
北京市居民区中蚊虫密度与气象因素关系的研究*
2021-11-08 06:02:44刘美德李秋红刘尤焕苓周小洁叶彩华曾晓芃
寄生虫与医学昆虫学报 2021年2期
刘美德 姜 江 佟 颖** 张 勇 阎 婷 李秋红刘 婷 尤焕苓 周小洁 叶彩华 曾晓芃
(1.北京市预防医学研究中心,北京市疾病预防控制中心,北京 100013;2.北京市气象服务中心,北京 100089)
蚊虫的生长、繁殖、行为以及疾病的传播等各个方面受到气象因素的影响是非常明显的,因此气象因素对蚊虫密度的影响和相关预测研究一直备受关注(Davidetal., 2002; Patzetal., 2005; Guillermoetal., 2005)。就影响蚊虫的气象因素而言,多数的研究结果认为对蚊虫密度、蚊虫活动性影响最大的气象因素就是气温,其他气象因素则在不同区域和时间段内存在着一些争议和差别,原因在于各个地域的主导气象因素的不同(王金娜等, 2015),或者同一气象因素在不同环境、不同时期也可能对蚊虫密度产生完全不同的影响(周毅彬等, 2010; 杨维芳等, 2010; 于德宪等, 2010; 景晓等, 2011; 仲洁等, 2015;李海龙等, 2015)。
城市环境中的居民区往往是城市居民生活高度集中的地方,既是城市人群密度大之处,也是蚊虫与人类接触机率最高的区域,蚊虫对人群的叮咬骚扰与蚊虫在人群中传播蚊媒疾病往往更多的是发生类似居民区这种人群聚居的区域中,所以研究城市生活区中蚊虫的密度及分布动态对于在城市环境中控制蚊虫密度、减少叮咬骚扰、降低可能蚊传疾病的机率至关重要。由于气象因素与蚊虫密度间的密切关系,如果能够利用气象因素开展蚊虫密度及分布的预测(Luisetal., 2012; Yunetal., 2014),所得结果可以很好地作为减少城市居民被蚊虫叮咬的机率或甚至降低蚊传疾病的传播概率。因此,本文运用北京各行政城区气象数据与相应区域的居民区蚊虫密度,详尽分析了较长时间序列中城区、近郊区、远郊区的蚊虫密度变化规律,及其与气象条件的关系,并采用多元回归分析进行了基于气象因素的蚊虫密度预测,所得结果对于做好北京城市区域中蚊虫及其传播疾病的防控工作有着重要的意义。
1 材料与方法
1.1 数据资料
本文所用气象资料来自于北京市气象局的逐旬气象数据,涉及要素包括2009—2019年逐旬平均气温、旬内最高气温、旬内最低气温、旬内降雨量、旬平均相对湿度、旬平均风速、旬平均气压。涉及14个站点:顺义54398站、海淀54399站、延庆54406站、密云54416站、怀柔54419站、平谷54424站、通州54431站、朝阳54433站、昌平54499站、门头沟54505站、大兴54594站、丰台54514站、房山54596站、石景山54513站。
就北京行政区划而言,应有16个区,分别为北京市区(城六区):东城区、西城区、朝阳区、海淀区、丰台区、石景山区;北京近郊区:大兴区、通州区、顺义区、昌平区、门头沟区、房山区;北京远郊区:怀柔区、平谷区、密云区、延庆区。但是城六区中的东城区和西城区没有建设国家级气象自动站,为了保证数据来源统一有效,在本文分析过程中未分析东城区和西城区,市区仅依赖朝阳区、海淀区、丰台区、石景山区的监测结果。
北京市的蚊虫多出现在每年的5~10月,因此,北京市疾病预防控制中心(CDC)每年5~10月都会采用CO2诱蚊灯(北京隆冠科技发展有限公司,型号: MT-1型)法获得蚊虫密度监测数据(单位为:只/(灯·小时))。本研究的蚊虫密度数据均来源于北京市疾病预防控制中心,区域涉及包括顺义、海淀、延庆、密云、怀柔、平谷、通州、朝阳、昌平、门头沟、大兴、丰台、房山、石景山14个城区的居民区;时间涉及2009—2019年;每年5~10月居民区环境点,采用CO2诱蚊灯法获得蚊虫密度监测数据(共198旬蚊虫密度数据),全市共设置109个采样点,每个点设置4台CO2,除了CO2做为引诱剂外不再添加别的引诱物。
1.2 研究方法
研究选择了研究中应用较多的多元回归方法,具体方法解释如下:(1)多元回归方法:多元回归方法用来研究一个因变量与多个自变量之间的关系问题。具体公式如下:yj=β0+β1x1+β2x2+···+βmxm+εj,式中β0,β1,...,βm均为模型的回归系数,εj为随机残差。
(2)检验方法:为客观的评价多元线性回归分析得到的预报差异,选用均方根误差来检验预报效果。
2 结果
2.1 蚊虫指数变化规律
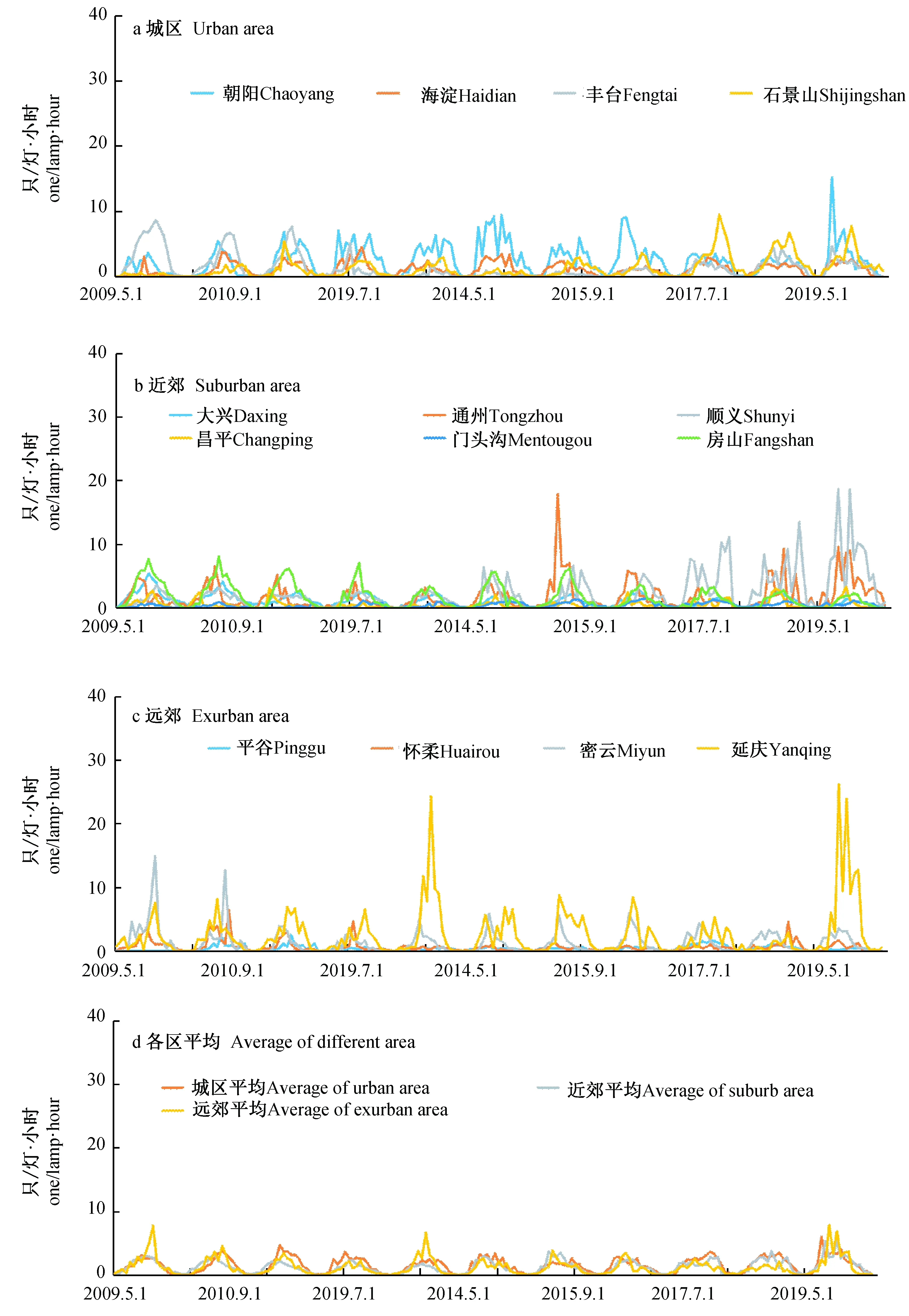
图1 2009—2019年北京市及各区蚊虫密度居民区监测数据(只/(灯·小时))Fig. 1 Mosquito density in residential areas of different districts of Beijing from 2009 to 2019 (individuals/(lamp·hour))
2009—2019年14个区各区平均居民区的蚊虫密度分析结果显示(图1),在每年的5~10月之间,居民区的蚊虫密度都呈现出周期性的波动,居民区内蚊虫密度高峰值集中出现在7月中旬到8月中旬,这个时期也正是北京地区气温最高和降水最集中的时期。十余年来,城区居民区内的蚊虫密度相对比较稳定,而近郊和远郊城区的居民区内(如通州、顺义、延庆)会在个别年份出现极大值(为其他同期测站数值的几倍或者几十倍),最大值出现在延庆区的2019年7月上旬,为26.13只/(灯·小时),最小值为0只/(灯·小时)。
另外,2009—2019年5~10月城区居民区内平均蚊虫密度依次要高于近郊和远郊,相应的数值分别为1.50、1.26和1.20只/(灯·小时)(图2)。
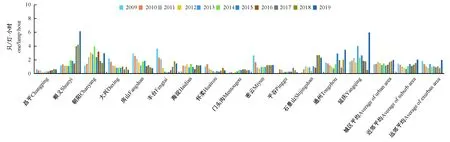
图2 2009-2019年北京各区居民区监测点多年平均蚊虫密度监测数据(只/(灯·小时))Fig. 2 Annual average monitoring data of mosquito density in residential areas of different districts in Beijing from 2009 to 2019
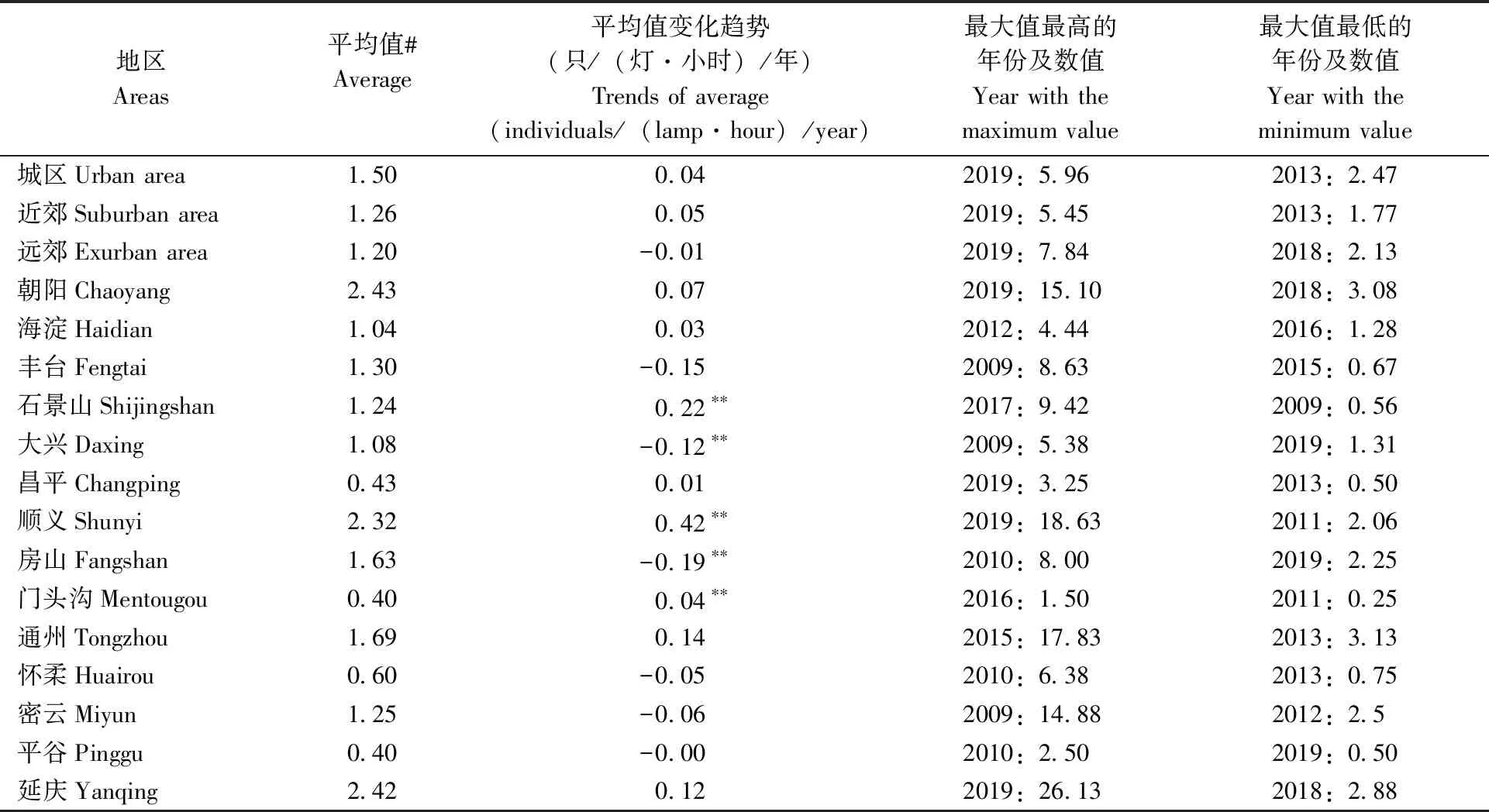
表1 2009—2019年5—10月各区居民区监测点平均蚊虫密度Tab.1 The average mosquito density in residential areas of different districts in Beijing from 2009 to 2019
5~10月城区、近郊和远郊区蚊虫密度最高的年份均出现在2019年,但是具体就各个区域而言,由于植被分布、地形等因素的差异,各个观测点并未表现出最高/低值集中出现在某一年份(表1, 图2)。
2009—2019年间年平均居民区内蚊虫密度变化趋势在多数地区表现为增加,丰台、大兴、房山、怀柔、密云、平谷和远郊表现为下降,变化趋势通过95%的显著性检验(T检验)的仅有石景山、大兴、顺义、房山和门头沟,分别为0.22、-0.12、0.42、0.04只/(灯·小时)/年,其余具体数值见表1。研究者之前的研究中也发现,受全球气候变暖的影响,北京地区整体的蚊虫密度整体处于上升状态(刘美德等, 2019)。
2.2 蚊虫指数与气象因子间的关系
在蚊虫与气象因素的关系方面,蚊虫的生长繁殖发育需要一定的气温条件,气温过低易导致发育减慢。气温过高,会导致成虫过小,存活率降低(罗成旺等, 2007);降雨则可以增加蚊虫的孳生地数量,使幼虫大量繁殖(易彬樘等, 2003);而相应的降雨带来的湿度增加也会影响蚊虫生长繁殖发育(Glenn, 2008);此外,气压对蚊虫密度的影响是一种负相关关系,并且具有一定的滞后效应(代培芳等, 2011);这些都是本研究中气象因素与城市居民区中关系的生态学机制。
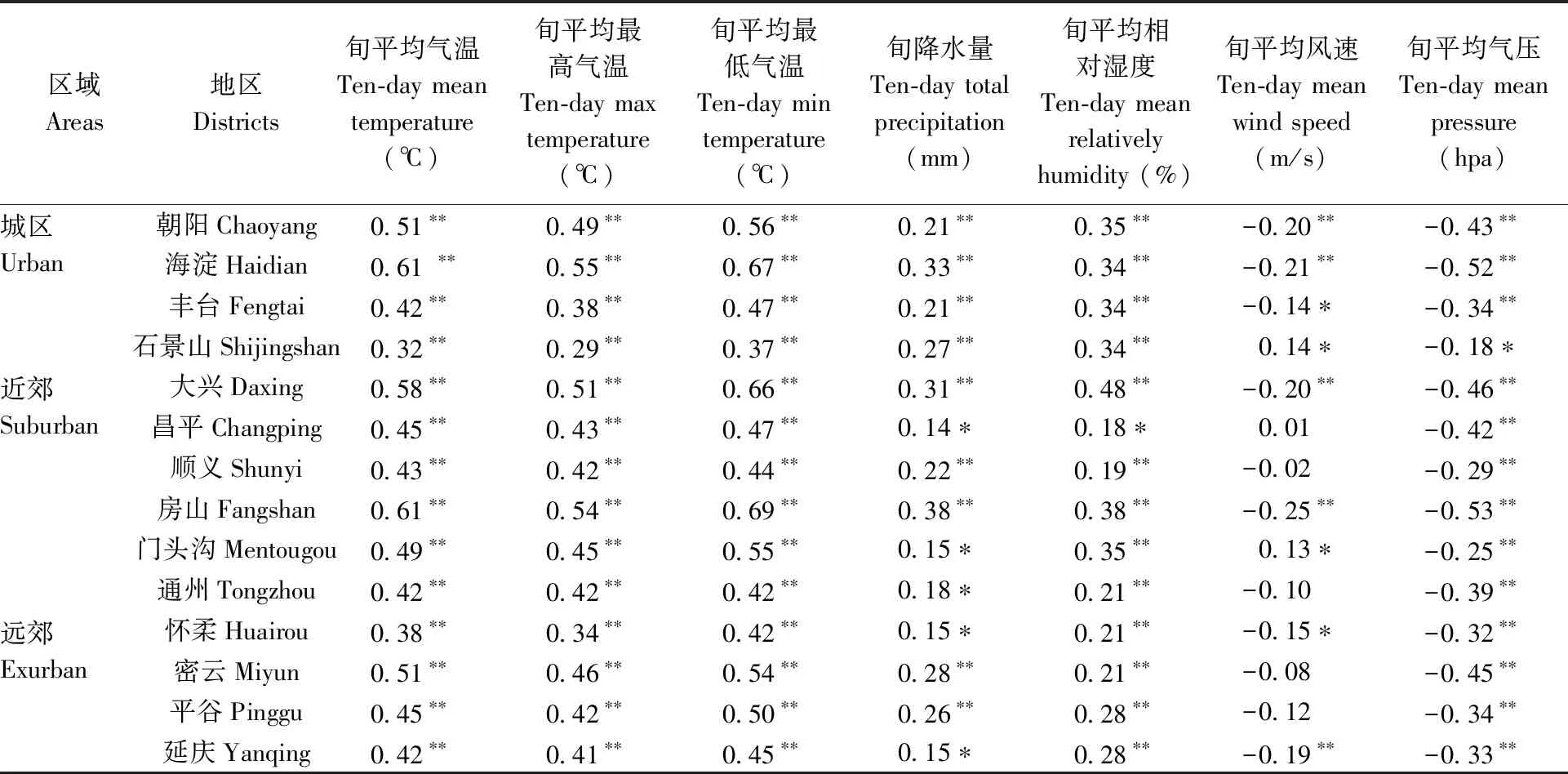
表2 气象要素与蚊虫指数的相关系数Tab.2 Correlation coefficient of meteorological factors to mosquito density
多数的研究认为在众多气象要素中,对蚊虫影响最大的就是气温。而其他因素则在不同区域、不同时间上表现出地域的差别。另外,已有的研究表明北京地区蚊虫密度相关的气象因素主要为最低气温、相对湿度和风速(赵瑶等, 2009)。基于2009—2019年长时间序列、多区域的居民区监测点蚊虫密度数据的分析结果表明,在北京居民区内蚊虫密度相关性从高到底的气象要素分别是旬平均最低气温、旬平均气温、旬最高气温、旬平均气压、旬平均相对湿度、旬降水量以及旬平均风速,且均通过了相关性的显著检验,其中旬平均气压和旬平均风速呈现负相关关系,其余要素都是正相关关系(表2)。
2.3 多元线性回归的蚊虫指数预测
除肯定气温、湿度与风速对居民区蚊虫密度的影响外,本研究还发现其他气象因素如:气压、降水量也与居民区蚊虫密度密切相关,这可能增加对蚊虫密度预测的可能性与精确性。北京市蚊虫密度高峰集中在7月中下旬到8月中旬,对应该地区最高气温和最大降水集中期;但考虑到降雨量对蚊虫密度影响复杂性且每一次降雨前后的天气状况都存在差别,现阶段无法获取日值的蚊虫密度数据,因而,无法进行更加细致的分析。
在前文旬数据相关分析的基础上,本文选择了旬平均最高气温、旬平均最低气温、旬平均气压、旬平均相对湿度以及旬平均风速5个重要气象要素来进行旬蚊虫密度预测。X1:旬平均最高气温;X2:旬平均最低气温;X3:旬平均相对湿度;X4:旬平均风速;X5:旬平均气压。利用2009—2018年180旬的数据做训练集,2019年18旬的数据做预测检验,基于X1-X55个气象要素所得到的预测结果均方根误差在0.224~8.218之间(表3)。
相对而言,居民区是蚊虫密度相对比较稳定的区域,预测效果更好,在城区、近郊和远郊在个别年份出现极大值或波动较大的观测站点,如朝阳、通州、顺义、延庆的蚊虫密度预测效果都较差(图3)。

表3 基于5个气象要素多元线性回归预测蚊虫密度Tab.3 Prediction of mosquitoe densities by multiple linear regression based on 5 meteorological factors

表4 加入前旬蚊虫密度多元线性回归预测蚊虫密度Tab.4 Prediction of mosquitoes’ densities by multiple linear regression with the baseline densities ten days before
考虑蚊虫密度在相连两旬之间会存在一些关联。在预测时,尝试将“X6:前旬蚊虫密度”滑入到多元回归方程中,预测效果得到了明显的提升,均方根误差在0.180~7.383之间。除朝阳区之外,相较于仅利用气象要素预测的均方根误差减少了9.82%~62.29%。
同样,因蚊虫密度相对比较稳定的各站,加入前旬蚊虫密度之后预测效果提升也更大,但是如朝阳、通州、顺义、延庆的蚊虫密度在个别年份有极大值出现或数值波动较大,即使加入了前旬的蚊虫密度也未能显著提升预测效果(图4)。
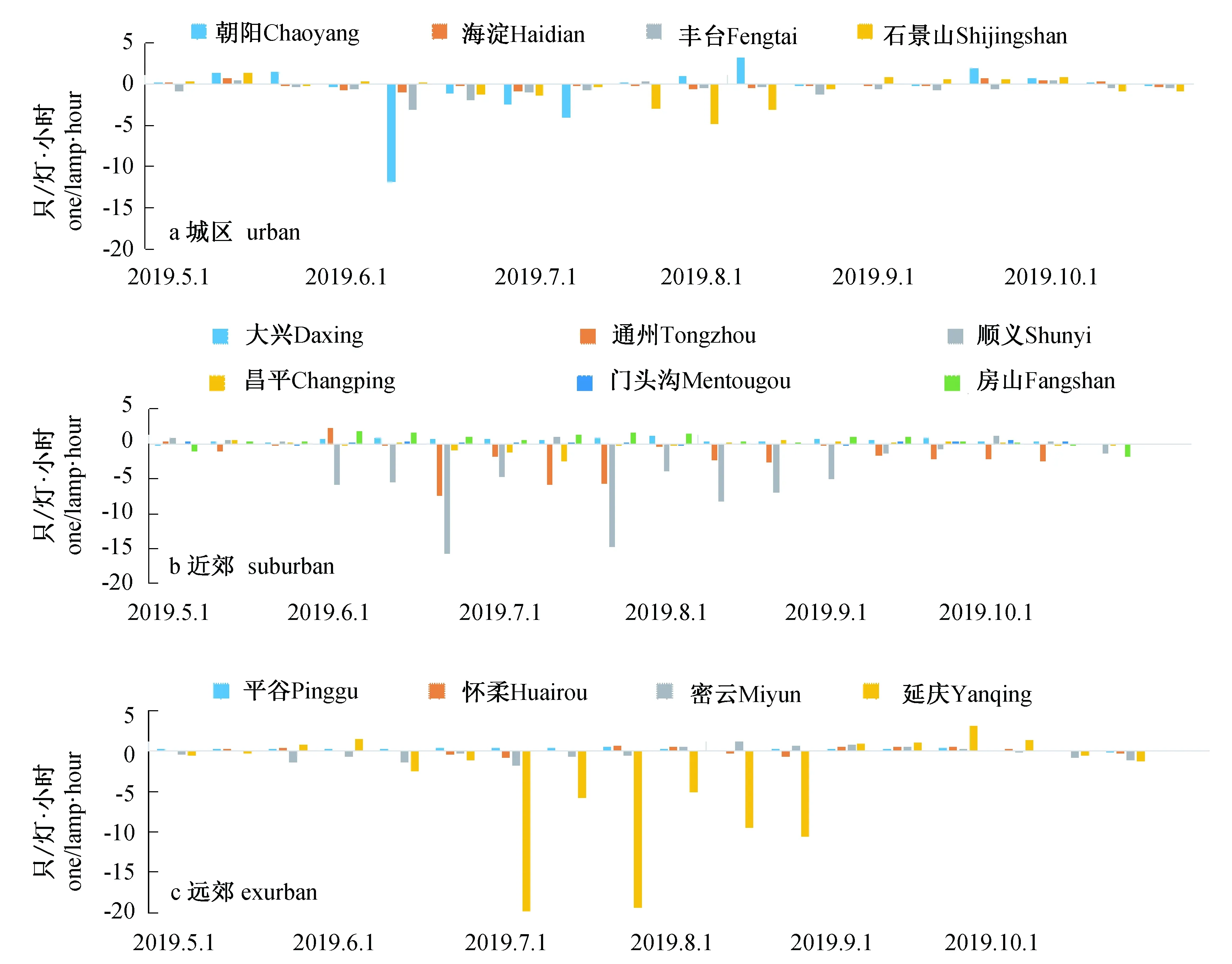
图3 2019年5—10月居民区监测点蚊虫密度预测值与真实值的差异(5要素预测)Fig. 3 Difference between predicted and observed mosquito densities in residential areas in Beijing from May to October, 2019 (based on 5-factor)
图4 2019年5-10月18旬居民区监测点蚊虫密度预测值与真实值的差异(6要素预测)Fig. 4 Difference between predicted and observed mosquito densities in residential areas in Beijing from May to October, 2019 (based on 6-factor)
3 讨论
3.1 北京不同城区居民区蚊虫密度的变化
研究发现2009—2019年5—10月城区居民区的平均蚊虫密度高于近郊和远郊,近郊也比远郊地区蚊虫密度高。这可能归因于卫生条件差,蚊虫的孳生地增加。在本研究中,却是发现城区生活区中的蚊虫密度比近郊和远郊地区的蚊虫密度要高,这可能是城市化对蚊虫密度的影响引起的原因。严杰等(2017)发现城市化中景观变化与蚊虫密度分布有密切的关系,蚊虫密度与温度因素(旬平均最低气温、旬平均气温、旬最高气温)密切相关。城市热岛效应可能是城市核心区居民区蚊虫密度要高于近郊或远郊区域的生态学原因。
3.2 北京各城区居民区蚊虫密度变化趋势
研究发现2009—2019年间北京各区域居民区的平均蚊虫密度整体呈上升趋势,说明在城市居民区亚环境中,蚊虫密度也会受到气候变暖的影响,需要持续做好居民区蚊虫密度监测和控制工作。同时,还需注意的是气温的升高可能会扩大蚊媒在空间和时间上的分布范围,缩短病毒等病原体在蚊虫体内的潜伏期,加快传播速率(Brower, 2001;Ramasamyetal., 2012)。随着全球气候变暖的进程,城市居民区蚊虫密度上升极有可能增加蚊媒疾病暴发的风险,高度重视气候变化对于城市居民区内蚊虫密度影响十分必要。石景山、大兴、顺义、房山和门头沟等部分居民区蚊虫密度呈下降趋势,这可能和这些地区爱卫工作(张勇等, 2018)的推进、居民区卫生条件的改善、蚊虫孳生减少相关(孙红专等, 2000)。
3.3 北京市居民区蚊虫密度的预测分析
本研究建立的多元线性回归模型结果表明,气象因素的合理应用可以对蚊虫密度进行比较准确的预测,在此基础上滑入前期蚊虫基线密度会有效提升预测的精度,进而实现城市居民区蚊虫密度及其传播疾病风险的精准预测,最大程度地减少蚊虫对居民区内人群的叮咬机率与传播疾病的风险。同时,也必需注意个别年份出现极大值或波动较大的区域,比如本研究中如朝阳、通州、顺义、延庆等,居民区蚊虫密度预测效果就比较差。这可能与这些区域环境变化太大有关,对此,突破线性回归方法的局限性,应用机器学习或深度学习等人工智能方法提高预测模型的智能化程度和精确度十分必要。
猜你喜欢
小学生学习指导(爆笑校园)(2022年9期)2022-09-30 06:49:16
成都信息工程大学学报(2022年3期)2022-07-21 09:35:50
保健医苑(2022年6期)2022-07-08 01:26:16
今日农业(2021年12期)2021-11-28 15:49:26
今日农业(2021年2期)2021-03-19 08:36:38
畅谈(2018年17期)2018-10-28 12:30:46
中学生博览(2016年17期)2016-09-10 13:59:26
广东第二课堂·小学(2016年9期)2016-05-14 19:18:22
IT时代周刊(2015年9期)2015-11-11 05:51:37
小雪花·成长指南(2015年10期)2015-10-23 08:52:46
