
超级稻超高产形成的光合机制研究进展
2021-11-11 01:13:54徐孟亮丁绿萍徐甜甜吴合周
生命科学研究 2021年5期
徐孟亮,丁绿萍,罗 佳,徐甜甜,吴合周
(1.湖南师范大学生命科学学院作物不育资源创新与利用湖南省重点实验室,中国湖南长沙410081;2.湖南桃花源农业科技股份有限公司,中国湖南常德415001)
全球每天有近半的人口需要食用稻米,从中摄取约2/3的能量与3/5的蛋白质。稻米在我国居民口粮消费中占60%,产量长期居粮食作物之首,对于保障我国乃至全球的粮食安全至关重要。随着全球人口的不断增加及购买力的不断增强,人们对稻米的需求量将相应增加。与此同时,由于工业化和城镇化的快速发展、稻田的重金属污染、自然灾害损毁等,可耕稻田呈逐渐减少趋势。如何在更小的稻田面积上生产出更多的粮食(即提高单产)以确保稻米安全供应成为了全球面临的紧迫而必须解决的重大问题。为此,日本、国际水稻所和我国先后提出了水稻超高产育种计划或超级稻育种计划[1~2]。通过形态改良与籼粳亚种间杂种优势利用相结合的育种途径,我国已提前或如期实现农业部1996年提出的一季籼型超级杂交稻第一期至第四期育种目标(即连续两年在同一生态区不同点的两个百亩片实现10.5 t/ha、12 t/ha、13.5 t/ha及15 t/ha的产量指标),并于2015年启动第五期超级稻育种计划,其产量指标是16 t/ha[3~4]。目前,云南个旧与河北永年县也实现了这一目标,正朝17 t/ha的目标迈进[3],大幅领先日本与国际水稻研究所。超级稻在我国的推广,为保障我国粮食安全作出了重要贡献。在超级稻育种不断取得单产突破的同时,为了能更好地指导超级稻的育种与栽培,许多学者对超级稻超高产形成的光合株型特征与光合生理特性进行了研究,并取得重要进展。
1 超级稻的光合株型特征及群体结构特点
1.1 超级稻的光合株型特征
优良的植株形态及群体结构是水稻高光效、高产及超高产形成的形态学基础。20世纪60年代,日本人松岛省三从栽培角度提出了“理想稻”的株型模式并制订了培养“理想稻”的6条标准,即:1)单位面积有足够的粒数;2)矮秆、短穗、多穗;3)出穗后上部3~4叶短、厚、直,叶面积指数约为5;4)出穗后叶色浓绿不淡;5)每茎绿叶数多,直至成熟期止;6)出穗前15天及出穗后25天天气好[5~6]。1968年,澳大利亚人C.M.Donald[7]提出了有别于以往基于“性状改良”或基于“产量选择”的作物育种新方法,即基于“模型设计”(源于飞机模型设计理念)的作物理想株型设计育种。他在文中指出:“作物的理想株型是指依据现有生理学和形态学理论知识、育种经验及育种技术将一些有利于作物光合作用、生长或产量的优良形态性状聚合起来的理想植株形态。具有理想株型的个体,不但自身形态优良,而且在群体中的竞争力弱、对群体中其他个体的影响小,因而群体能最经济地利用环境资源(如光能、地力等)来最大限度地生产同化物。禾本科矮而粗壮的茎秆、直立的叶及大穗,大麦与小麦的芒(能进行光合作用)以及因地域而异的优良抗病虫性与适宜的熟期等优良性状都是构成理想株型的要素。今后的作物育种,均应基于理想株型育种,以提高育种效率”。自作物理想株型育种新思路提出以后,国内外一些水稻育种家也将这一新的育种理念应用到水稻超高产育种上,并提出了多个水稻超高产的理想株型模式。例如:20世纪70年代,针对我国华南地区台风暴雨频繁、高温多湿及昼夜温差小的气候特点,为增强品种的耐密性,进一步解决多穗数与高穗重的矛盾,提高增产潜力,黄耀祥等[8~9]提出了“丛化育种”的策略和“丛生、快长”的新矮秆株型模式,即营养生长期丛生矮生、生殖生长期快长的新矮秆株型,该株型能充分利用光能,实现高产、超高产并得到实践的验证[9]。杨守仁等[10]总结出矮秆稻种的三大特点:耐肥抗倒、适于密植、谷草比大,并认为“理想株形”都是基于这三大特点发展而来,没有统一的模式,可以因地制宜,并提出通过亚种间杂交进行超高产“理想株形”育种的研究路线。周开达等[11]经过多年的超高产育种探索,认为超高产育种必须通过亚种间杂交选择“重穗型”组合,并提出了“重穗型”株型模式,其特点是:1)根:根系强大,根粗,功能时间长,衰退缓慢;2)茎与株型:要能支撑5 g以上的大穗,要求茎杆坚韧、杆壁厚、输导组织发达,维管束中有一定的叶绿素,株高120 cm左右,株型松紧适中,每亩有效穗15万左右;3)叶:分蘖期叶片稍散,拔节后叶角较小,叶肉较厚,叶片内卷直立,叶色较深,各生育阶段的色泽变化明显,黄熟期转色顺调,熟色好,剑叶和倒二叶长度40 cm左右,保持功能叶层厚度在100 cm左右。此外,稻瘟病、白叶枯病中抗以上,品质优于汕优63等。国际水稻研究所的G.S.Khush[2]基于经济产量与生物产量及收获指数的函数关系,提出了突破当时热带高产纯系品种产量潜力的水稻新株型模式(图1)。其主要特点是:低分蘖力(直播3~4个,移栽8~10个),没有无效分蘖,每穗200~250粒,株高90~l00 cm,茎秆强健,叶片深绿、厚、直立,根系粗壮、深扎、活力强,生育期100~130 d,抗多种病虫害,米质可接受。该模式设计的生物学产量潜力由当时热带高产纯系品种产量潜力的20 t/ha提高到22 t/ha,收获指数由50%提高到60%(谷/草重量比=6∶4),经济产量潜力可达13 t/ha左右,比当时热带的高产纯系品种增产20%。在此基础上,新株型稻将用于籼粳亚种间杂交稻选育,产量可再增加20%~25%,二者相结合,可把热带水稻的产量潜力提高50%。1995年,湖南杂交水稻研究中心对国际水稻研究所初选的21个新株型品系在长沙的表现进行过观察,发现这些材料的优点是秆粗、穗大、分蘖少,但籽粒不充实,产量很低,认为它们存在库大源不足的缺点[12]。后来的新株型育种研究虽取得一些进展[13~14],但至今未能像我国一样取得巨大成就,不过该理想株型的一些指标对水稻的超高产育种还是有借鉴作用的。受培矮64S/E32在江苏试种时的超高产表现和其植株形态特征的启发,袁隆平[1,3,12]于1997年及时提出了长江中下游生态区中熟中稻(生育期130 d左右)的超高产植株形态模式(图2)及其具体指标。这个形态模式有3个突出特点。第一,高冠层。由叶片组成的冠层高达120 cm以上,上部三片功能叶表现为长、直、窄、凹、厚。修长直挺的叶片,不仅叶面积较大,而且可两面受光和互不遮蔽;窄而略凹的叶片,所占的空间面积小,但整叶的面积并不因窄而减少,同时凹形有利于叶片紧挺不披;较厚的叶片,其光合功能高且不易早衰。因此,该形态模式对光能和CO2摄取率较高。总之,具有这种叶片形态结构的水稻品种,才能有最大的有效叶面积指数和群体光能利用率,为超高产提供充足的同化产物;第二,大库容。在每公顷270~300万穗的情况下,每穗颖花数为250粒左右,每公顷颖花数可高达6.75亿以上;第三,矮穗层。高度抗倒。
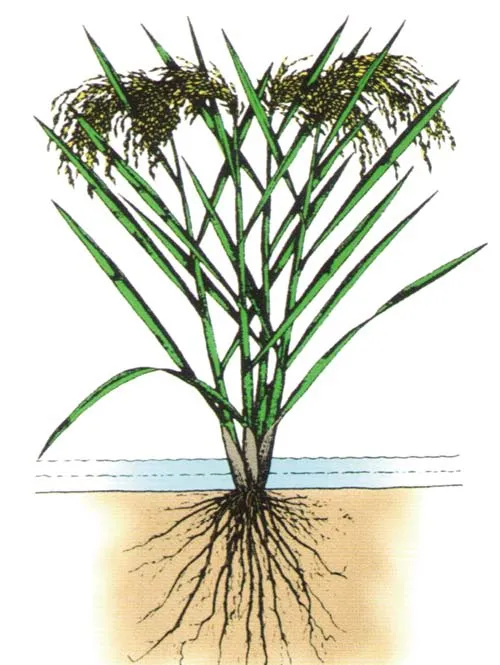
图1 国际水稻研究所设计的新株型水稻(引自文献[1])Fig.1 A New plant type of rice designed by the International Rice Research Institute(quoted from Reference[1])

图2 袁隆平提出的超高产水稻的形态模式(基于文献[12])1:高冠层;2:中大穗;3矮穗层。Fig.2 The morphological model of super high-grainyield rice proposed by Yuan Longping(based on Reference[12])1:High canopy;2:Medium big spike;3:Low spike layer.
在超级稻根系形态与生理特点方面,已有许多研究,总的来说,超级杂交稻较普通杂交稻在根系上有明显优势。朱德峰等[15]的研究表明,超级杂交稻协优9308的根系平均总生物量、根系密度、根系平均深度均比对照汕优63、特三矮2号和65002大或深。全永明[16]观测到,超级稻培矮64S/E32根丛总量与根质量明显高于对照汕优63,分布也比汕优63深。唐文邦等[17]的研究表明,C两优系列组合的根系干质量、不定根条数、不定根总长、根系总体积、发根力、根系总吸收面积、根系活跃吸收面积和根系α-萘胺氧化量显著高于对照汕优63。袁小乐等[18]的研究表明,超级稻在抽穗后15 d(早稻)或20 d(晚稻)的根量与对照相比有明显优势。龚金龙等[19]的研究表明,与超级杂交籼稻相比,常规粳型超级稻抽穗后根系生长优势不断加大,特别是群体生长优势,成熟期粳稻所有根系形态生理特征指标均优于籼稻,是粳稻高产形成的重要原因和保障。褚光等[20]研究了超级杂交稻两优培九(籼稻)和扬粳4038(粳稻)及常规高产品种扬稻6号(籼稻)和扬辐粳8号(粳稻)各生育期根系形态生理特征及其与产量形成的关系。结果表明,超级稻品种在整个生育期较大的根量、生育早中期较高的单株根系氧化力、根系总吸收表面积与活跃吸收表面积及根系中较高的玉米素与玉米素核苷含量是其产量库容大和产量高的重要原因。同时,他们也观察到超级稻存在着结实率较低的问题,推测与其在灌浆期根系活性下降较快有关,并认为提高超级稻灌浆期根系活性,是提高其结实率、促进其产量进一步提高的重要途径。魏中伟等[21]比较研究了超级杂交水稻超优1000与对照Y两优1号的根系性状差异,结果发现,与Y两优1号相比,超优1000各生育时期的根干重、根冠比、根体积、根密度、根系伤流量及颖花根流量较大,且生育后期根干重、根系伤流量下降幅度较小,下层土壤(10 cm以下)的根干重、根体积占其总量的比例较大,扎根较深。Meng等[22]以两个籼/粳杂交稻、两个粳/粳杂交稻和两个籼/籼杂交稻为材料,在浙江宁波与江苏杨州两地各进行了为期两年的根系形态生理特征与超高产的关系研究,认为籼/粳杂交稻抽穗后,更好的根系形态生理特性(更长、更大、更深、更有活力的根)有助于茎秆保持绿色性状,从而能积累更多的同化产物,并最终形成比粳/粳杂交稻和籼/籼杂交稻更高的产量。
1.2 超级稻的群体结构特点
在超级稻群体结构特点方面也有一些研究报道。刘建丰等[23]的研究表明,与对照汕优63相比,高产杂交稻组合剑叶长度增加,叶角较小,冠层光分布比较均匀、消光系数小。邓启云等[24]对具有超级杂交稻模式株型的培矮64S/E32的群体光合优势进行了研究,与三系杂交稻汕优63相比,培矮64S/E32的叶面积指数、群体光合生产力优势明显。Wu等[25]的研究表明,超级中籼杂交稻具有叶面积指数大和叶日积(=叶面积指数×日数,又称光合势)高的特点。Katsura等[26]研究表明,两优培九有较大的叶面积指数持续时间。Zhong等[27]以20个依产量高低划分为高产组、中产组和低产组的中籼杂交稻组合为材料,对它们的产量性状和穗、叶、茎形态特征与产量的关系在重庆进行了为期两年的田间试验,结果表明:高产组产量高的主要原因是单位面积有效穗数显著高于中产组和低产组;在株型构成因素中,叶型因素是导致3组产量差异的最重要原因。平均叶卷曲指数、平均叶面积、平均叶宽、平均叶长宽比、平均叶缘距离、平均叶基角、平均叶下垂角和平均上三叶叶枕距与产量呈显著正相关或显著负相关,其中叶下垂角是影响产量的最重要因素。基于这些结果,他们进一步量化了高产组的叶型指标值并构建了超高产杂交中稻的理想株型。
基于对上述研究工作的综合分析,我们认为,超级稻植株形态及群体结构一般具有如下特点:1)茎秆较高(株高100 cm左右)、粗壮,植株高度抗倒;2)后三叶叶面积大、叶厚、叶色浓绿、寿命长不早衰,叶与穗轴间夹角小(10°~20°),叶挺直不披垂且叶面凹陷呈瓦状;3)根系发达、健旺;4)穗大粒多、穗重,单穗重大于5 g;5)冠层厚(大于100 cm)、叶面积指数较大(一般为 6~7)、叶日积(光合势)大、冠层结构合理、冠层消光系数小,单位土地面积上承载的颖花数量多,生物量大(地上部分生物学产量潜力大于20.0 t/ha),稻谷产量潜力大于10.5 t/ha,收获指数略大于0.5;6)抗性强,米质可接受。这样的株型与群体结构特点有助于提高光能利用率从而实现超高产。但各地生态条件有所不同,超级稻株型及群体结构也应因地而异。
2 超级稻的光合生理特性
优良的光合生理特性是水稻高光效、超高产形成的生理学基础。两优培九(培矮64S/93-11)是实现我国超级稻育种计划第一期育种目标的标志性组合,有关其光合生理特性的研究工作有很多报道[28~32]。吕川根等[33]对其进行了详细综述,并指出:两优培九在正常生长和衰老过程中,光能的吸收、传递与转换效率以及碳同化等方面均具有优势,光合性能优异;高产杂交稻育种要做到“遗传上有性状杂种优势,形态上有优良光合株型,功能上有优异光合性能”的遗传、形态和功能“三优”相结合。我们曾以汕优63与93-11为对照,对其抽穗后结实期的功能叶面积、叶绿素含量、光合速率、可溶性糖含量等光合性能指标进行了测定,结果表明,两优培九在生育后期光合面积大,叶绿素下降慢,光合速率高,可溶性糖含量多,说明其生育后期的光合能力较强[34]。严斧等[29]也对其生育后期的光合特性进行了研究,结果表明其生育后期具有叶绿素功能期较长和光合速率较稳定的优势。但我们在抽穗期进行的研究表明,其净光合速率、表观量子效率、羧化效率、光与CO2补偿点及饱和点等光合气体交换参数与汕优63、威优46和93-11相比并无优势[35]。因此,作者认为,两优培九之所以能超高产,主要是其生育后期功能叶光合性能强以及优良的光合株型、群体光能利用率高。除了对两优培九的光合特性进行了系统深入研究之外,研究人员对其他超级稻尤其是更高产的超级稻的光合特性也进行了一些研究工作。邓启云等[24]对超级杂交稻理想株型的原形组合培矮64S/E32的光合优势进行了研究,与三系杂交稻汕优63相比,培矮64S/E32抽穗后剑叶和倒二叶的比叶重、后期上三叶叶绿素含量衰减速率、上三叶单位叶面积气孔数目、耐低浓度CO2胁迫能力及抽穗后干物质积累速率等光合相关指标性状优势明显。程式华等[36]对协优9308的研究表明,其始穗后灌浆期剑叶光合速率高于对照协优63,瞬时净光合速率最高值和净光合速率高值持续期也均高于对照。翟虎渠等[37]的研究也表明,协优9308剑叶的光合作用能力显著强于协优63。Chang等[38]比较研究了第二期超级稻代表性组合Y两优1号(YLY1)与两优培九(LYP9)的光合形态与生理指标性状,并探讨了它们与超高产的关系,与LYP9相比,YLY1在光合有效光的高、低光强下,冠层光合作用分别高出5.5%和47.3%,在灌浆阶段,分配到地上的同化物比例平均高出1.5%,籽粒灌浆前叶鞘中储备的光合同化物多4.5%~10.5%,且能更有效地转运,最终有效分蘖数平均增加25.2%。Chang等[39]也比较研究了第四期超级稻代表性组合Y两优900与高产杂交稻汕优63的光合相关指标性状,结果表明,与汕优63相比,Y两优900具有较高的叶绿素含量、较厚的叶片、较大的叶面积、较高的光合速率及更高的有机物质积累和更大的生物量,尤其是在籽粒充实阶段,并认为这些鉴别出的特征可以作为超级稻育种的指标。龙继锐等[40]研究了大田条件下6种施氮水平对超级杂交稻Y两优1号抽穗到蜡熟期剑叶叶绿素荧光动力参数的影响,认为在当时的试验条件下,施氮量为135~180 kg/ha时叶绿素荧光指标组合最优。方宝华等[41]以一、二、三、四、五期超级杂交稻组合两优培九、Y两优1号、Y两优2号、Y两优900及超优1000为材料,用4种典型的光合模型对参试品种齐穗期剑叶光合光响应曲线进行了拟合,并对各品种气孔导度的光响应曲线进行了比较。结果表明,4种光合模型对参试5个品种的光合光响应曲线都可以拟合,决定系数R均大于0.99,其中直角双曲线修正模型拟合得到的光合参数与实测值最接近;光能利用率以超优1000和Y两优900最高,剑叶净光合速率(光合能力)以Y两优900最强,光饱和点以Y两优900和Y两优1号最高,暗呼吸速率以Y两优900和两优培九最大,气孔导度除Y两优2号随光强增加增长缓慢外,其他4个品种均增长较快,但未出现气孔关闭现象,对强光表现出良好适应性。
在超级杂交稻同化物运输与分配特点和生理生化机制方面,报道相对较少。严进明等[42]对重穗型杂种稻光合产物运转特性进行了研究,结果表明,两优培九、Ⅱ优162、特优124等3个重穗型杂种稻与汕优63相比,光合产物的运转效率差,结实率(77.7%~82.9%)都低于汕优63(90.6%)。该研究团队认为,进一步增强现有重穗型杂种稻同化物的运转效率和对外界变化光强的光合适应能力,是实现水稻超高产育种的可能的生理途径。Li等[43]以两优培九和汕优63为材料,通过盆栽试验,进行高氮与低氮栽培,发现低氮可增加腺苷二磷酸-葡萄糖焦磷酸化酶、淀粉合成酶、淀粉分支酶、α-淀粉酶、β-淀粉酶和蔗糖磷酸合酶的活性,导致汕优63中茎淀粉及NSC的积累和再活化增加,认为减少氮肥施用量、选择适当品种可以通过增强茎干NSC的积累和转运来提高水稻籽粒产量,从而降低成本,并促进水稻生产的可持续性。因此,加强现代超级杂交稻叶片同化物运输与分配特点及其机制研究将有助于充分发挥现代超级杂交稻的产量潜力,具有重要研究价值。
综上所述,超级稻在光合生理方面一般具有如下特性:净光合速率高,气孔导度大,CO2补偿点低,光饱和点高,光合色素含量高,光化学量子效率高,RuBP羧化酶含量高,灌浆前叶鞘光合同化物积累多,叶片光合功能期长尤其是后三叶光合功能期长,光合作用对强光、弱光、高温、干旱等逆境的适应性强,等等。
3 展望
虽然在超级稻超高产形成的光合机制方面取得上述重要研究进展,但相关研究也明显存在一些不足。如:这些研究采用的超级杂交稻组合主要是前四期的,而对更高产的第五期超级稻攻关组合超优1000的光合研究鲜见报道。近年来,中国科学院亚热带农业生态研究所夏新界研究员团队与湖南新春农业生物高科技有限公司合作,选育出具有超大生物量的巨型稻种质(图3),但对该材料光合特性的研究未见报道,因此,目前所获得的认识在研究对象上有明显局限性;此外,对个体光合特性的研究多而对群体光合特性的研究少,顺境(适宜环境)下的光合特性研究多而逆境下的光合特性研究较少,对同化物的生产研究多而对其配置、转运特点研究少。因此,未来需要弥补这些方面的不足,并进一步加强对超级稻超高产形成的光合机制研究,以便更好地为水稻的超高产理想株型育种和超高产栽培提供有力理论支撑。目前,我们正在开展相关研究,相信在广大水稻科技工作者的共同不懈努力下,一定能不断深化对超级稻超高产形成的光合机制认识。

图3 巨型稻(徐孟亮摄于湖南省长沙县金井镇湘丰村;左边与中间照片摄于2017年,右边照片摄于2019年。)Fig.3 Giant rice(Taken by Xu Mengliang in Xiangfeng Village,Jinjing Town,Changsha County,Hunan Province;photos at the left and in the middle were taken in 2017,and that in the right was taken in 2019.)
猜你喜欢
作物学报(2022年7期)2022-05-11 12:58:02
农家之友(2018年12期)2018-03-12 00:50:28
中国农业信息(2017年12期)2017-09-03 04:33:01
上海农业学报(2017年4期)2017-04-10 12:40:28
养生保健指南(2016年12期)2017-01-06 22:20:23
上海农业学报(2016年2期)2016-10-27 00:50:01
福建稻麦科技(2015年1期)2016-01-06 12:22:06
发明与创新(2015年37期)2015-02-27 10:40:26
作物研究(2014年6期)2014-03-01 03:39:11
作物研究(2014年6期)2014-03-01 03:39:10
