
我国鸡球虫抗药性现状和抗药机制研究进展*
2021-11-08 06:02:54汤新明吕艳丽于咏兰刘贤勇
寄生虫与医学昆虫学报 2021年2期
李 超 汤新明 吕艳丽 于咏兰 刘贤勇 索 勋**
(1.中国农业大学动物医学院,北京 100193;2.中国农业科学院北京畜牧兽医研究所,北京 100193)
抗球虫药物的使用大大降低了球虫病对养鸡业造成的损失,但耐受各种药物的抗药虫株的不断出现,影响了抗球虫药物的作用效果,球虫病的防控面临新的挑战。自首次报道磺胺喹噁啉的抗药虫株(Waletzky, 1954)后,针对所有投入使用的抗球虫药都出现了抗药虫株。我国自1994年后,在6年之内已经出现了针对12种抗球虫药的抗药虫株(郝振凯等, 2019)。已有一些文献对不同时期的球虫抗药性情况进行了综述(李佩国等, 2011; 彭昊等, 2011; 毕菲菲等, 2018),本文拟对近年来(2011—2021)我国的球虫抗药性现状进行分析,并讨论鸡球虫抗药性产生的机制和应对策略。
1 我国鸡球虫抗药性现状
1.1 文献检索与筛选
以“球虫”、“抗药性”/“耐药性”/“药物敏感性”、“鸡”等组合,在中国知网、万方知识服务平台上共检索到968篇文献,按如下标准进行筛选。纳入标准:(1)报道鸡球虫抗药性调查的文献;(2)采用综合的检测指标(孔繁瑶等, 1994; 黄兵等, 2001),至少应有抗球虫指数(ACI),若没有ACI,则相对卵囊产量(ROP)、病变记分减少率(RLS)和最适抗球虫活性百分数(POAA)3项应均有;(3)采样时间应在2011年及以后、抗药性判定标准应有清晰描述。排除标准:(1)综述、新闻、评论;(2)研究资料重复或与球虫抗药性关联不密切的文章;(3)无具体数据的会议摘要。
关于抗球虫指数ACI的阳性判断标准,在不同文章之间不甚统一(谢明权等, 1999; 陈兆国等, 2001; 黄兵等, 2001; 李亚林, 2014; 王黎霞等, 2014)。综合分析后,本文采用如下标准:ACI≥180,判为敏感;160≤ACI<179,为轻度抗药;140≤ACI<160,为中度抗药;ACI<140,为完全抗药,以便对不同文献进行统一分析。
删除重复文献后,进一步阅读摘要和全文进行筛选,最后纳入32篇文献进行分析。其中,检测柔嫩艾美耳球虫的文章占65.6%,检测混种球虫的文章占28.1%,检测巨型艾美耳球虫的文章占6.3%。在调查区域上,跨多省市的调查有2篇(王黎霞等, 2014; 黄杰等, 2021)其他多是某一或邻近两省市区域内的调查,其中,安徽鸡球虫抗药性研究报道最多,河北次之。
1.2 我国各地区鸡球虫抗药性现状
由于我国各地区养鸡业发展不一致,我们对各省市的调查结果按七大地理区划分别进行分析。总体来说,我国各地区鸡球虫对大部分常用药物均具有中度以上的抗药性,华东和华南地区最严重,西北和东北地区的抗药性最弱(表1~2)。
在各类药物中,对聚醚离子载体类药物和地克珠利的检测最多,而对前者接近一半地区达到完全抗药,对地克珠利基本为中度抗药,对其他化学合成药物过半数达完全抗药。
在跨区域的调查中,2014年在北京、山东、上海的检测显示,柔嫩艾美耳球虫对地克珠利、马杜霉素和莫能菌素均具有中度抗药性(王黎霞等, 2014)。江苏、山东、安徽、福建和河南的13株柔嫩艾美耳球虫对地克珠利和盐霉素具有中度抗药性(黄杰等, 2021)。
1.2.1华东地区:华东地区关于鸡球虫流行病学的报道相当丰富(李超等, 2018)。本地区球虫的抗药性出现较早,并且目前多为中度到完全抗药,耐受多种药物。
柔嫩艾美耳球虫对聚醚离子载体类药物为中度到完全抗药,其中对马杜霉素和莫能菌素在2011—2015年为中度抗药,在2016—2020年则升为完全抗药;对盐霉素在2016—2020年为完全抗药;对海南霉素和拉沙里菌素分别在2011和2020年检测1次,均为完全抗药(王中宝, 2013; 熊苏翔等, 2015; 周飞亚等, 2015; 黄月月, 2017; 黄仪娟等, 2020)。对地克珠利和妥曲珠利平均为中度抗药,但样品中分别有70%和40%为完全抗药(苏子超, 2012; 吴昌标等, 2013;文献同前者不再显示,下同);对磺胺类药物为中度到完全抗药;对氯羟吡啶在2011—2015年为轻度抗药,到2016—2020年升为完全抗药;对癸氧喹酯平均为中度抗药,样品中37.5%为完全抗药;对二硝托胺和氯苯胍均在福建莆田和龙岩各检测1次,对二硝托胺平均为完全抗药,对氯苯胍则平均为中度抗药,样品中有40%为完全抗药;对尼卡巴嗪从中度抗药升为完全抗药;对氨丙啉和常山酮均为中度抗药。
混种艾美耳球虫对聚醚离子载体类、三嗪类、磺胺类药物、癸氧喹酯和氨丙啉为完全抗药,对氯苯胍、尼卡巴嗪和常山酮为中度抗药,基本上也是逐年升高的(孙冰冰, 2014; 蔡兰等, 2015; Lanetal., 2017; 王海霞, 2020)。
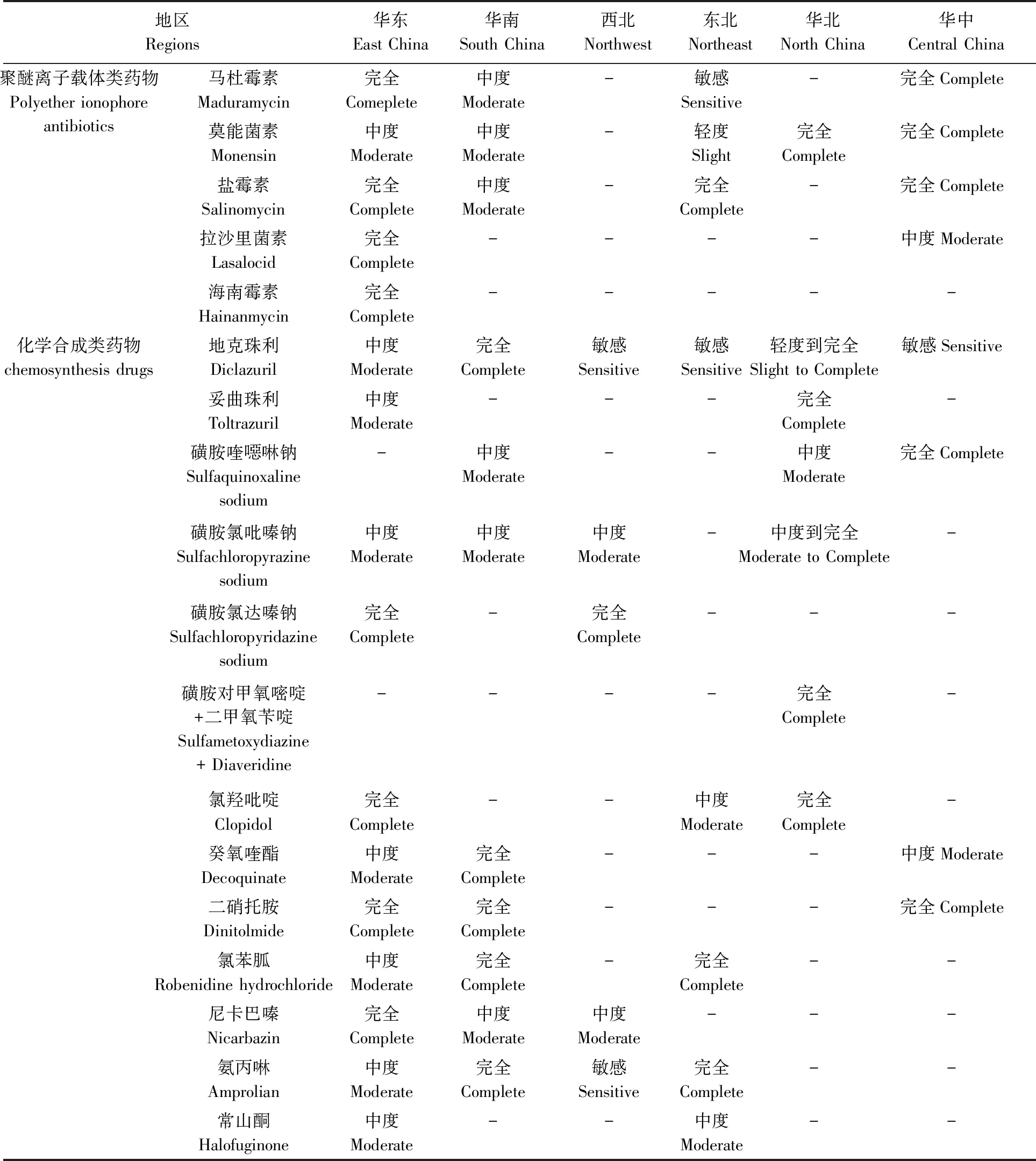
表1 我国各地区柔嫩艾美耳球虫的抗药性(2011-2021)Tab.1 Drug resistance of Eimeria tenella in various regions in China (2011-2021)
1.2.2华南地区:华南地区柔嫩艾美耳球虫对所检测的各种药物为中度到完全抗药(韦园园, 2012; 陈玉婷等, 2013; 易永萍, 2018)。其对马杜霉素和盐霉素为轻度到中度抗药;对地克珠利为完全抗药;对磺胺氯吡嗪钠和磺胺喹噁啉为中度抗药,其中对前者66.7%的样品为完全抗药;对癸氧喹酯、二硝托胺、氯苯胍、氨丙啉均为完全抗药;对尼卡巴嗪为中度抗药。
混种艾美耳球虫对地克珠利、磺胺氯吡嗪钠、磺胺喹噁啉、氯苯胍和氨丙啉完全抗药,对磺胺甲噁唑、尼卡巴嗪和百球清敏感(张艳等, 2012; 谢婷等, 2018)。
1.2.3西南地区:西南地区于2019年报道了1次混种球虫抗药性调查,以中度抗药居多(黄仪娟等, 2019)。柔嫩艾美耳球虫和堆型艾美耳球虫对聚醚离子载体类药物中度抗药,对地克珠利、复方磺胺氯达嗪钠、二硝托胺完全抗药;对氯羟吡啶、癸氧喹酯、氯苯胍、尼卡巴嗪均为中度抗药。可见其抗药谱很广,对各类药物均具有较强的抗药性。
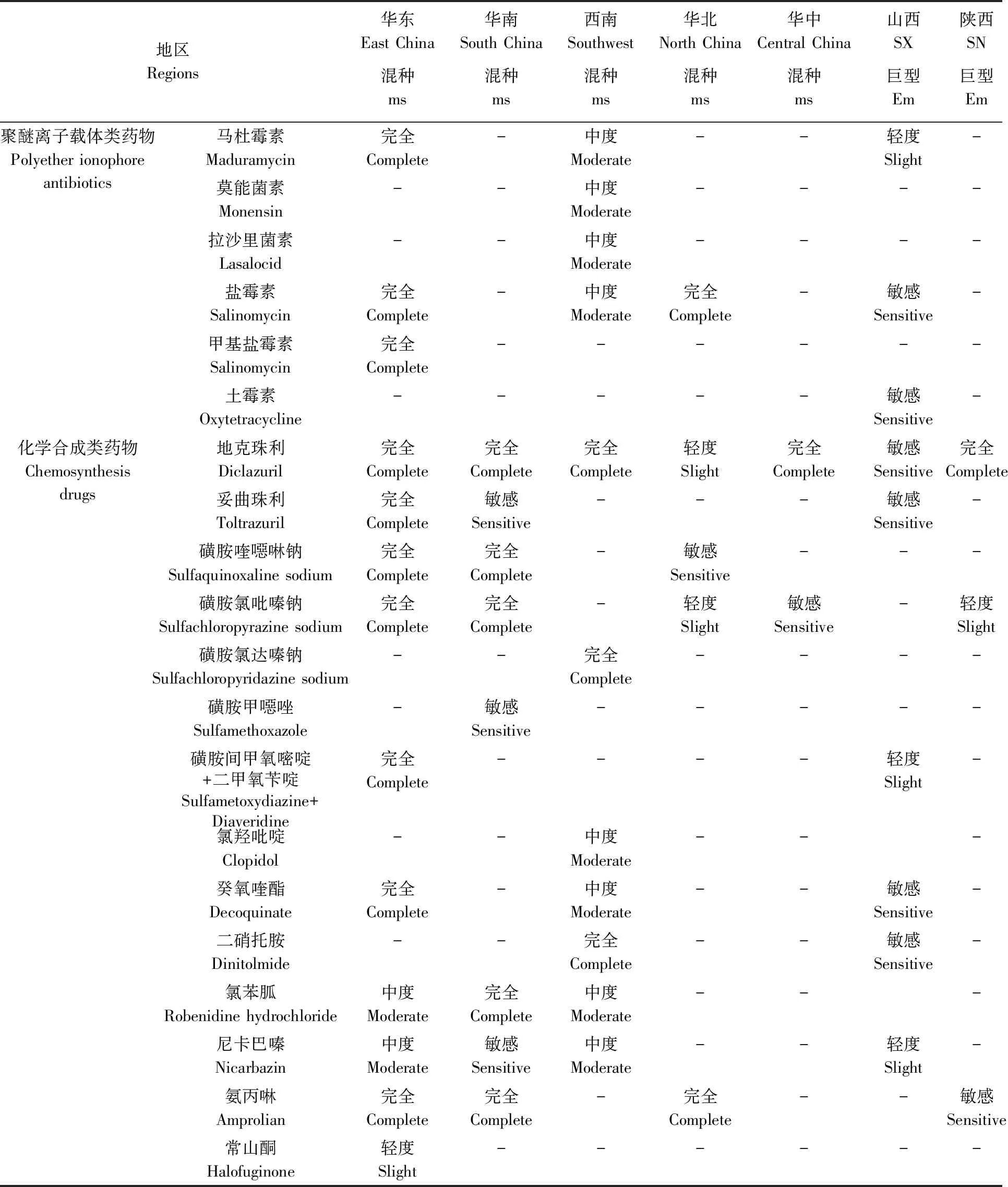
表2 我国各地区混种和巨型艾美耳球虫的抗药性(2011-2021)Tab.2 Drug resistance of E. maxima (Em) or multi-species (ms) in various regions in China (2011-2021)
1.2.4西北地区:西北地区近年来球虫抗药性报道也较少,且抗药性程度较低,基本为轻度抗药到中度抗药。青海果洛柔嫩艾美耳球虫对聚醚离子载体类药物的敏感性未检测,对地克珠利、氨丙啉敏感,对磺胺氯吡嗪钠、尼卡巴嗪为中度抗药;对磺胺氯达嗪钠为完全抗药(刘成录等, 2011)。陕西杨凌巨型艾美耳球虫对地克珠利同样为完全抗药,对磺胺氯吡嗪钠轻度抗药,对氨丙啉敏感(丛湄湄等, 2012)。
1.2.5东北地区:东北地区在2011年于吉林长春报道过1次球虫抗药性检测,其对不同药物的敏感性呈多样化,这不同于前几个地区在整体上较为一致的情况。柔嫩艾美耳球虫对聚醚离子载体类药物从敏感到完全抗药不等;对地克珠利敏感,对氯羟吡啶和常山酮中度抗药,对氯苯胍和氨丙啉完全抗药(姚新华等, 2011)。
1.2.6华北地区:华北地区球虫抗药性较强,在河北和北京之间存在一定的差异。在聚醚离子载体类药物中,本地区仅检测了柔嫩艾美耳球虫对莫能菌素的敏感性,为完全抗药;对地克珠利,河北各虫株平均为轻度抗药,而北京株则为完全抗药;对磺胺氯吡嗪钠,河北各虫株平均为中度抗药,北京株为完全抗药;对其他药物则一致,对妥曲珠利、磺胺对甲氧嘧啶+二甲氧苄啶、氯羟吡啶均为完全抗药;对磺胺喹噁啉钠为中度抗药(孟令慧等, 2011; 蒋禄峰等, 2015; 王静等, 2018)。可见,柔嫩艾美耳球虫对各种药物的抗药性比较强。
与此相反,混种球虫对各种药物的抗药性较弱,除了对盐霉素、氨丙啉为完全抗药外,其对地克珠利、磺胺氯吡嗪钠为轻度抗药,对磺胺喹噁啉钠敏感(赵京媛, 2014)。类似地,山西巨型艾美耳球虫仅对少数药物表现出弱抗药性,对马杜霉素、尼卡巴嗪和磺胺间甲氧嘧啶+二甲氧苄啶为轻度抗药,对地克珠利、妥曲珠利、土霉素、盐霉素、二硝托胺、癸氧喹酯均敏感(张雪松等, 2018)。
1.2.7华中地区:华中地区球虫抗药性报告较少,检测药物也不多,仅有ACI指数作为判定,整体显示出比较强的抗药性。除拉沙里菌素外,柔嫩艾美耳球虫对聚醚离子载体类药物为完全抗药,在化学合成药物中,其对地克珠利敏感,对癸氧喹酯为中度抗药,对磺胺喹噁啉钠、二硝托胺为完全抗药(李亚林, 2014; 刘瑞良, 2018)。
2017—2018年,两次在湖北与河南的大样本量调查显示,混种球虫对地克珠利有完全抗药性,对磺胺氯吡嗪钠中度抗药的虫株占16.88%,轻度抗药的虫株占20.78%(雷振宇, 2018; 叶成, 2019)。
2 球虫抗药性机制研究
2.1 抗药虫株的获得
除了分离田间抗药虫株,在实验室中培育抗药虫株也是研究球虫抗药性常采用的手段(Cuckleretal., 1955)。常用的实验室诱导方法有2种:(1)Weppelman 法,Weppelman等进行大剂量卵囊感染试验,成功筛选到胺喹甲酯的抗药虫株,但未获得对其他药物如氨丙啉、氯苯胍和莫能菌素的抗药虫株(Weppelmanetal., 1977)。(2)药物浓度递增法,以低于推荐使用剂量的药物浓度开始诱导,逐步提高浓度,多次继代后,获得抗药虫株,是抗药性研究中普遍采用的一种方法。如柔嫩艾美耳球虫莫能菌素(Chapman, 1984)、马杜霉素(安健等, 2006)、地克珠利和马杜霉素(韩红玉等, 2004)、盐霉素抗药株(王海霞等, 2019)都是利用这种方法获得的。不同抗药虫株的起始诱导浓度和传代次数不同(郝振凯等, 2019)。
2.2 常见抗球虫药物抗药机制研究
常见的抗球虫药物主要有聚醚离子载体药物和化学合成类药物。不同药物的对球虫的作用机理不同,因此其抗药性产生的机制也会不一样,可能涉及超微结构如细胞膜的离子通道、物质代谢如糖类代谢或能量代谢如细胞色素氧化酶。
Jeffers设想球虫抗药性的产生是因虫体基因发生的突变(Jeffers, 1974)。那么这种突变体是球虫群体本来就有,还是在诱导过程中产生的呢?Williams指出有些球虫个体在从未接触过的抗球虫药作用之下仍然能够生存,因此提出球虫存在“固有抗药性”(Williams, 1972)。Weppelman对胺喹甲酯突变体的选择(Weppelmanetal., 1977)、Williams对癸氧喹酯抗药虫株的选择成功(Williams, 1998)都证明了球虫群体中确实存在抗药性的突变体。也有学者认为抗药性是对存在于球虫群体中突变体定向选择的结果(Chapman, 1984),其根据在于应用Weppelman法无法获得对常山酮的抗药虫株,但运用浓度递增法能诱导出这种抗药虫株(Chapman, 1986)。
2.2.1聚醚离子载体类药物:包括莫能菌素、马杜霉素、盐霉素、拉沙里菌素等。它们的作用机制相似,主要作用于无性生殖阶段,可扰乱虫体内的阳离子浓度,进而破坏正常的渗透压(Chapmanetal., 2010)。抗药虫株的出现可能是由于基因突变,或某些基因表达的变化,导致虫体内的结构蛋白或代谢酶等发生改变,使细胞壁结构改变或离子通道通透性发生变化,使虫体内部渗透压在药物作用时,仍能处于平衡状态,同时减少药物向内运输。
柔嫩艾美耳球虫对本类药物抗药性主要集中在马杜霉素和莫能菌素的研究上。前期主要在探索抗药虫株和敏感虫株的差异蛋白条带,涉及表面抗原与代谢酶等。如莫能菌素抗药虫株比敏感株多了两条蛋白;盐霉素、拉沙里菌素抗药虫株的乳酸脱氢酶酶谱比敏感虫株少1条带;马杜拉霉素抗药株与敏感株的差异蛋白质之一为子孢子表面抗原TA4(Zhuetal., 1994; 余丽芸等, 1999; 姜连连等, 2006)。
随着分子生物学的发展,在特异序列和抗药相关基因方面的研究越来越多,且差异基因多集中在入侵相关基因和代谢酶基因特别是能量代谢酶上。马杜霉素抗药株表达量上调较高的6个EST序列中,有2个和半胱氨酸蛋白酶同源性较高;其孢子化卵囊18 S rDNA基因有3个碱基发生突变;mRNA的4条差异序列中,有1条与MIC5高度同源(韩红玉等, 2005; 安健等, 2006; 蔡兰等, 2006)。在差异基因表达方面,一些入侵相关基因如MIC-1上调表达,而一些能量代谢相关基因如NAD(P)转氢酶下调表达(Chenetal., 2008)。近期的转录组学研究表明,一些差异基因和糖酵解/糖异生、肌动蛋白细胞骨架的调控和DNA复制等过程相关;抗药株表面抗原(SAG10)表达下调,而苹果酸脱氢酶(MDH)上调表达(王海霞等, 2019; Xieetal., 2020)。
莫能菌素抗药虫株的研究也类似,莫能菌素抗药虫株中有25.8%的蛋白表达发生变化(25/97),其中肌动蛋白明显上调表达,而MIC4则显著下调表达;一些骨架蛋白和代谢相关酶如色氨酸合酶基因上调表达(Chenetal., 2008; Thabetetal., 2017)。另有研究表明,莫能菌素会引发虫体细胞自噬,自噬途径的破坏有可能导致抗药性出现(Qietal., 2020)。
2.2.2三嗪类:主要有地克珠利和妥曲珠利,其对有性生殖和无性生殖阶段均有抑制作用。该类药物可抑制虫体的多种生理功能,如抑制细胞核分裂;使内质网膨胀,破坏空间结构;抑制呼吸关键酶活性,影响能量代谢(Harderetal., 1989; Wangetal., 2013; Zhouetal., 2013; Shenetal., 2014)。
柔嫩艾美耳球虫地克株利抗药株的5个差异蛋白质斑点中,有2个蛋白质分别为球虫子孢子表面抗原TA4和热休克蛋白Hsp70(姜连连等, 2005)。转录组测序结果显示,一些差异基因和过氧化物酶体,不饱和脂肪酸的生物合成以及脂肪酸代谢等过程相关;并且第2代裂殖子的顶膜抗原-1、海藻糖-6-磷酸合酶、胸苷酸激酶假定蛋白等16个基因与敏感株有明显差异(韩贞艳, 2018; Xieetal., 2020)。与马杜霉素抗药株相似,地克珠利抗药株的表面抗原(SAG10)也发生下调表达,而苹果酸脱氢酶(MDH)上调表达(王海霞等, 2019)。
可见,球虫对三嗪类产生抗药性的原因涉及多个方面,可能包括与入侵相关的表面抗原的基因突变或表达量改变、脂肪酸代谢及三羧酸循环。
2.2.3磺胺类药物:磺胺类药物主要作用于裂殖生殖阶段,机理是抑制二氢叶酸合成酶的活性,阻止二氢叶酸合成,进而干扰叶酸正常代谢,抑制球虫繁殖。参考疟原虫和弓形虫的研究,虫体对磺胺类药物产生抗药性的原因,可能是二氢叶酸还原酶编码基因发生突变,导致药物与酶的结合能力下降。
疟原虫对乙胺嘧啶抗药虫株的二氢叶酸还原酶基因发生突变,突变位置接近酶的活性位点,可能会影响虫体与药物的结合(Petersonetal., 1988)。同样地,弓形虫二氢叶酸还原酶的突变也导致其对乙胺嘧啶产生抗药性(Reynoldsetal., 2001)。
2.2.4癸氧喹酯和氯羟吡啶:这两种药物都可以阻断球虫线粒体内的电子转运过程,从而抑制球虫细胞呼吸作用,影响能量代谢。前者主要作用于子孢子和第1 代裂殖子生殖阶段;后者则主要作用于第2代裂殖子和配子体。虫体对药物的吸收和聚集作用减弱可能会带来其对癸氧喹酯抗药(Wang, 1975)。抗癸氧喹酯的疟原虫虫株在细胞色素b基因上存在的2个突变位点(Nametal., 2011),会降低虫体对该药物敏感性(Tanetal., 2017)。氯羟吡啶的作用位点与癸氧喹酯不同,二者不存在交叉抗药性(Fryetal., 1984),球虫对其产生抗药性的机制还有待探索。其他常见药物如二硝托胺、氯苯胍、尼卡巴嗪、氨丙啉和常山酮的抗药性机理则仍有待深入研究。
3 结论及展望
1994—2021年,我国各地区柔嫩艾美耳球虫对大部分常用药物的抗药性越来越强,且均具有中度以上的抗药性,而华东和华南地区更严重,具有完全抗药性。在文章梳理中,我们发现鸡球虫抗药性的调查缺乏系统性,不利于清楚判断当前球虫抗药性的实际情况。但因球虫抗药性的监测比较繁琐和昂贵,费时费力,单靠养禽企业、科研院所难以持续进行,建议由畜牧兽医行政部门牵头并有配套制度和资金支持,进行定期监测,方能制定科学的用药方案。另一方面,加强球虫抗药性机制产生的基础研究投入,挖掘抗性基因位点,建立高效快速的检测方法也是球虫抗药性研究亟需突破的瓶颈。
当前已有研究表明,球虫抗药性和入侵相关抗原及多种代谢酶密切相关。虽然近年来基因组、转录组和蛋白质组学技术的应用有力促进了鸡球虫抗药性的研究,但目前其机制仍未能得到透彻阐释。近期随着球虫基因编辑技术建立(Tangetal., 2020),为球虫抗药性产生机制的解析提供了有力的技术支撑。
目前形势及养殖需求模式下,需要采取综合措施防控鸡球虫病:(1)进行定期监测,及时掌握球虫抗药性变化趋势;(2)针对当前的抗药性情况,合理用药,如进行联合用药、穿梭用药、轮换用药、尝试使用停用很久的老药、应用中草药等;(3)加强环境清洁工作,如及时处理鸡舍粪便,彻底消毒,切断球虫的繁殖周期;(4)加强饲养管理,合理控制鸡舍温湿度和饲养密度,调整日粮营养配比;(5)疫苗免疫和药物控制交替或轮换使用,最大限度降低疫苗的副作用和消减抗药虫株产生的频率。
猜你喜欢
河南畜牧兽医(2020年23期)2020-01-10 03:19:52
世界农药(2019年3期)2019-09-10 07:04:08
农药科学与管理(2019年10期)2019-04-20 07:13:08
武警医学(2018年10期)2018-11-06 07:04:34
新农业(2017年2期)2017-11-06 01:02:23
畜牧与饲料科学(2016年9期)2016-11-01 21:07:43
湖南农业(2016年12期)2016-03-10 12:48:45
杂草学报(2015年2期)2016-01-04 14:57:53
中国预防兽医学报(2015年10期)2015-03-09 05:40:50
无机化学学报(2014年3期)2014-02-28 17:30:55
