叶色深绿,叶形饱满。利用便携式光合仪(LI-6400-40,LI-COR INC.,USA)于晴天上午9:00-11:30进行数据测量。数据测量前进行开机预热及各项检查。在自然光下诱导1.5~2 h后,采用开放式气路,流速为400 μmol·s-1。叶室温度约为29.47 ℃(大气温度约为30.48 ℃左右),空气相对湿度为45%~70%。根据前期光响应曲线测量及拟合结果得知,小麦叶片的饱和光强约为2 000 μmol·m-2·s-1。因此,测量时光合有效辐射强度(PAR)设置为2 000 μmol·m-2·s-1。应用CO2注入系统提供不同的外界CO2梯度(Ca):1 400、1 200、1 000、800、600、380、200、150、100、80、50和0 μmol·mol-1。应用自动测量程序进行数据测量。每次程序记录的最小等待时间设置为2 min,最大等待时间设置为3 min。数据记录前仪器均自动进行参比室和样品室之间的匹配。
1.4 数据处理
对于Pn-Ca曲线和Pn-Ci曲线,把用内插法得到的CO2补偿点作为观测值;对于Pn-Ci曲线,把用外推法得到小麦的Rp作为观测值[24]。其他光合参数则用“光合模型拟合软件(PMSS)”http://photosynthetic.sinaapp.com/calc.html(井冈山大学数理学院)中有关模型对Pn-Ca(Pn-Ci)、WUEi-Ca(WUEi-Ci)和WUEinst-Ca(WUEinst-Ci)曲线进行拟合,并获得相应的光合参数。数据分析均取5个重复(平均值±标准误差,n=5),用SPSS18.5软件(SPSS,Chicago,IL,United States)对拟合结果与观测值之间进行方差分析(analysis of variance,ANOVA)。作图由Origin8.5软件完成,最后用Adobe Illustrator CS5进行图形处理。此外,本研究中用确定系数(determination coefficient,r2=1-SSE/SST,其中SST和SSE分别为平方总和与误差平方和)评价模型拟合程度。
2 结果和分析
2.1 光合作用对CO2的响应
图1为小麦的Pn-Ca(左)和Pn-Ci(右)曲线。由图1可知,在较低CO2浓度(≤400 μmol·mol-1)下,随Ca或Ci的增加,Pn快速上升;CO2浓度超过400 μmol·mol-1后增加速度逐渐放缓,Pn逐渐达到最大值,即出现磷酸丙糖利用(TPU)的限制阶段。对比图1的Pn-Ca(左)和Pn-Ci(右)曲线,可以发现两者的变化趋势极为相似。而Pn-Ci(右)曲线的初始斜率是Pn-Ca(左)曲线初始斜率的2.16倍(表1)。由于Pn-Ci曲线的初始斜率较大(表1),因而基于该曲线用外推法得到的Rp值也较大(表1)。此外,由图1可知,叶子飘[23]构建的Pn-Ca或Pn-Ci模型均可较好地拟合小麦的Pn-Ca(左)和Pn-Ci(右)曲线,确定系数(r2)分别高达0.999 9和0.999 6。

图1 小麦光合作用对大气和胞间CO2浓度的响应曲线Fig.1 Response curve of wheat photosynthesis to atmospheric and intercellular CO2 concentration
通过式(1)分别拟合小麦的Pn-Ca和Pn-Ci曲线,进而得到的主要光合参数值(表1)。在这些参数中,拟合Pn-Ca曲线得到的初始羧化效率(α)、Rp以及Csat与拟合Pn-Ci曲线得到结果均存在显著差异,其余光合参数(Pnmax和Γ)则不存在显著差异。用式(1)拟合Pn-Ca或Pn-Ci曲线得到的Pnmax、Csat、Γ和Rp拟合值与其观测值之间均无显著差异。进一步说明由叶子飘[23]构建的模型可很好地拟合Pn-Ca或Pn-Ci曲线。

表1 由式(1)拟合小麦叶片的Pn-Ca和Pn-Ci曲线得到的光合参数值及其对应的观测值(n =5)Table 1 Observed values and fitted values of photosynthesis indices by Eqn.(1)for Pn-Caand for Pn-Ci curves of T.aestivum(n=5)
2.2 内禀水分利用效率对CO2的响应
从图2中的WUEi-Ca曲线来看,当外界CO2浓度(Ca)低于800 μmol·mol-1时,WUEi随Ca的增加而直线上升;Ca达到800 μmol·mol-1后,WUEi随Ca的增加而缓慢增加,直到最大值;当Ca超过1 200 μmol·mol-1后,WUEi则随Ca的增加而下降。对于WUEi-Ci曲线,当胞间CO2浓度(Ci)低于530 μmol·mol-1时,WUEi随Ci的增加而直线上升;Ci达到530 μmol·mol-1后,WUEi随Ci的增加而缓慢增加直到最大值;当Ci大于890 μmol·mol-1后,WUEi则同样随Ci的增加出现下降趋势。利用式(2)可较好地拟合小麦的WUEi-Ca(左)和WUEi-Ci(右)曲线,r2分别高达0.998 9和 0.997 9。

图2 小麦叶片的内禀水分利用效率对大气和胞间CO2浓度的响应曲线Fig.2 Response curve of intrinsic water use efficiency in wheat leaf to atmospheric and intercellular CO2 concentration
由表2可知,由式(2)拟合WUEi-Ca和WUEi-Ci曲线得到的初始斜率之间存在显著差异,且后者是前者的2.10倍;同样,饱和CO2浓度的两个拟合值之间也存在显著差异。而WUEi-max的两个拟合值间及其拟合值与对应观测值之间均不存在显著差异。另外,饱和CO2浓度的两个拟合值间及其拟合值与对应观测值之间也均不存在显著差异。

表2 由式(2)拟合小麦叶片的WUEi-Ca和WUEi-Ci曲线得到的参数值及其对应的观测值(n=5)Table 2 Observed values and fitted values of indices by Eqn.(2)for WUEi-Ca and for WUEi-Ci curves of T.aestivum(n=5)
2.3 瞬时水分利用效率对CO2的响应
小麦瞬时水分利用效率对CO2的响应规律类似于内禀水分利用效率(图3)。对于WUEinst-Ca曲线(左),当Ca低于800 μmol·mol-1时,WUEinst随Ca的增加而直线上升;Ca高于800 μmol·mol-1后,WUEinst随Ca的增加而缓慢增加,直到最大值;当Ca超过1 200 μmol·mol-1后,WUEi随Ca的增加而出现下降;对于WUEinst-Ci曲线(右),在Ci低于530 μmol·mol-1的条件下,WUEinst随Ci的增加而直线上升;当Ci高于530 μmol·mol-1后,WUEinst随Ci的增加而缓慢增加,直到最大值;当Ci超过890 μmol·mol-1后WUEinst随Ci的增加而出现下降。此外,用式(5)可较好地拟合小麦的WUEinst-Ca(左)和WUEinst-Ci(右)曲线,r2分别高达0.999 2和 0.998 3。

图3 小麦叶片的瞬时水分利用效率对大气和胞间CO2浓度的响应曲线Fig.3 Response curve of instantaneous water use efficiency in wheat leaf to atmospheric and intercellular CO2 concentration
由表3可知,由式(5)拟合WUEinst-Ca和WUEinst-Ci曲线得到的初始斜率之间存在显著差异,且后者是前者的2.08倍;同样,饱和CO2浓度的两个拟合值之间也存在显著差异。而WUEinst-max的两个拟合值间及其拟合值与对应观测值之间均不存在显著差异;同样,饱和CO2浓度的两个拟合值间及其拟合值与对应观测值之间也均不存在显著差异(表3)。

表3 由式(5)拟合小麦叶片的WUEinst-Ca和WUEinst-Ci曲线得到的参数值及其对应的观测值(n=5)Table 3 Observed values and fitted values of indices by Eqn.(2)for WUEinst-Ca and WUEinst-Ci curves of T.aestivum(n=5)
3 讨论和结论
3.1 Pn-Ca和Pn-Ci响应曲线的选择
CO2是植物光合作用最重要的原料,也是C3植物光合作用限制的主要限制因子之一。全球CO2浓度快速升高的背景下,探究CO2浓度的升高对植物光合的影响已成为研究热点[25-26]。当前,研究者普遍通过光合作用对CO2响应曲线的测量及拟合,探讨植物光合特性对外界CO2浓度升高的响应。在此基础上可进一步研究植物叶片水分利用效率对CO2的变化趋势。然而,用不同模型拟合光合作用对CO2响应曲线而得到的结果往往存在较大差异,如Michaelis-Menten模型[27]估算植物的Pnmax远高于其实测值,且不能估算出与Pnmax相对应的饱和CO2浓度(Csat)[28-30]。而叶子飘[23]构建的光合作用对CO2的响应模型克服了Michaelis-Menten模型[27]高估Pnmax和不能估算Csat等问题。因此,可以用叶子飘[23]构建的模型拟合植物的Pn-Ca和Pn-Ci响应曲线。虽然用光合仪可以获得植物叶片的Pn-Ca和Pn-Ci两种响应曲线,但目前少有学者对该两种响应曲线进行比较[31]。在本研究中,用式(1)拟合Pn-Ca和Pn-Ci得到的Pnmax与观测值之间均不存在显著差异;拟合得到的CO2补偿点(Γ)也均与其对应的观测值较为接近。然而,式(1)拟合Pn-Ca和Pn-Ci得到的初始羧化效率之间,以及拟合得到的Rp之间均存在显著差异(表1)。这与已有的研究结果相一致[31]。在本研究中,与基于Pn-Ca响应曲线的估算值相比,由Pn-Ci响应曲线估算的小麦初始羧化效率和Rp显著高于Pn-Ca响应曲线(表1)。而高估植物的Rp会导致人们低估植物的碳同化积累,同时高估光呼吸在逆境环境下对植物的保护作用。由此可见,选择不同形式CO2响应曲线会得到不同的初始羧化效率和Rp。推测这是由于胞间CO2浓度(Ci)估算不准确造成的。那么在Pn-Ca和Pn-Ci两种响应曲线的基础上构建的WUE对Ca和Ci的响应模型,在部分拟合数据上也将会有不同。
此外,由式(1)还可以准确判断植物叶片发生磷酸丙糖利用(TPU)限制时的CO2浓度[32-34]。Miao等[34]测量了4种植物的CO2响应曲线,其中80%的测量数据出现了TPU限制现象。虽然FvCB生化模型[35]被广泛地应用于拟合植物的Pn-Ci曲线,并估算发生RuBP限制过渡到TPU限制的CO2浓度(Csat)[45],然而当前FvCB模型拟合Pn-Ci曲线的方法多达6种[34],且这些方法拟合给出的Csat存在较大的差异[34,36]。因此,由FvCB模型估算植物Csat的值并不可靠。而整体来看,由式(1)拟合小麦的CO2响应曲线得到的光合参数,除Rp外均与观测值较为接近(表1)。由此可见,式(1)是一个较为理想的探讨光合速率对CO2响应的数学工具。因此,在式(1)的基础上构建水分利用效率对CO2浓度的响应具可 行性。
3.2 WUEi-Ca和WUEi-Ci响应曲线
由植物的Pn-Ca和Pn-Ci曲线可得到内禀水分利用效率对Ca和Ci的响应曲线,进而可以得到植物的WUEi-max及其相应的饱和CO2(Ci-sat)等。这些参数对评估植物最大潜在的水分利用效率极为重要[37,38]。同时,这一方法已被用作研究植物种间WUEi的差异,以及植物的空间和时间变化时WUEi之间差异的基础[8,9,39,40]。已有的研究结果表明,植物叶片的WUEi会随CO2浓度的增加而提高,其中关键的原因气孔导度会随CO2浓度的升高而减小[26,41-43]。本研究发现,小麦的WUEi随CO2浓度的增加而先直线上升,随后WUEi随CO2浓度的增加而缓慢增加,直到达到最大值,之后WUEi则出现下降趋势。由小麦的WUEi-Ca和WUEi-Ci曲线的变化趋势可知,它不能用指数方程或对数函数进行拟合,其原因是这2个方程或函数均没有极值。而式(2)则可较好地描述小麦的WUEi-Ca和WUEi-Ci曲线,且得到的拟合值与其对应的观测值高度相符。在本研究中,比较WUEi-Ca和WUEi-Ci曲线拟合结果发现,得到的WUEi-max均与观测值非常接近(表2),但尚不清楚为何WUEi-Ci曲线的初始斜率约为WUEi-Ca曲线的2.10倍(表2)。
3.3 WUEinst-Ca和WUEinst-Ci响应曲线
研究表明,植物叶片的WUE在低CO2浓度时会随CO2浓度的增加快速提高,然后进入较为平缓的增长区域,到达最大值之后随CO2浓度的增加而下降[10,15,31]。其中有两方面的原因:(1)在低CO2浓度条件下,光合速率会随CO2浓度的升高而增大直到发生RuBP到TPU的过渡,而后随CO2浓度的增加而下降;(2)气孔导度会随CO2浓度的升高而减小,使得蒸腾速率降低,减少水分的流失[25-26,41-42]。与WUEi-Ca和WUEi-Ci曲线类似,小麦的WUEinst随CO2浓度(Ca或Ci)的增加而先上升直到最大值,之后WUEinst随CO2浓度的增加出现下降。此外,由小麦的WUEinst-Ca和WUEinst-Ci曲线的变化趋势可知,它们同样不能用指数方程或对数函数进行拟合,因为这2个方程或函数描述下的WUEinst是随CO2浓度的增加而一直增加。因此,这2个方程或函数不能很好地描述小麦的WUEinst-Ca和WUEinst-Ci曲线。而式(5)则可以较好地拟合小麦的WUEinst-Ca和WUEinst-Ci曲线,且对比WUEinst-Ca和WUEinst-Ci曲线拟合结果发现,拟合得到的最大WUEinst与其对应的观测值之间不存在显著差异(表3),同样地,尚不清楚为何WUEinst-Ci曲线的初始斜率约为WUEinst-Ca的2.08倍。
总之,新构建的WUE对CO2浓度响应模型即式(2)和式(5),不仅可以很好地拟合小麦叶片的WUE对CO2浓度的响应曲线(包括WUEi-Ca、WUEi-Ci、WUEinst-Ca和WUEinst-Ci曲线),而且还可以直接准确地给出小麦的最大WUEi和WUEinst以及相对应的饱和CO2浓度,且这些参数的拟合值与其对应的观测值之间无显著差异。因此,新构建的WUE对CO2的响应模型为定量研究植物的WUE对未来CO2增加的变化规律提供了有效的数学工具。然而,WUE对Ca和Ci的响应模型给出的初始斜率差异较大,其原因尚有待于进一步研究。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
中原商报·科教研究(2022年1期)2022-05-13
国际太空(2021年11期)2022-01-19
中国化妆品(2017年12期)2017-06-27
物联网技术(2016年11期)2017-01-12
太空探索(2016年7期)2016-07-10
红岩(2015年3期)2015-11-28
高中生学习·高三版(2014年3期)2014-04-29
