
组合抗体库技术的研究进展
2021-10-27 08:25:10强敏施晓疌张楚悦马培翔杨光
自然杂志 2021年5期
强敏,施晓疌,张楚悦,马培翔,杨光
上海科技大学 免疫化学研究所,上海 201210
组合抗体库(combinatorial antibody library)技术是现代免疫化学的重要组成部分[1-2]。它是一种构建大容量、高多样性的文库,并利用生物进化原理进行优化重组和淘筛的技术。其特征是通过组合策略获得丰富的多样性,将基因型和表型有机整合以实现表型的可复制,并在一个可筛选的体系中将符合所需表型的分子淘筛出来。这一技术可以从B细胞中克隆抗体重链和轻链的基因序列,在体外重组构建抗体基因文库,其多样性高达1011~1012[3]。建立组合抗体库的基本原理是模拟获得性免疫系统的特征,通过基因重排和筛选的策略获得适合的抗体。组合抗体库的生物学特征包括:①可在试管中构建多样性达1011的合成免疫系统;②模拟天然免疫系统基因型与表型相联系的复制系统;③减少传统免疫抗体获得时对活体动物的依赖;④规避免疫耐受问题,可获得识别自身抗原的抗体;⑤包含供者终生完整抗体反应的所有记录;⑥可揭示一些罕见的特殊抗体谱。根据组合抗体库的大小,通过核糖体载体、噬菌体载体、酵母载体、细菌载体、细胞载体等的表面展示抗体,其中噬菌体载体可展示的库容量最大[4]。此外,将抗体分泌表达在细胞质中、锚定在细胞膜上或滞留在内质网膜上等技术,可以进一步拓展组合库的应用范围。根据表面展示抗体形式的不同,组合抗体库分为Fab抗体库、单链抗体(scFv)库和纳米抗体(nanobody)库等。
细胞展示技术巧妙地将组合库抗体的基因型与表型有机整合在一起,而细胞所承载的生物功能决定了可采用的筛选方法[5]。噬菌体库的筛选原理是基于亲和力进行淘筛的。大多数情况下,研究人员希望筛选获得的抗体也具有靶点调控功能。由于噬菌体的宿主大肠杆菌缺乏蛋白质翻译后修饰系统,而后者是发挥生物功能的关键之一,因此噬菌体系统可用的筛选策略受到限制。针对这一问题,科研人员对筛选体系进行了改造:在哺乳动物细胞中同时表达功能报告系统,开发出基于单细胞自分泌[6-7]和基于细胞-细胞相互作用的组合库筛选体系[8-10]。组合抗体库技术的发展历程如图1所示。
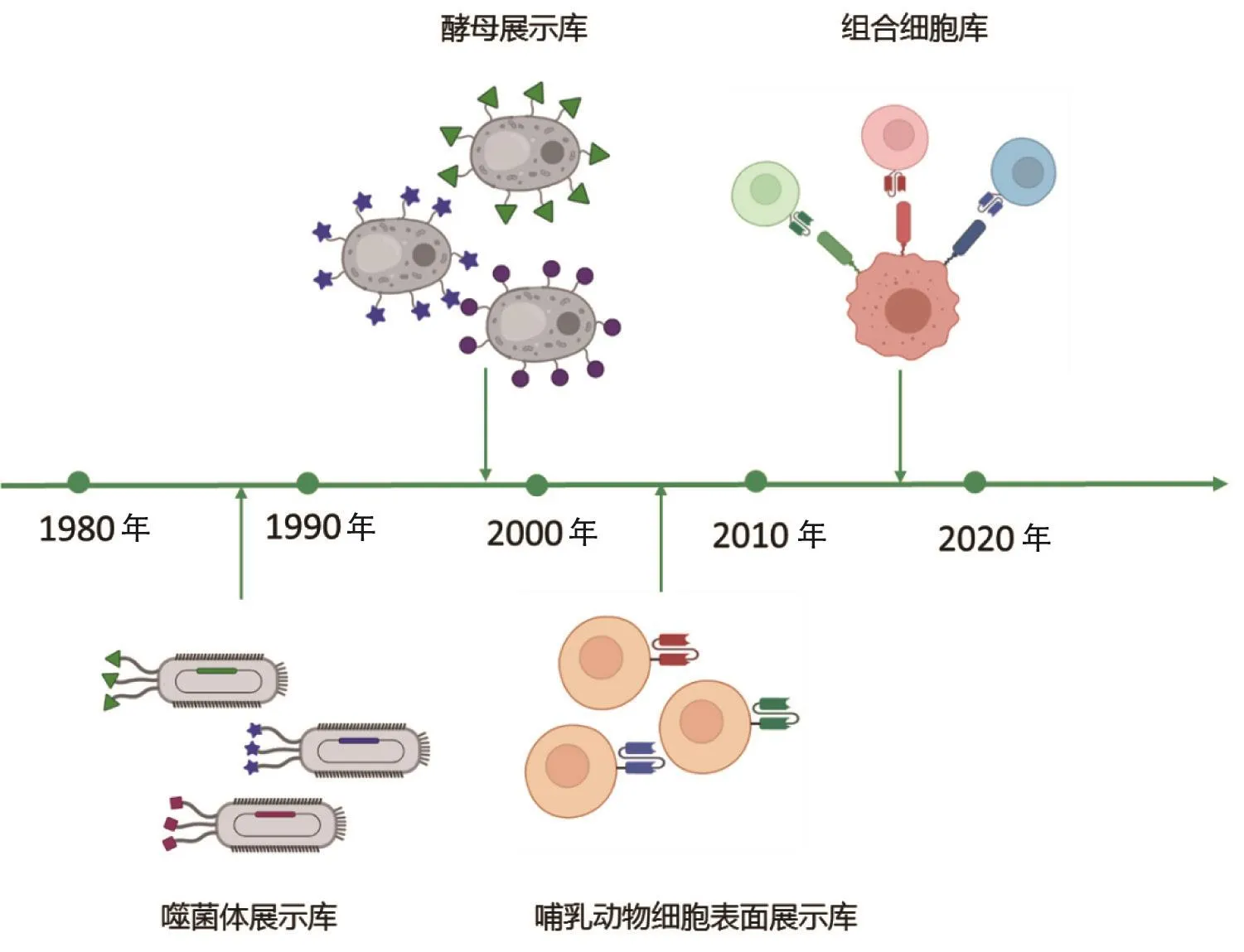
图1 组合抗体库技术的发展历程
1 基于亲和力的组合抗体库筛选系统
基于亲和力的组合抗体筛选系统包括噬菌体展示抗体库和酵母表面展示抗体库。噬菌体展示抗体库是体外筛选特定抗原高亲和力抗体最常用的技术(图2)。其原理为:从免疫个体或健康个体的B淋巴细胞中扩增全套抗体的重链和轻链mRNA或RNA文库,经噬菌体表面展示系统表达后形成噬菌体文库,即噬菌体抗体库(phage antibody library)。1985年George P.Smith[11]首次描述噬菌体展示技术,并证实丝状噬菌体可用于展示多肽。1989年Richard A.Lerner实验室报道首个噬菌体展示抗体库,构建了展示于λ噬菌体表面的鼠源组合Fab抗体库,其多样性与小鼠的天然免疫系统相当甚至更高,并从中鉴定出功能性抗体[12]。1990年Gregory Winter实验室报道了一个基于scFv的噬菌体展示库,也证实从噬菌体群体中筛选出抗体的可行性[13]。噬菌体抗体库可用于结合被固定的抗原做筛选,筛选富集的噬菌体经测序可获得对应的抗体序列。因为噬菌体颗粒的尺寸较小且高度可溶 (1013个/mL),所以噬菌体库的多样性可达1011,这有利于在体外筛选全人源序列的抗体。目前已有超过70种噬菌体来源的抗体进入临床研究,14种已获批准[14]。如今在获美国食品及药品管理局(FDA)批准的100多种抗体中,包括销售额最高的阿达木单抗等,至少有11种是通过组合抗体库技术发现的(表1)[5]。2018年,组合抗体库技术获得诺贝尔化学奖。

表1 FDA批准的来源于组合抗体库的抗体类药物[5]
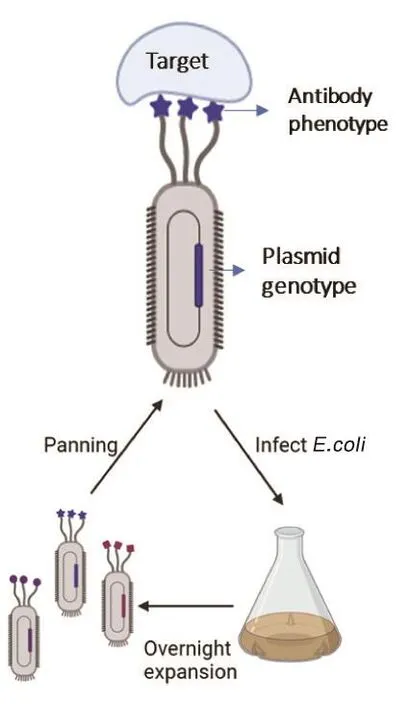
图2 基于亲和力的筛选原理图
噬菌体库筛选得到的治疗性抗体可用于癌症、炎症、感染及免疫性疾病的治疗[14],这些抗体靶向的是针对细胞命运、细胞信号通路、病毒信号通路或GPCR等细胞膜和胞外的蛋白。此外,也有关于胞内表达抗体筛选的技术和基于细胞膜呈递抗原的技术被报道[4,28]。
离子通道是多种神经性、心血管性、代谢性疾病,以及癌症和免疫调节的重要药物靶点,它在可兴奋性组织和非兴奋性组织中都发挥了关键作用。这类靶点的生物药以功能性的毒素肽和抗体为主,它们识别与作用的位点是离子通道的胞外区域[29]。但是,筛选对离子通道亚型具有选择性和特异性的药物很有挑战。通过技术的优化,噬菌体组合抗体库技术针对酸敏感离子通道1a(acid-sensing ion channel 1a,ASIC1a)及连接蛋白26(connexin 26,Cx26)成功筛选到了高特异性的抗体。
酸敏感离子通道(ASICs)是受质子调控的一类非电压敏感型阳离子通道,其中ASIC1a是缺血性卒中治疗的一个极有潜力的靶点[30]。Qiang等[31]通过表达人源ASIC1a的膜蛋白并将其组装至纳米磷脂盘(nanodisc)中获得模拟该离子通道保持天然构象的hASIC1a-nanodisc复合物,以此为抗原进行噬菌体库的抗体筛选。使用多样性达1011的全人源噬菌体抗体库进行了3轮筛选,获得数条富集的scFv抗体序列,其中ASC06抗体具有特异性识别细胞表面表达的ASIC1a的特点。细胞水平的研究显示,该ASC06-IgG抗体可浓度依赖地抑制ASIC1a通道的电流,且能挽救酸中毒引起的细胞死亡。对脑中动脉阻塞模型小鼠,缺血1小时再灌注2小时后注射ASC06-IgG,可使小鼠的脑梗死体积显著下降(约42%)。ASC06-IgG显示出强效的神经保护作用。制备ASIC1a与ASC06抗体的复合物时,负染电镜结果显示该抗体以1:1的计量比结合在一个ASIC1a亚基上,即每个三聚体组成的ASIC1a通道上可结合3个ASC06抗体,这与其他毒素肽的结合模式类似[31]。此抗体具有良好的应用前景。
连接蛋白(connexin,Cx)是组成细胞间隙连接通道(gap junction channel,GJC)的关键蛋白之一[32],它的错义点突变会形成泄露的半通道(hemi channel,HC),引起角膜炎-鱼鳞病-耳聋(KID)综合征等疾病[33-34]。Xu等[35]针对Cx26进行了抗体筛选,他们以人源Cx26蛋白胞外第一段环形结构域多肽为抗原,使用噬菌体抗体库进行筛选,获得了特异结合细胞表面Cx26的scFv-Fc抗体AbEC1.1。后续研究发现该抗体是一种完全可逆的Cx26 HCs抑制剂,且不影响GJCs的功能。更重要的是,该抗体能有效抑制Cx26 HCs突变体(G45E、D50N)引起的病理性过度活跃[35]。晶体结构显示该抗体会组装为双链抗体(diabody)。使用分子动力学模拟和半通道电流记录的组合,进一步鉴定了该抗体的表位,即两个AbEC1.1形成的双链抗体结合在一个Cx半通道的顶部,像盖子一样阻塞该半通道的孔道并抑制其开放,能减缓因其泄露引起的表型。研究还发现该抗体还可特异结合与Cx26具有相似第一段环形胞外域的Cx30,且抑制Cx30 A88V的半通道泄露和ATP过度释放,并减轻Cx30 A88V突变小鼠(人类Clouston综合征模型)的皮脂腺增大、皮肤过度增生等异常症状[36]。
除了新抗体的筛选,抗体的亲和力成熟是基于亲和力筛选系统的另一项重要应用。抗体和抗原结合的强度称为亲和力,它可以用解离常数KD进行表征。亲和力是评价抗体性质的重要参数。在生物个体内,抗体对抗原的高亲和力意味着强有力的识别,可更高效地激活免疫系统,清除病原体。体内B细胞作为获得性免疫的重要部分,在首次接触一种病原抗原后会产生一定量的抗原特异性IgM和IgG类型抗体,而一段时间后当相同抗原的二次免疫发生时,相应B细胞基因组上的抗体序列将发生体细胞高频突变,并且这些细胞在二级淋巴器官生发中心进行高亲和力突变克隆的克隆选择,保留并扩增亲和力更好的B细胞克隆,完成抗体的体内亲和力成熟,亲和力往往可以达到数个数量级的提升。最后这些被选择的细胞将进一步分化成为浆细胞和记忆细胞。
对于抗体药物而言,亲和力特征是直接影响药效、稳定性甚至安全性的一个重要参数。通常情况下,人们希望获得的抗体候选药物具有高亲和力,因此,高亲和力抗体往往具有更大的成药潜能。进入临床研究阶段的抗体候选药物的亲和力大多已经强于纳摩尔级别,如靶向TNFα的adalimumab的KD为30 pmol/L、靶向PD-1的pembrolizumab的KD为27 pmol/L、靶向IL-5的mepolizumab的KD约为100 pmol/L等。初筛抗体亲和力不高时,需要额外的方法和步骤对所获抗体进行优化,此步骤被称为抗体的体外亲和力成熟。
抗体体外亲和力成熟的基本原理是模仿抗体的体内亲和力成熟过程。对于已知抗体,利用点突变(定点突变和随机诱变)、CDR区重组、链替换以及DNA重组等方法,在已有序列上进行多样性扩增建成二级库,筛选获取更优亲和力的候选抗体[37]。亲和力成熟的筛选方法,基本原理是优化抗体和抗原相互作用时的动力学特征,主要是对抗体的解离速率进行优化。通过增加解离孵育时间、降低抗原结合浓度、引入抗原竞争性结合物等方法或者几种方法的组合,从已结合的抗体中筛选出结合强度更好的抗体[38]。
体外亲和力成熟使用较多的是酵母展示及淘筛技术[39-40]。酵母相较于噬菌体,拥有更接近于哺乳动物细胞的蛋白翻译后修饰机制,而这对于亲和力可能产生重要影响。酵母展示的Fab抗体更接近于天然抗体,可有效解决一些情况下scFv库筛选获得的抗体转化成全长IgG后的效力损失问题[41]。此外,酵母尺寸远大于噬菌体,可利用流式细胞仪分选,筛选过程便能可视化地区分不同强度结合状态的克隆,并任意设置筛选阈值,非常有利于亲和力成熟的筛选。
Shi等[42]在对噬菌体组合抗体库筛选后获得了抑制CXC基序趋化因子受体2(CXC-motif chemokine receptor 2,CXCR2)的候选抗体,然而在面对体内CXCR2高亲和力(纳摩尔级别KD值)、且局部高浓度的天然配体IL-8时,此抑制剂亲和力不足,使得无法有效竞争IL-8,从而使其不具备足够的药物开发潜力。因此,他们在抗体表位识别中起到关键作用的重链互补决定区3(H-CDR3)进行随机突变,筛选酵母展示的突变文库。为了不使突变程度过大而远离设定靶点的识别,研究人员采用基于核酸序列的每个碱基30%随机突变概率,最终获得皮摩尔级别亲和力的抑制型抗体,在动物体内的炎症模型验证中显示出极强的中性粒细胞抑制功能和炎症消退功能。
Tao等[43]利用噬菌体库对瘦素受体(LepR)筛选靶向抗体,初筛获得两个具有激动剂功能的scFv抗体。虽然具有新型的激动剂模式,然而它们对LepR的激活能力弱于它的天然配体(瘦素),因此使用酵母展示突变库筛选的方法优化候选抗体亲和力。他们设计了一种基于氨基酸序列的随机饱和点突变策略,首先控制突变范围为重链CDR3区域随机的4个氨基酸残基,然后每个密码子采用NNK突变策略,从而实现向20个氨基酸密码子的饱和突变,可产生2×107变体多样性,最终成功地获得了7个亲和力大幅提高的突变体,在磷酸化报告系统试验和受体信号相关的细胞增殖试验中均展现出优于天然配体(瘦素)的性质。
在可成药靶点中,尤其是对于通常不能跨膜进入细胞内的抗体药物而言,膜受体如GPCR、离子通道、膜受体酪氨酸激酶、各种转运受体等占据着重要的位置。在抗体亲和力成熟的过程中,任何影响抗体-抗原相互作用的干扰因素都不可忽视,例如线性表位和靶点天然构象间的差异。因此,膜受体相对复杂的胞外域结构、配体结合的多样性,不仅给有效的抗原纯化带来了困难,而且纯化的抗原与天然抗原的差异也成为其功能性抗体亲和力成熟中一大干扰因素[44-45]。针对这个问题,Yang等[9]开发了基于酵母-细胞相互作用的亲和力成熟体系,利用酵母展示Fab抗体库,对表达在细胞表面的受体成功地进行了亲和力成熟。与对于全细胞展示的抗原筛选方法不同的是,Yang等的策略结合了细胞-细胞相互作用的特性,并利用流式细胞分选技术,在已有结合的群体中差异性地筛出高亲和力抗体,有效地解决了非特异性的问题[46-47]。
抗体亲和力成熟的筛选过程不仅可用于优化抗体的亲和力,基于筛选设计的选择方向,同样可以用抗体识别选择性的优化。例如,在抗体初始的筛选过程中,出于治疗应用的目的往往选择人源受体作为筛选抗原,使得所获抗体可能具有种属特异性。但这可能对于之后验证抗体效果的动物实验带来困扰。对于已经获得的特异性识别人源抗原的抗体,可构建突变的酵母展示抗体库,用人源靶点和鼠源靶点进行交叉筛选,最终获得可交叉识别人和鼠靶点的新抗体,如LepR激动型抗体和CXCR2抑制性抗体,为之后的临床前动物研究铺平道路[43]。值得注意的是,这样的方法能够同时在抗体的特异性和亲和力方面获得优化。
2 基于细胞自分泌的组合抗体库筛选系统
由于噬菌体宿主大肠杆菌的局限性,为了拓展功能抗体的筛选策略,部分课题组将靶抗原和对应其功能的报告系统同时表达在同一株细胞中,用于抗体筛选。例如,为了使阳性筛选表型与相应抗体的基因型相偶联,Lerner等[6]创立了一套细胞自分泌筛选系统(图3),即用上述带有靶点报告系统的细胞展示抗体库,展示的抗体作用于自身细胞上的靶点,产生阳性信号的细胞被分选富集并获取胞内相应抗体的DNA序列,这一体系的原理类似于细胞分泌的因子作用于自身受体的自分泌效应。
自分泌筛选系统的主要目的是直接筛选功能性抗体。Tao等[43]根据LepR的胞内段可以介导瘦素激活受体后引起的JAK-STAT信号通路响应的原理, 构建了STAT3磷酸化激活响应激活报告系统,包括GFP、luciferase和β-lactamase,再通过转座子系统等方法构建了此报告系统和LepR过表达的稳转筛选细胞株[43]。另外,研究人员还构建了长程响应LepR激活的增殖相关联的Ba/F3筛选细胞系。接着他们采用慢病毒转导,将LepR预富集的抗体库展示在这些构建好的功能筛选细胞上,结合FACS技术和增殖筛选方法,筛选获得了具有激动剂功能的抗体。最后,研究人员在体外各种信号检测模型中鉴定了抗体的功能,并在肥胖小鼠的体内模型中,验证了此抗体通过激活LepR产生节食、代谢调控的减肥功能。
Xie等[6,48-49]利用自分泌功能筛选,针对粒细胞集落刺激因子受体(GCSF-R)也成功筛选获得了激动型抗体。更有意思的是,他们发现此激动型抗体可以促使人骨髓造血干细胞向神经前体细胞的分化,展现了通过功能筛选获得抗体,可能具有天然配体所不具备的受体多效性激活能力。这种情况同样在利用功能筛选系统获得的促血小板生成因子受体(TPO-R)的激动型抗体上出现,其可以诱导急性髓系白血病病人的癌细胞向自然杀伤性细胞(NK cell)分化,并反过来杀伤其他白血病细胞[50]。针对红细胞生成素受体(EPO-R)建立自分泌筛选系统,筛选其激动剂,最终成功获得激动型抗体[51]。这些功能都是受体相应的天然配体所没有的,展现了功能性抗体筛选在临床上拓展应用的巨大价值。
功能筛选的另一个优势是,可以进行靶点未知的非偏向性功能筛选。为获取具有诱导干细胞定向分化能力的功能性抗体,Lerner实验室研究人员以幼红细胞系TF1为筛选细胞,慢病毒转导展示抗体库,并在凝胶克隆形成实验中根据细胞形态筛选找到可以使细胞克隆分化至树突状细胞的功能性抗体,经鉴定其靶点为细胞整合素[52]。值得注意的是,在此筛选工作前并未预设抗体功能和靶点,是完全非偏向性的筛选,此方式适用于新靶点和新功能的筛选。在实验室另一个抗病毒作用抗体的筛选工作中,研究人员同样在未预设靶点的情况下,筛选得到可以保护HeLa-H1细胞免受鼻病毒感染致死的功能抗体,随后鉴定其靶点为病毒的3C蛋白酶[53]。该工作证明此靶点作为抗病毒药物靶点的可行性和重要性,另外还展现了此方法直接将靶点验证和药物筛选合二为一的应用潜能。2017年,同样运用这种非偏向性自分泌功能筛选系统,Lerner实验室和合作者共同报道了能够替代转录因子Sox2诱导成纤维细胞重编程至iPS细胞的功能性抗体,随后鉴定其靶点为细胞表面蛋白Basp1[54]。此工作不仅提供了新的重编程相关靶点,也将重编程诱导从细胞核内的转录因子向细胞外信号转变进了一步,具有重要的学术和应用双重价值。
3 基于细胞-细胞相互作用的组合抗体库筛选系统
细胞相互作用是生物功能的基础,而细胞相互作用的基础就是蛋白与蛋白的相互作用。新型的免疫细胞药物,正是通过免疫细胞表面的受体和肿瘤细胞表面的配体结合进行识别,因此利用细胞-细胞相互作用进行免疫细胞药物的筛选将极大地推动新型免疫细胞药物的开发。嵌合抗原受体T细胞(CAR-T)免疫疗法是免疫细胞疗法中较为成功、使用较为广泛的疗法。临床试验中,针对B细胞表面抗原CD19的CAR-T用于治疗血液瘤产生了显著的疗效[55-56],靶向成熟B细胞抗原(BCMA)的CAR-T疗法也对多发性骨髓瘤产生了显著的疗效[57]。2017年8月30日,FDA批准首个CAR-T疗法Kymriah上市,用于治疗难治或复发的急性淋巴细胞白血病(ALL)。截至2021年4月,全球已经有5款CAR-T产品上市。但是,CAR-T治疗仍然存在着一些问题,例如on-target/off-tumor毒性、细胞因子风暴(CRS)、神经毒性等[58]。
在整个CAR-T免疫疗法中,嵌合抗原受体(CAR)是一个非常重要的组成部分。CAR是表达在T细胞膜上模块化的融合蛋白,用于识别抗原,通常是由胞外识别区、跨膜区和胞内信号区,包括CD3ζ和共刺激分子(例如CD28、CD137等)组成[59]。胞外识别区识别肿瘤细胞表面的抗原,引导T细胞识别肿瘤靶细胞,常用单链抗体(scFv)作为胞外识别区。肿瘤表面抗原分肿瘤特异性抗原(TSA)和肿瘤相关抗原(TAA),TSA非常少,绝大部分可用抗原为TAA。TAA在正常组织细胞中也有表达,这就使T细胞会对正常的组织细胞进行杀伤,从而产生on-target/off-tumor的脱靶效应[60]。例如,针对肾细胞癌的第一代碳酸酐酶Ⅸ(CAⅨ)的CAR-T治疗中,由于CAⅨ在正常胃黏膜、小肠细胞上低表达,脱靶效应引起自身免疫性胆管炎和严重的肝损伤[61]。
针对脱靶问题,Ma等[8]开发了一个利用细胞间相互作用进行筛选的组合CAR细胞库(CCC)策略(图4)。该CCC文库在细胞中整合了以单链抗体库为基础的CAR文库,细胞携带有报告基因,可通过细胞间的相互作用直接进行CAR筛选。在该研究中,通过CCC分别与CD38低表达的正常细胞和CD38高表达的肿瘤细胞孵育进行负向和正向差异化筛选,获得多条有效CAR序列,其中包括优选序列CAR-RP02。虽然CARRP02的亲和力弱于传统方法得到的CAR-028,但是装载了CAR-RP02的CAR-T细胞无论是在对肿瘤细胞的特异性杀伤还是细胞因子分泌等方面均优于CAR-028。CAR-RP02具有更低的脱靶效应,当RP02-CAR-T细胞和028-CAR-T细胞分别与低表达CD38的正常细胞共孵育后,RP02-CAR-T细胞基本被CD38低表达的正常细胞激活,且不杀伤这些细胞;而高亲和力的028-CAR-T细胞清除了大部分的CD38低表达的正常细胞。通过CD38的mRNA定量研究发现,RP02和028区分肿瘤细胞和正常细胞的关键因素是CD38在细胞中的表达量。结构表征进一步证明RP02在CD38上的结合靶位亦不同于028。所构建的RP02-CAR-T细胞在小鼠的体内抑瘤实验中显示良好的治疗效果。该研究为解决CAR-T开发过程中所面临的特异性靶点不足、无法区分肿瘤/正常细胞所造成的脱靶效应,以及免疫细胞治疗中的免疫耐受问题提供了一种新的解决方案。
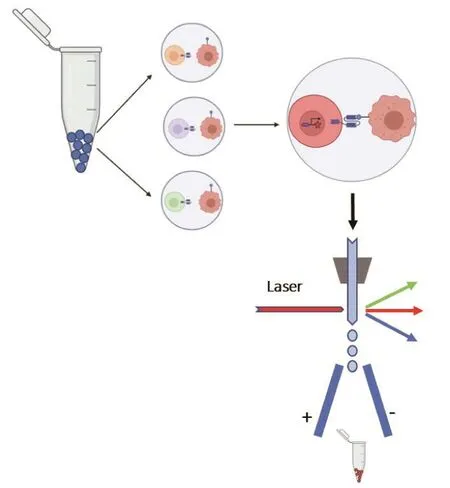
图4 基于细胞-细胞相互作用的筛选原理图
除了单一的细胞-细胞相互作用,组合库还可以在更为复杂的体系中发挥作用。Zheng等[10]对噬菌体展示技术进行改良,他们利用微液滴构建了一个小的生态系统库,每个液滴中都含有一个展示抗体的噬菌体和报告细胞,如果抗体具有功能性,报告细胞将表达荧光,再通过分选、测序等方法得到候选抗体。这一系统首次证实,组合库可用于复杂生态系统的功能筛选。
4 总结和展望
组合抗体库技术针对特定靶标,构建大容量、高多样性的文库进行淘筛,已成为一种重要的药物发现手段。随着新型药物形式的出现,组合库技术也需要不断改进。目前组合库技术已经从噬菌体文库、酵母表面展示文库发展到哺乳动物细胞表面展示文库。由于展示系统的宿主复杂性的提高,能模拟的生物学功能更复杂,所能筛选的药物的种类也更多,因此这些高级的宿主未来能够承载药物形式更复杂的文库。从筛选策略上看,组合库从抗体和抗原的亲和力筛选,发展到单一细胞的自分泌筛选系统、细胞-细胞互作的筛选系统和迷你生态系统的筛选系统,这些筛选系统将在实体瘤等更为复杂的疾病的药物开发中发挥更重要的作用。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:20
中老年保健(2021年3期)2021-12-03 02:32:25
中国动物传染病学报(2021年3期)2021-07-21 03:19:28
中国生殖健康(2020年7期)2020-12-10 07:48:51
新闻传播(2018年11期)2018-08-29 08:15:30
新闻传播(2018年13期)2018-08-29 01:06:52
新闻传播(2016年9期)2016-09-26 12:20:34
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
新闻传播(2015年7期)2015-07-18 11:09:57
医学研究杂志(2015年7期)2015-06-22 11:01:36
