知母皂苷B-Ⅱ通过调控LncRNA XLOC_032768减轻LPS诱导心肌细胞损伤
2021-10-26 12:06薛兴翠孙军奎贺连栋
中成药 2021年10期
薛兴翠, 孙军奎, 贺连栋
(青海红十字医院全科医学科,青海 西宁 810000)
急性心力衰竭是临床常见的一种心血管疾病,其中感染性内毒素血症是引发心力衰竭的重要原因之一,而感染性内毒素血症与革兰氏阴性菌有关,脂多糖(LPS)是革兰氏阴性菌细胞壁外膜上的重要致病因素,可诱导心肌组织损伤从而引发心力衰竭[1-2]。天然植物提取物可能通过调控微小RNA(miRNA)表达从而减轻LPS诱导心肌细胞损伤,长链非编码RNA(LncRNA)在心肌细胞损伤中表达异常并可能作为治疗的潜在靶点[3-4]。
知母为我国传统中药,知母皂苷B-Ⅱ(Timosaponin B-Ⅱ,TB-Ⅱ)是其主要活性成分,对异丙肾上腺素诱发的大鼠心肌梗死的心脏发挥保护作用[5],但该成分对LPS诱导心肌细胞损伤的作用机制尚未阐明。长链非编码RNAXLOC_032768(LncRNAXLOC_032768)在顺铂诱导的肾小管上皮细胞中表达水平降低,上调其表达可减轻肾小管上皮细胞炎症反应及抑制细胞凋亡[6],但它在知母皂苷B-Ⅱ影响LPS诱导心肌细胞损伤过程中的作用机制尚不明确。因此,本研究采用LPS诱导心肌细胞建立细胞损伤模型,探讨知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡、炎症、氧化应激的影响及对XLOC_032768的调控作用。
1 材料与方法
1.1 材料 知母皂苷B-Ⅱ购自上海莼试生物技术有限公司(粉末状,纯度≥98%,溶于DMSO中,批号20171203)。大鼠心肌细胞H9C2购自广州吉妮欧生物科技有限公司;LPS购自北京寰宇集团有限公司;DMEM培养基、胎牛血清购自美国HyClone公司;胰蛋白酶购自美国Gibco公司;Lipofectamine2000购自美国Invitrogen公司;Annexin V-FITC/PI双染法细胞凋亡检测试剂盒购自美国Sigma公司;GSH-Px、SOD检测试剂盒购自南京建成生物工程研究所;si-NC、si-XLOC_032768转染质粒购自上海吉玛制药技术有限公司;MDA检测试剂盒购自碧云天生物技术研究所;NO检测试剂盒购自上海岚派生物科技有限公司;TNF-α、PGE2检测试剂盒购自上海仁捷生物科技有限公司;Trizol试剂购自北京全式金生物技术有限公司;反转录与荧光定量检测试剂盒购自美国Thermo Fisher公司;兔抗鼠Bcl-2、Bax、cleaved caspase-3抗体购自美国Santa Cruz公司;HRP标记的山羊抗兔IgG二抗购自北京中杉金桥生物技术有限公司。
1.2 方法
1.2.1 分组 常规培养心肌细胞,取对数生长期者接种于96孔板(1×104个/孔)上,分别加入含1 mg/L LPS的培养基培养24 h[7],作为模型组;分别加入10、50、100 μg/mL 知母皂苷B-Ⅱ与含1 mg/L LPS的培养基培养24 h[8],作为低剂量组、中剂量组、高剂量组;将正常培养的细胞作为对照组。参照Lipofectamine2000转染试剂说明书,将si-NC、si-XLOC_032768转移至心肌细胞,加入100 μg/mL知母皂苷B-Ⅱ与1 mg/L LPS的培养基培养24 h,分别作为高剂量+si-NC组、高剂量+si-XLOC_032768组。
1.2.2 细胞凋亡率检测 分别取各组心肌细胞,加入预冷的PBS,4 ℃、3 000 r/min离心6 min,弃上清,向细胞沉淀中加入500 μL结合缓冲液重悬细胞,加入Annexin V-FITC、PI各5 μL,室温避光孵育10 min,流式细胞仪检测细胞凋亡率。
1.2.3 MDA水平及GSH-Px、SOD活性检测 取各组心肌细胞,采用硫代巴比妥酸法检测MDA水平,再加入0.25%胰蛋白酶消化,反复冻融法裂解细胞,采用黄嘌呤氧化酶法检测SOD活性,5,5′-二硫代双(2-硝基苯甲酸)比色法检测GSH-Px的活性,均严格按照试剂盒说明书进行操作。
1.2.4 NO、TNF-α、PGE2水平检测 取各组心肌细胞,反复冻融法裂解细胞,采用硝酸还原法检测NO水平,ELISA法检测TNF-α、PGE2水平,均严格按照试剂盒说明书进行操作。
1.2.5 细胞中XLOC_032768表达检测 采用实时荧光定量聚合酶链反应(RT-qPCR)法。取各组心肌细胞,通过Trizol法提取细胞中总RNA,Nanodrop2000c超微量分光光度计检测RNA浓度,XLOC_032768正向引物5′-CAAGGATTCT ACAGTCCAACAATTTC-3′,反向引物5′-CGATA CAAGTAATTTATTTCTAATTG-3′;GAPDH正向引物5′-AACGGATTTGGTCGTATTG-3′,反向引物5′-GGAAGATGGTGATGGGATT-3′,均由上海生工生物工程股份有限公司设计合成。RT-qPCR反应体系为cDNA 2 μL,Real-Time Master Mix 10 μL,正、反向引物各1 μL,RNase-Free ddH2O补足体系至20 μL;反应条件为95 ℃ 2 min,95 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s,循环40次。以GAPDH为内标,采用2-ΔΔCT法计算XLOC_032768相对表达。
1.2.6 Bcl-2、Bax、cleaved caspase-3蛋白表达检测 采用Western blot法,收集各组心肌细胞,加入RIPA裂解液提取细胞总蛋白,BCA法检测蛋白浓度,每孔加样40 μg,SDS-PAGE电泳分离蛋白,转移至PVDF膜,5%脱脂牛奶封闭2 h,加入一抗稀释液(1∶1 000),在4 ℃下孵育24 h,在TBST洗涤,加入二抗稀释液(1∶2 000),室温下孵育1 h,TBST洗涤,滴加ECL化学发光剂,暗室内曝光显影,通过Image J软件分析各条带灰度值。

2 结果
2.1 知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡的影响 与对照组比较,模型组凋亡率及Bax、cleaved caspase-3表达升高(P<0.05),Bcl-2表达降低(P<0.05);与模型组比较,各剂量组凋亡率及Bax、cleaved caspase-3表达降低(P<0.05),Bcl-2表达升高(P<0.05),见图1、表1。
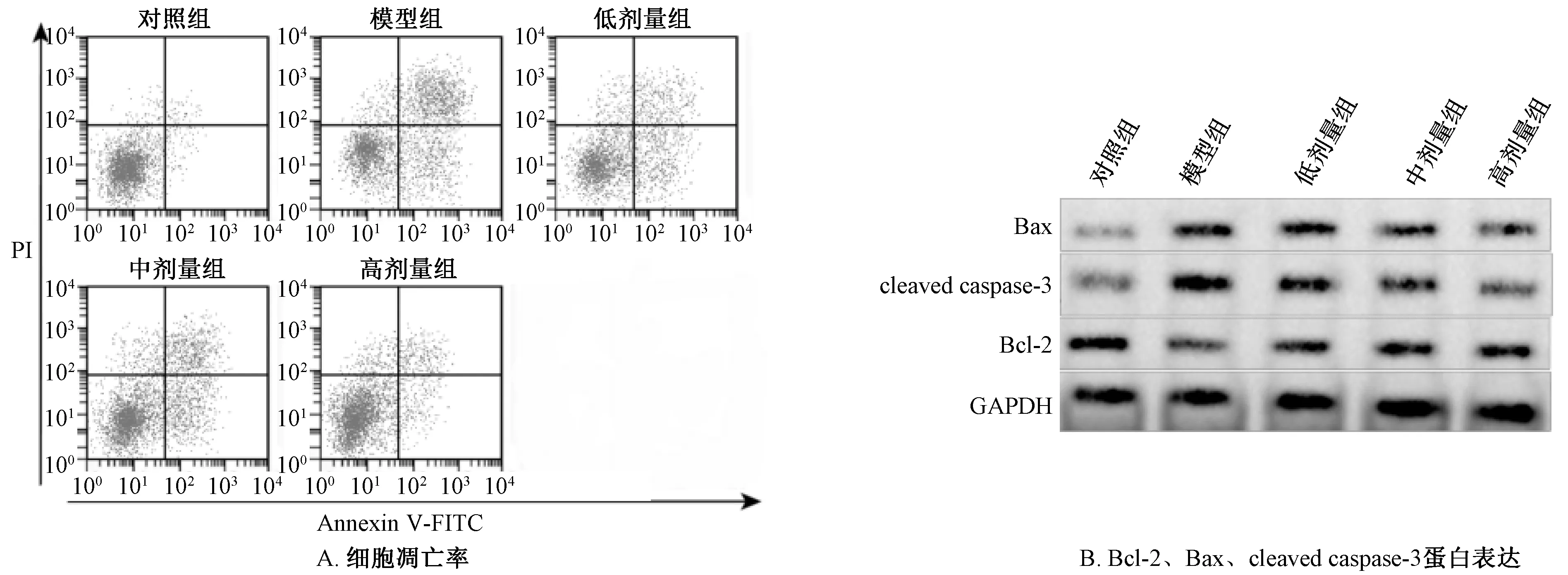
图1 知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡的影响(Ⅰ)

表1 知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡的影响
2.2 知母皂苷B-Ⅱ对LPS诱导心肌细胞氧化应激的影响 与对照组比较,模型组MDA水平升高(P<0.05),GSH-Px、SOD活性降低(P<0.05);与模型组比较,各剂量组MDA水平降低(P<0.05),GSH-Px、SOD活性升高(P<0.05),见表2。
表2 知母皂苷B-Ⅱ对LPS诱导心肌细胞氧化应激的影响
2.3 知母皂苷B-Ⅱ对LPS诱导心肌细胞炎症因子的影响 与对照组比较,模型组NO、TNF-α、PGE2水平升高(P<0.05);与模型组比较,低、中、高剂量组NO、TNF-α、PGE2水平降低(P<0.05),见表3。
表3 知母皂苷B-Ⅱ对LPS诱导心肌细胞炎症因子水平的影响
2.4 知母皂苷B-Ⅱ对LPS诱导心肌细胞中XLOC_032768表达的影响 与对照组比较,模型组XLOC_032768表达降低(P<0.05);与模型组比较,各剂量组XLOC_032768表达升高(P<0.05),见表4。
表4 知母皂苷B-Ⅱ对LPS诱导心肌细胞中XLOC_032768表达的影响
2.5 干扰XLOC_032768表达与知母皂苷B-Ⅱ联合作用对LPS诱导心肌细胞凋亡的影响 与高剂量+si-NC组比较,高剂量+si-XLOC_032768组凋亡率及Bax、cleaved caspase-3表达升高(P<0.05),Bcl-2表达降低(P<0.05),见图2、表5。
表5 干扰XLOC_032768表达与知母皂苷B-Ⅱ联合作用对LPS诱导心肌细胞凋亡的影响
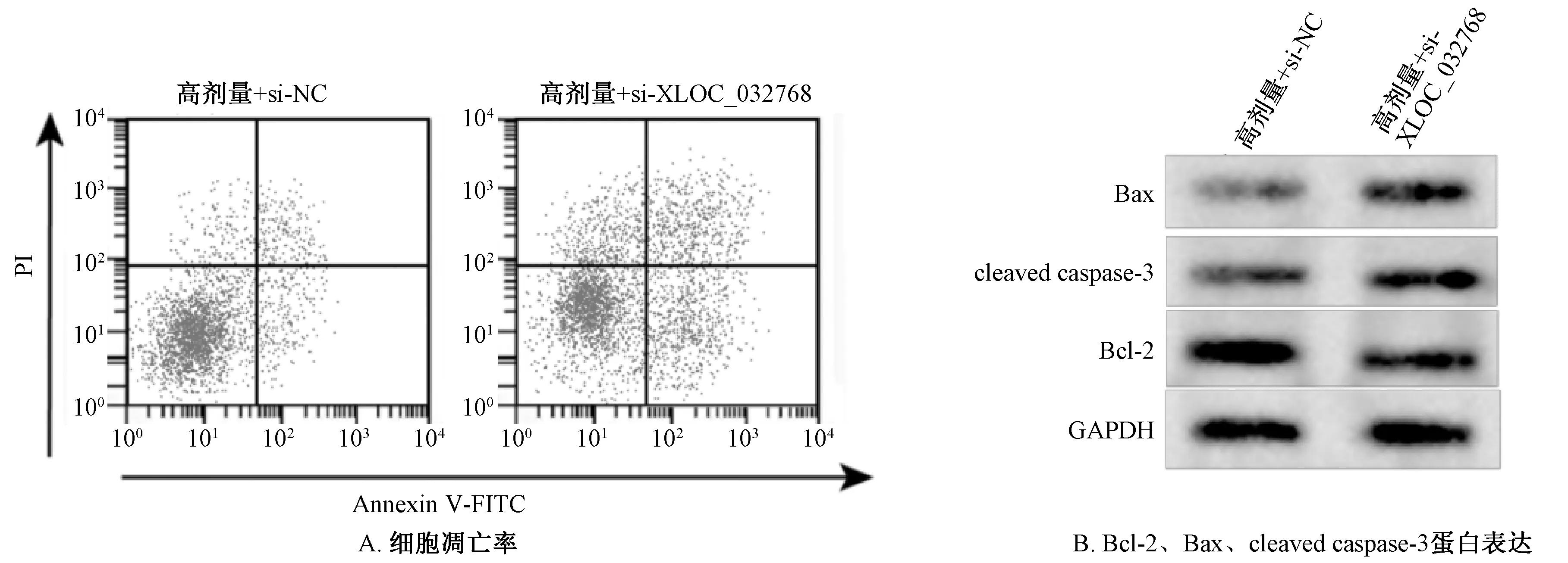
图2 知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡的影响(Ⅱ)
2.6 干扰XLOC_032768表达与知母皂苷B-Ⅱ联合作用对LPS诱导心肌细胞氧化应激及炎症因子的影响 与高剂量+si-NC组比较,高剂量+si-XLOC_032768组MDA、NO、TNF-α、PGE2水平升高(P<0.05),GSH-Px、SOD活性降低(P<0.05),见表6。
表6 干扰XLOC_032768表达与知母皂苷B-Ⅱ联合作用对LPS诱导心肌细胞氧化应激及炎症因子水平的影响
3 讨论
心肌细胞凋亡与氧化应激、炎症等密切相关,而心肌细胞凋亡可引发各种心血管疾病,因而抑制心肌细胞凋亡成为心血管疾病治疗的重要环节,既往研究显示中草药具有抗肿瘤、抗氧化应激等作用,并可抑制心肌细胞凋亡从而抑制心血管疾病的发展[9-11]。但知母皂苷B-Ⅱ在心血管疾病治疗中的作用机制尚未完全阐明。
知母皂苷B-Ⅱ可抑制氧-糖剥夺脊髓前角运动神经元瘤细胞凋亡从而对细胞发挥保护作用[12]。知母皂苷属于百合科植物,其是知母干燥根茎的主要成分,知母皂苷B是其主要活性成分,其具有抑制氧自由基生成及修复神经元损伤等作用[13]。知母皂苷B-Ⅱ可用于缺血性脑卒中的防治,此外,其可通过上调IL-18基因表达从而抑制肺癌细胞增殖及迁移[14]。本研究结果显示LPS诱导后心肌细胞凋亡率升高,而不同剂量的知母皂苷B-Ⅱ处理后凋亡率明显降低,且随着药物剂量的增加而明显降低。研究表明细胞接受到凋亡信号时,Bcl-2的表达降低,Bax的表达升高可促进线粒体释放细胞色素C而激活Caspase级联反应(cleaved caspase-3)从而促进细胞凋亡[15]。本研究结果显示LPS诱导心肌细胞中Bax、cleaved caspase-3的表达升高,Bcl-2的表达降低,不同剂量的知母皂苷B-Ⅱ处理后Bax、cleaved caspase-3的表达降低,Bcl-2的表达升高,证实知母皂苷B-Ⅱ可抑制LPS诱导心肌细胞凋亡。心肌细胞氧化损伤可诱导心肌损伤,氧自由基可被抗氧化酶降解从而促使机体维持动态平衡状态,GSH-Px、SOD属于抗氧化酶,其活性降低可促使细胞发生氧化损伤,氧自由基生成过量可促使细胞膜发生脂质过氧化从而破坏细胞膜完整性,MDA属于脂质过氧化的产物[16]。本研究结果显示LPS处理后心肌细胞中MDA水平升高,GSH-Px、SOD的活性降低,而不同剂量的知母皂苷B-Ⅱ处理后MDA水平降低,GSH-Px、SOD的活性升高,提示知母皂苷B-Ⅱ可抑制LPS诱导心肌细胞氧化应激,且呈剂量依赖性。炎症因子NO、TNF-α、PGE2水平升高可加重心肌细胞炎症损伤[17]。本研究结果显示LPS处理后心肌细胞中NO、TNF-α、PGE2水平升高,而不同剂量的知母皂苷B-Ⅱ处理后NO、TNF-α、PGE2水平明显降低,提示知母皂苷B-Ⅱ可抑制LPS诱导心肌细胞炎症反应,且呈剂量依赖性。
XLOC_032768表达下调可促进炎症因子的分泌从而加重急性肾损伤[18]。本研究结果显示LPS诱导心肌细胞中XLOC_032768的表达下调,而不同剂量的知母皂苷B-Ⅱ处理后XLOC_032768的表达升高,且随着剂量的增加而升高,提示知母皂苷B-Ⅱ可能通过上调XLOC_032768的表达从而减轻LPS诱导心肌细胞损伤。为验证知母皂苷B-Ⅱ是否可通过调控XLOC_032768的表达从而发挥作用。本研究将si-XLOC_032768与知母皂苷B-Ⅱ共同处理LPS诱导心肌细胞,结果显示凋亡率升高,Bax、cleaved caspase-3的表达上调,Bcl-2的表达下调,MDA、NO、TNF-α、PGE2水平升高,GSH-Px、SOD的活性降低,提示干扰XLOC_032768表达可降低知母皂苷B-Ⅱ对LPS诱导心肌细胞凋亡、炎症及氧化应激的抑制作用。
综上所述,知母皂苷B-Ⅱ可抑制LPS诱导心肌细胞凋亡、氧化应激及炎症反应,其作用机制可能与上调XLOC_032768的表达有关,XLOC_032768可能作为心血管疾病治疗的潜在靶点,还可为进一步揭示知母皂苷B-Ⅱ抗心血管疾病的分子机制奠定实验基础。但关于其具体作用机制仍需进一步探究。
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25
世界科学技术-中医药现代化(2021年7期)2021-11-04
Digital Chinese Medicine(2021年4期)2021-02-14
中成药(2018年9期)2018-10-09
中成药(2018年9期)2018-10-09
中成药(2018年5期)2018-06-06
中成药(2017年9期)2017-12-19
中成药(2017年9期)2017-12-19
中成药(2017年6期)2017-06-13
海南医学(2016年8期)2016-06-08