水稻镉积累的遗传调控及低镉水稻育种的研究进展与展望
2021-11-11 01:14李东屏郝小花陈良碧田连福
生命科学研究 2021年5期
李东屏,郝小花,陈良碧,田连福
(1.湖南师范大学生命科学学院作物不育资源创新与利用湖南省重点实验室,中国湖南长沙410081;2.湖南文理学院生命与环境科学学院,中国湖南常德415000)
镉(cadmium,Cd)是一种重金属元素,自然条件下土壤本底Cd含量很低。然而,人类的活动,如采矿、冶炼化工等工业生产、污水灌溉、肥料施用等,导致部分耕地Cd含量显著增加。目前国内约有27.8万公顷的耕地被Cd污染[1~2]。
水稻从稻田吸收Cd并在稻米中积累,人们在食用含Cd的大米后,大部分Cd会被吸收并积累在体内产生毒性,危害人体健康。Cd在人体器官中积累,其中肾脏和肝脏的积累量占体内Cd总量的约60%。长期的Cd毒性易导致脏器功能受损,严重的甚至诱发癌变[3~5]。
已有大量的研究表明,遗传因素对Cd吸收和转运起重要调控作用。在相同环境条件下,水稻吸收、转运和积累Cd的能力存在遗传多样性[6~9]。同时,环境因素一方面对稻田Cd形态有重要影响,如土壤酸碱度(pH值)、氧化还原状态(Eh)、其他离子的含量、土壤微生物等会影响游离态Cd的含量,从而影响水稻的Cd吸收;另一方面环境因素的变化也会影响水稻相关基因的表达,从而可能影响水稻Cd的吸收与转运。因此,施用石灰、硒、硅等及全程淹水、施加含有益微生物的菌肥等措施,能钝化稻田中的Cd,进而有效降低水稻根系的Cd吸收,是重要的应急农艺措施,但在执行的可持续性、易操作性或环境友好方面存在一些不足[10~14]。要在Cd污染的稻田生产出Cd含量达标的稻米,主要得依靠筛选和培育低Cd水稻品种。因此,阐明影响水稻Cd吸收、转运及Cd在器官间运输分配的遗传调控因子,可以为低Cd水稻育种提供理论指导。近年来,对水稻Cd积累遗传调控方面的研究取得了较大进展。本文综述了影响水稻Cd积累的遗传因素及籽粒低Cd积累分子育种的相关研究进展。
1 遗传因素对水稻Cd吸收、转运和积累的影响
1.1 水稻镉积累QTL定位及Cd基因克隆
基于稳定的低Cd积累和高Cd积累水稻资源,研究者们构建了多个群体,并将数十个Cd连锁数量性状位点(quantitative trait locus,QTL)定位到12条染色体的特定区域(表 1)[15~36]。例如,qCdT7/qGCd7作为Cd积累的主要QTL,被定位于7号染色体的短臂上[22~24]。利用与Cd紧密连锁的QTL,可以进一步精细定位,克隆到Cd基因,也可以开发出与Cd积累特性关联的分子标记,在分子聚合育种中起辅助作用。
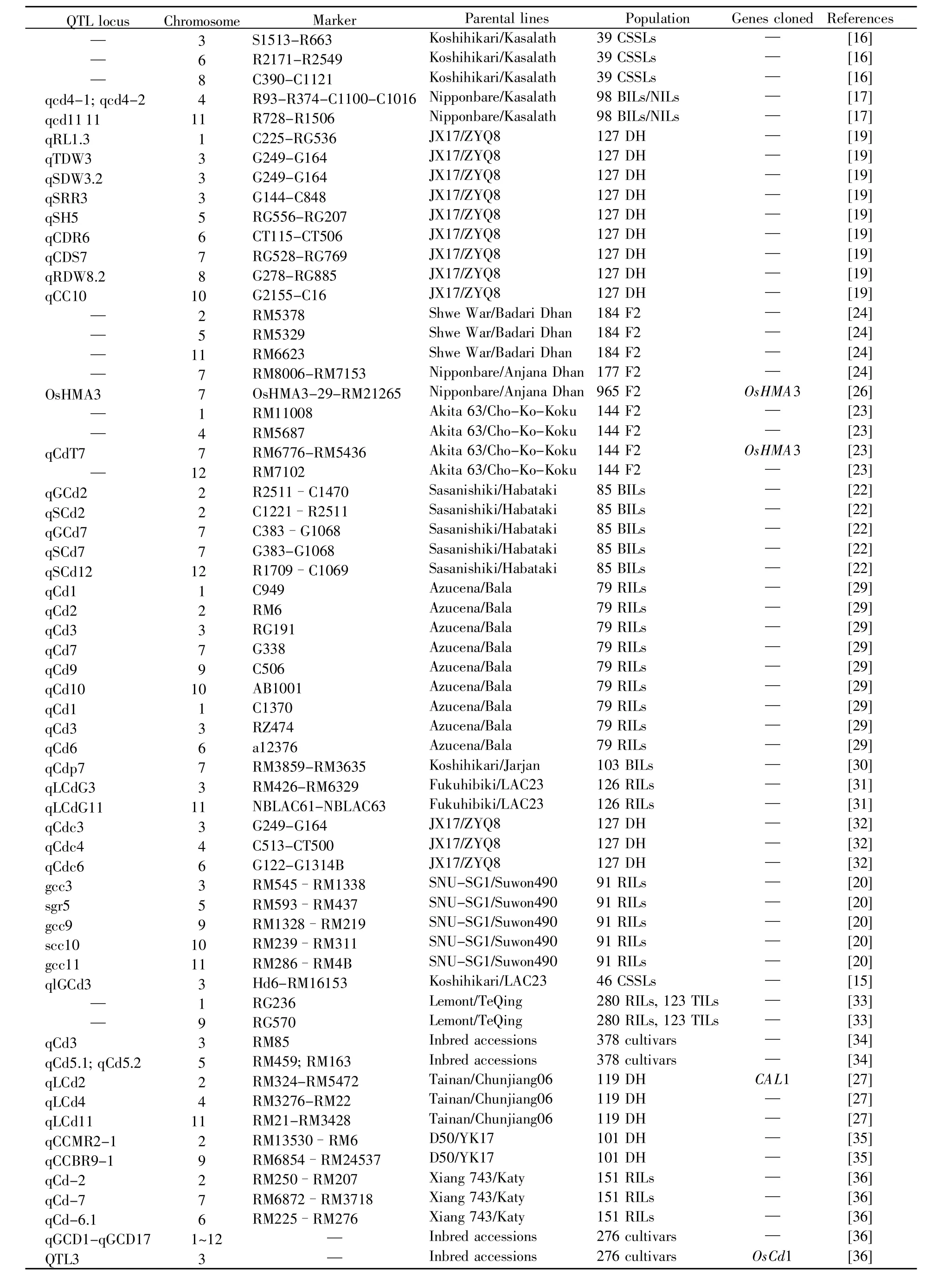
表1 水稻Cd积累QTLsTable 1 QTLs for Cd accumulation in rice
到目前为止,已通过QTL定位鉴定出3个Cd相关基因。OsHMA3编码一种P型ATPase蛋白,定位于所有根细胞的液泡膜。低Cd积累品种中的OsHMA3具有正常功能,它能将Cd螯合到根液泡中,从而减少Cd由根向地上部的转运,降低Cd在地上部和籽粒中的含量。然而,高Cd积累籼稻品种Anjana Dhan的OsHMA3等位基因突变,导致蛋白功能缺失,不能将Cd螯合到根液泡中,导致高Cd品种根-地转运率提高[25~26]。CAL1是通过QTL克隆到的第2个Cd基因,编码一个类防卫素蛋白,富含半胱氨酸残基,能够把Cd螯合在叶组织中但不转移到籽粒[27]。OsCd1是主要易化子超家族转运蛋白,定位在质膜上,调控Cd吸收及籽粒Cd积累[28]。OsCd1中一个具有错义突变的自然变异(Val449Asp)是导致籼稻和粳稻之间水稻籽粒Cd积累差异的原因之一。
1.2 Cd转运体基因
利用各种正向和反向遗传学手段,目前已从水稻中克隆和鉴定出数十个Cd积累相关基因,其中Cd转运体基因在Cd吸收和Cd分配中起着关键的作用(表2)。
1.2.1 自然抗性相关巨噬细胞蛋白(NRAMP)家族基因
NRAMP是跨膜转运蛋白家族,参与抗病及多种离子转运过程。水稻NRAMP基因家族包含7个成员[37],其中3个介导Cd的吸收和转运。Os-NRAMP5是迄今为止发现的参与水稻根部Cd吸收最重要的基因,该基因突变阻断了根部Cd吸收,使得即使在高Cd污染稻田中,osnramp5突变体中的Cd积累量也显著下降,是低Cd水稻育种的关键靶基因[38~42]。OsNRAMP5主要在水稻根中表达,包括根表皮、皮层和维管束近木质部的薄壁细胞,同时在颖壳、叶片和叶鞘等组织也有低表达,暗示OsNRAMP5不仅参与Cd吸收,还参与Cd在水稻组织中的迁移。有趣的是,OsNRAMP5的过表达转基因水稻也显著降低了地上部分,包括籽粒的Cd含量,只增加了根中Cd积累量[43]。Os-NRAMP5过表达水稻根尖组织中OsNRAMP5的定位和极性发生改变,使得更多吸收的Cd被转运进了根部细胞,而不是通过径向运输通路转运到中柱装载进木质部,导致根部Cd积累比野生型高,但地上部分的Cd显著低于野生型[43]。正常情况下,OsNRAMP5介导锰和铁在水稻体内的吸收、根-茎转运以及在根和叶中的分布,在维持水稻体内锰、铁离子的动态平衡中起着重要作用[38~40,44]。osnramp5强突变体中必需微量元素锰的含量大幅下降,导致农艺性状(如株高、穗型、结实率和产量)受到显著影响,但增加根际环境中的锰离子含量可以部分回复突变对农艺性状的负效应,或者在不同OsNRAMP5位点进行突变,可以培育出既不影响农艺性状又能大幅减低Cd吸收的弱突变体材料[41~43,45]。OsNRAMP5过表达对水稻的株高、穗型和产量也有一定的负效应[43]。
除了OsNRAMP5,家族成员OsNRAMP1也可以吸收和转运Cd和Mn[46~47]。OsNRAMP1主要在根部除中央维管组织外的其他细胞和叶肉细胞的质膜上表达,受Cd处理和缺Fe诱导。OsNRAMP1基因敲除突变体osnramp1的根和地上部分包括籽粒中的Cd和Mn积累显著降低,与osnramp5突变体的Cd和Mn积累表型相似,但osnramp1突变体降低Cd和Mn的能力要小于osnramp5,且osnramp1 osnramp5双突变体中Cd和Mn的积累量比单突变体降得更多[47]。不同Cd积累品种间OsNRAMP1蛋白的氨基酸序列无显著差异,但不同Cd积累品种间OsNRAMP1蛋白的表达水平存在显著差异。高Cd品种的OsNRAMP1蛋白表达水平显著高于低Cd品种[46]。OsNRAMP1在拟南芥异源的过表达增加Cd在叶片中的积累,OsNRAMP1还参与As3+的转运[48]。另外,OsNRAMP1在酵母中显示有转运Fe的活性,但在突变体osnramp1中未能观察到Fe相关表型,暗示OsNRAMP1可能与其他转运体在水稻Fe吸收和转运中存在功能冗余[48]。
此外,OsNRAMP2也参与Cd的转运。Os-NRAMP2定位在液泡膜上,通过介导Cd从液泡到细胞质运输参与籽粒Cd积累[49]。
OsNRAMP其他成员参与Mn、Fe或Al的运输,但不运输Cd。其中,OsNRAMP3介导锰在水稻中的分布,参与锰从老叶运输到新叶的再分配[50~51]。OsNRAMP4/OsNrat1介导根细胞对Al3+的吸收并促进铝解毒[52~53],还可能介导Cs吸收[54]。OsNRAMP6在酵母中表达具有铁和锰的转运活性,但是否在水稻铁和/或锰/或Cd的吸收或转运中起作用尚不清楚[55]。
1.2.2 P型重金属ATP酶(HMA)家族基因
HMA是一种利用ATP水解释放的能量驱动重金属离子跨膜转运的转运体。水稻HMA基因家族共有9个成员,其中OsHMA3和OsHMA2先后被鉴定为Cd运输关键基因。
OsHMA3(qCdT7)是利用QTL定位克隆到的首个Cd转运基因,在Cd由根部到地上的转运调控中起着关键的作用,是低Cd水稻育种的重要候选基因[23,25~26]。OsHMA3位于水稻根部细胞液泡膜上,有功能的OsHMA3负责将Cd转运到根部细胞液泡中贮存,降低Cd通过木质部装载转运到地上组织的能力,导致地上组织包括籽粒的Cd含量比较低。粳稻品种一般具有有功能的OsHMA3,籽粒Cd积累量较低。而籼稻由于OsHMA3基因突变导致蛋白质功能失活,不能将吸收的Cd扣留至根部细胞的液泡中,使得Cd从根部转运到地上部包括籽粒中的活性增强,籽粒Cd积累量较高,这被认为籼稻品种籽粒Cd含量普遍高于粳稻的主要原因之一[23,25~26]。进一步研究发现,该基因自然变异较丰富,可分为8种基因型,其中Ⅰ、Ⅵ、Ⅴ型是具有功能的基因,而Ⅱ、Ⅷ型是功能失活型。Ⅰ型为有功能的基因型,为低Cd品种类型,主要为日本晴等粳稻,但“洞庭晚籼”、PA64S等带有粳稻血缘的籼稻品种也属于该类型;Ⅱ型是功能失活型,为高Cd积累粳稻品种类型;Ⅴ型也是有功能的基因型,但却是高Cd籼稻品种类型,93-11、中优早81等属于这种类型,它们的这种高Cd特性不能用OsHMA3活性的有无来解释[56~57]。最近,研究人员通过对PA64S杂93-11构建的NIL群体进行QTL定位和图位克隆分析,发现OsHMA3启动子的序列存在变异,尤其是-683 bp到-557 bp之间的7个核苷酸差异,是OsHMA的两个等位基因GCC793-11和GCC7PA64S的差异表达(分别表达低和高)所必需的。将两个启动子互换,则GCC793-11和GCC7PA64S的表达会发生相反的变化(分别升高和降低),根和地上组织的Cd积累也会发生相应的变化,表明在Ⅰ、Ⅱ、Ⅴ型等具有功能性OsHMA3的水稻品种中,启动子遗传差异对品种间Cd积累多样性的调控起重要的作用。籼稻中主要是GCC793-11等位基因,粳稻中主要是GCC7PA64S等位基因,GCC7PA64S等可以应用到低Cd籼稻育种[58]。同时,无论籼稻还是粳稻,功能性OsHMA3过表达使得根-茎Cd转运效率显著下降,Cd耐受能力显著增加,地上组织的Cd含量显著减低,过表达籼稻糙米中的Cd积累量可以降低90%以上,且OsHMA3过表达对水稻株高、产量及稻米品质等农艺性状无影响[59~60]。OsHMA3是一个功能性的Zn转运蛋白,但其无论是失活或过表达,对水稻地上部分的Zn稳态似乎影响不大[61]。把水稻OsHMA3转到小麦过表达,能大幅降低小麦籽粒中的Cd含量,显示了该基因在作物低Cd籽粒育种中的重要潜力[62]。
OsHMA2是一个质膜定位的锌离子转运体基因,在营养生长阶段在根部中柱鞘组织组成型表达,介导Cd和Zn从根到地上部的运输;在生殖生长阶段,还在地上部分的维管束,尤其是水稻倒一节(nodeⅠ)中的大维管束和分散维管束的韧皮部细胞膜上表达。敲除突变体中穗颈节及穗中的Cd含量显著降低,表明OsHMA2在穗颈节部位介导Cd从木质部到韧皮部的转移,以及通过节向穗的分配[63~65]。有趣的是,OsHMA2过表达的转基因水稻和hma2突变体的籽粒中Cd含量都低于野生型[63]。最近,研究人员把OsHMA2启动子驱动OsHMA3表达,转基因水稻根中Cd增加,地上部分Cd降低,其中籽粒中Cd含量减少到对照的1/10水平[66]。
1.2.3 参与Cd运输的其他转运体基因
OsLCT1(低亲和性阳离子转运体基因)定位于质膜上,主要在生殖生长成熟期的穗颈节大维管束周围和分散的维管束中表达,介导Cd通过韧皮部的外运向籽粒分配。OsLCT1功能缺失株系改变Cd在节内由韧皮部向籽粒的转运,籽粒中Cd含量仅为野生型的50%,其他金属元素含量与野生型无显著差异,且对水稻生长无影响[67]。除了低亲和性阳离子转运体基因,水稻ZIP基因家族的许多成员也被鉴定介导Cd吸收和转运,如OsZIP1[68~69]、OsZIP3[68]、OsZIP6[70]、OsZIP7[71]。其中 Os-ZIP1编码一种金属解毒转运蛋白,通过激活其外排转运活性介导水稻中过量的锌、铜和Cd积累。OsZIP7编码一种内向转运蛋白,在根中木质部的装载和节中的维管间转移中起着不可或缺的作用,优先向发育中的组织和水稻籽粒输送锌和Cd。亚铁离子转运体基因OsIRT1和OsIRT2也属于OsZIP基因家族成员,但在系统树上成单独的一支,与其他成员的亲缘关系较远。OsIRT1和Os-IRT2也转运Cd,缺铁可增强OsIRT1-/OsIRT2介导的Cd吸收和转运[72]。OsIRT1过表达的水稻对过量的锌和Cd表现出更高的敏感性[73]。
此外,属于水稻金属耐性蛋白基因家族(MTP)的成员OsMTP1/OZT1,赋予植物对锌和Cd的耐受性。在其RNAi干扰植株中,苗期对重金属的敏感性提高;在低浓度重金属胁迫下,成熟期不同器官重金属积累发生改变,提示该基因在锌和Cd的转运以及维持其内稳态方面发挥着重要作用[74]。ABC转运体基因家族成员也参与Cd在细胞中的外向运输,能够增强植株对Cd的耐性。OsABCC9定位于根部中柱细胞的液泡膜上,受Cd快速诱导表达。其基因敲除水稻对Cd敏感,根和地上部分包括籽粒中的Cd含量都比对照高[75]。OsABCG36是质膜表达的转运体基因,在根和地上都有低表达,在根中除了表皮细胞以外的其他细胞中都有分布,其表达受Cd快速诱导表达。其基因敲除水稻对Cd敏感,根中Cd含量比对照高,但地上部分Cd含量不受影响[76]。另外,钙/阳离子反向转运体基因OsCCX2也被证明参与水稻Cd的运输和分配,功能缺失突变体各器官中Cd的分配发生变化,同时籽粒中Cd含量降低[77]。OsCCX2导致的Cd分配和积累发生变化的机理还不清楚,其作用机制尚需进一步研究。此外,主要易化子超家族(MFS)基因OsCd1,也参与水稻根系Cd吸收,并参与水稻籽粒Cd积累[28]。
1.3 镉螯合相关基因
植物螯合素(PCs)是由植物螯合素合成酶(PCS)合成的一类富含巯基、具有较强Cd螯合活性的小肽。PCs与游离巯基处的Cd离子形成螯合物,随后将结合的Cd隔离在液泡内[78]。PCS催化谷胱甘肽(GSH)非核糖体合成PCs,GSH在Cd螯合和解毒中起重要作用[79]。水稻籽粒中OsPCS1和Os-PCS2特异性表达下调导致籽粒中Cd和As含量显著降低,表明PCs合成会增强水稻籽粒中Cd和 As的积累[80~81]。PCs合成的前体,如 GSH、谷氨酸或半胱氨酸,在Cd耐受中也起着重要作用。这些前体合酶基因的表达变化也会改变植物的Cd耐受性[82~83]。
最近,一种类防御素基因编码的蛋白CAL1被证明能改变Cd的螯合作用和Cd向细胞外空间的分泌,从而降低细胞内Cd的浓度,同时增强Cd通过木质部导管的远距离运输。CAL1可通过木质部特异调控Cd从根到地上部的转运,但不能通过韧皮部调控Cd从地上部到籽粒的转运,从而导致叶片而不是籽粒Cd的积累。以上信息为培育双功能水稻品种提供了一个潜在的目标,即在修复水稻土的同时生产Cd安全谷物[27]。
1.4 水稻Cd积累的调控基因
尽管有许多关键的转运蛋白负责Cd的吸收和转运,但关于水稻对Cd转运活性的调控知之甚少。基因表达的转录调控是控制Cd转运活性的有效方法之一,其中转录调控因子和microRNAs(miRNAs)等起着重要作用。转录调节因子Os-TITANIA是一种水稻PHD指状蛋白,通过调控多种金属转运蛋白基因的表达发挥作用。突变株LC5表现出低Cd积累特性[84]。OsHB4参与调节Cd的积累,OsHB4的过表达增加Cd的敏感性和Cd在叶片和籽粒中的积累。然而,敲除OsHB4可增强Cd耐受性。miR166以OsHB4为靶点,降解OsHB4转录物,减轻Cd积累[85]。miR390也参与水稻对Cd的耐受和积累。OsSRK是miR390的靶基因,miR390在Cd处理后表达量上调。过量表达的miR390表现低镉耐受性和高镉积累。同时,Os-NRAMP5和OsHMA2表达水平增加,表明miR-390是水稻镉胁迫耐受性的负调节因子[86]。
泛素介导的26S蛋白酶体降解是翻译后调控的重要机制。水稻E3连接酶基因OsHIR1在As和Cd吸收中发挥作用。E3泛素连接酶OsHIR1通过与OsTIP4;1转运蛋白相互作用将OsTIP4;1泛素化,后者被26S蛋白酶体系统识别并降解。OsHIR1的过量表达导致植物体内Cd和As的累积显著减少,表明OsHIR1 E3连接酶正调控Os-TIP4;1,影响 Cd 的摄取[87]。
OsLCD(low-Cd)是水稻维管束组织中优势表达的基因,编码一种可溶性蛋白,参与水稻对Cd胁迫的耐受性及Cd的转运和积累,主要表达于根的维管束和叶片韧皮部的伴胞中,定位于细胞质和细胞核。在Cd浓度较低的土壤中,lcd突变体籽粒中Cd含量比野生型降低一半,但其营养器官中Cd含量差异不显著[88]。OsLCD不具有转运蛋白和解毒蛋白的特征,推测OsLCD可能通过激活其他Cd转运体介导Cd的积累;通过酵母双杂交实验,研究人员已筛选出与运输和调控相关的蛋白质,为进一步阐明OsLCD介导的水稻Cd积累机制提供了线索[89]。
2 水稻籽粒低Cd积累的分子育种
2.1 低Cd水稻品种的筛选与鉴定
研究者对具有不同Cd累积性状的水稻品种进行了有效的基因分型筛选和鉴定[90~92]。许多品种被报道为低Cd或高Cd类型(表3),其中一些表现出Cd超积累特性,如Cho-ko Koku、Akita110、TJTT8等,有应用于污染土壤植物修复的潜力[24,93~96]。高Cd粳稻品种失去OsHMA3(温带粳稻)或Os-NRAMP2(热带粳稻)基因的功能[49,57],而一些低Cd籼稻品种与OsNRAMP5、OsNRAMP2或OsHMA3基因的功能密切相关[42,45,60]。其中OsNRAMP5对低Cd品种的培育是最关键的,通过基因编辑技术敲除OsNRAMP5,突变体Cd污染环境中的Cd含量大幅减低,远远低于国家大米Cd安全水平(0.2 mg/kg)。最近,湖南杂交水稻研究中心的研究人员从国内外大量的水稻资源中,筛选到武汉大学朱英国院士等培育的红莲型籼稻不育系珞红3A和珞红4A(https://www.ricedata.cn/variety/varis/614203.htm)的Cd积累量特别低(分别为0.01 mg/kg和0.03 mg/kg),其他品种最低为0.16 mg/kg,最高为1.92 mg/kg,平均0.75 mg/kg,说明珞红3A和珞红4A是典型的低Cd积累材料。通过基因组测序,研究人员发现7号染色体上缺失一段包含OsNRAMP5、OsNRAMP1基因的片段(408 Mb,Org),插入一段2 980 bp(Tons)的片段,是自然突变的Cd低吸收品种[97]。另外,有些籽粒低Cd水稻品种还具有茎秆高Cd的特性,如四川农业大学培育出的恢复系品种“雅恢2816”[97](www.ricedata.cn/variety/varis/618270.htm),以及湖南师范大学陈良碧教授培育的温敏不育系品种KT27S(https://www.ricedata.cn/variety/varis/615324.htm),但它们的Cd调控机理尚不明了。尽管如此,这类基因型品种有望成为既能生产出Cd安全稻米,又能修复土壤的“两用”资源材料。
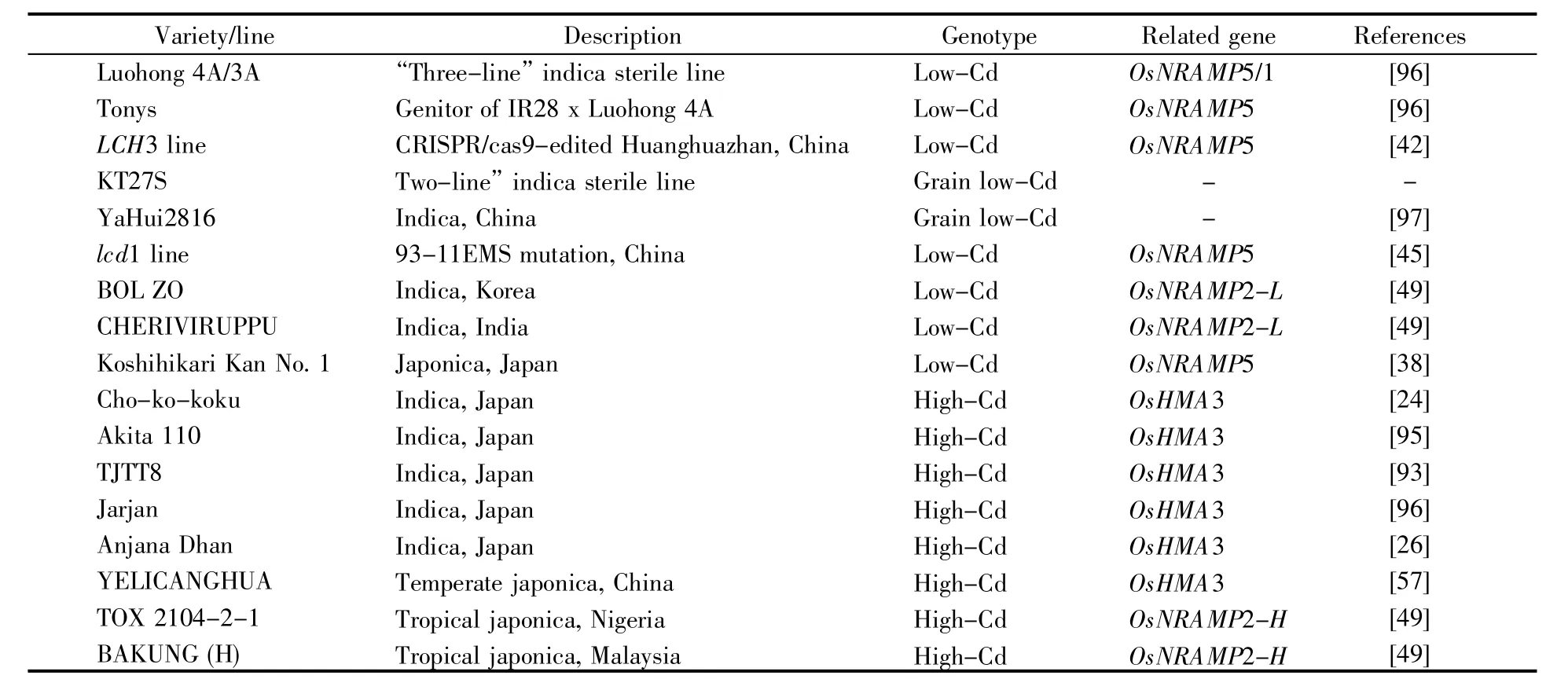
表3 不同Cd积累基因型的品种(株系)Table 3 Varieties/lines of different Cd accumulation genotypes
2.2 低Cd分子标记辅助的水稻低Cd基因聚合育种
随着水稻Cd吸收和转运有关的关键基因被定位和克隆,OsHMA3、OsNRAMP5、OsNRAMP1、OsNRAMP2等的分子标记也被开发。OsNRAMP5功能失活等位基因(lcd1、lcd-kmt2、LCH3)被认为可以显著降低根对Cd的吸收,但如果突变位置发生在基因3′末端附近,则不会影响锰的吸收和产量[39,41~42,45]。此外,粳稻类型的启动子驱动功能性的OsHMA3表达,可以将Cd螯合到根液泡中[26,60]。这些发现显示了将多个低Cd基因聚合到一个品种中进行低Cd水稻育种的潜力。通过与低Cd品种的杂交和轮回亲本的连续回交,许多轮回亲本的籽粒Cd积累性状得到了较大的改善。对于杂交水稻育种,首先将雄性不育系和恢复系培育为低Cd亲本,然后通过杂交获得低Cd杂交种子。随着人们对水稻籽粒Cd积累机制的深入了解,利用低Cd基因聚合的方法进行低Cd积累水稻培育研究已经进行了初步尝试,如隆平高科种业科学研究院的杨远柱研究员将粳稻的OsHMA3-OsNRAMP5-OsNRAMP1转育到籼稻93-11,改良93-11籽粒中的Cd积累大幅下降[98]。目前,已有珞红3A、珞红4A等超低Cd积累材料,利用轮回亲本通过与其的杂交及连续回交,再利用Tons分子标记筛选转育成功的轮回亲本Tonys,经检测是超低Cd材料,显示了调控OsNRAMP5活性在低Cd水稻育种的光明前景,有望通过调控这一个Cd吸收基因解决稻米Cd安全问题[99]。另外,在低Cd品种的基础上,可以把营养品质和产量方面的调控基因聚合在一起,如培育高锌、高硒的低Cd 品种[100]。
3 结论和展望
Cd暴露会对包括人类在内的有机体造成一系列有害影响。因此,了解水稻对Cd的吸收、转运和积累机制,对于加强有效地降低Cd的策略具有重要意义。近20年来,随着一些介导Cd转运的基因在水稻中的发现,对Cd转运和影响Cd积累的环境因子的研究取得了进展,阐明了稻田土壤酸碱度、土壤通气状况、根际微生物、环境中其他离子和化合物等主要环境因子对水稻Cd积累的影响及其机理,提出了降低水稻籽粒Cd积累的对策。大量田间试验表明,低Cd水稻品种,结合水稻全生育期的灌溉和较高的pH调节,可显著降低籽粒Cd的积累。一些根际微生物在缓解水稻Cd吸收和转运过程中也起着重要作用。此外,施用多种叶面复合肥、生物炭、酸碱改良剂等农艺措施也能有效地减少籽粒Cd的积累。
对于籽粒低Cd品种的选育,近期目标是培育低吸收Cd或不吸收Cd的材料,以便在各种Cd污染稻田中能生产出镉达标的稻米。利用珞红4A/3A等超低镉品种,通过杂交置换培育各种低Cd水稻品种,解决稻米Cd超标的问题,应该在不长的时间里可以达到。远期目标是培育Cd高吸收、营养体尤其是茎叶高积累Cd,但籽粒低Cd的“两用”水稻品种,以在生产安全大米的同时,能够随着稻草的移除,逐步修复土壤。目前,KT27S、雅恢2816等品种已被鉴定具有茎秆高Cd、籽粒低Cd的特性。类防卫素蛋白OsCAL1能够螯合Cd,并将其外排到胞外,可促进根部Cd通过导管运送到地上,但阻碍了通过韧皮部向籽粒的Cd转移,是稻草高Cd、籽粒低Cd的基因聚合育种中是重要的遗传资源[27]。在未来,须进一步从低Cd材料中筛选和鉴定茎秆超高Cd积累水稻资源,并进行遗传定位和克隆,鉴定既能高效吸收Cd,又能高效控制Cd转移到籽粒的关键基因,为“两用”低Cd水稻育种提供遗传资源。
CRISPR/Cas9的基因编辑技术具有简单、高效、多功能等优点,可以对染色体上特定基因进行基因修饰,比利用杂交转育的方法转移突变等位基因要简单快速得多。利用CRISPR/Cas9系统进行低Cd水稻遗传改良已取得较大进展[41~42],但受转基因政策的限制尚不能应用。由于可以从基因编辑植株后代中筛选出含有非转基因元件的株系,很多国家或地区已经把利用CRISPR/Cas9编辑的材料不纳入转基因材料进行管理。随着技术和管理的进一步成熟与完善,基因编辑技术创制的低Cd水稻品种一定会走向市场应用。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
今日农业(2021年15期)2021-10-14
农业科技通讯(2021年1期)2021-03-06
中国药剂学杂志(网络版)(2021年1期)2021-02-24
农民致富之友(2020年32期)2020-12-03
中国农业科技导报(2020年3期)2020-03-15
中国食品学报(2019年12期)2019-01-13
中国人兽共患病学报(2018年10期)2018-12-08
中国当代医药(2018年21期)2018-11-10
农村百事通(2017年8期)2017-04-26