
苇状羊茅EST-SSR标记在草甸羊茅和多年生黑麦草的通用分析
2021-10-19 13:50王子玥刘凌云常智慧
草地学报 2021年9期
王子玥, 刘 曼, 刘凌云, 常智慧
(北京林业大学草业与草原学院, 北京 100083)
草甸羊茅(FestucapratensisHuds)是禾本科羊茅属的多年生草本植物,其植株高大、分蘖力强、叶量大,且耐酸、耐碱,营养丰富[1],是一种优良的牧草和草坪草,在我国主要分布于新疆伊犁。多年生黑麦草(LoliumperenneLam)是禾本科黑麦草属的多年生草本植物,原产于欧洲、亚洲和北非,是全球温带地区最广泛栽培的多年生冷季型草,主要用于草坪建植和牧草栽培[2]。草甸羊茅、多年生黑麦草这两种草与苇状羊茅(FestucaarundinaceaSchred)具有很近的亲缘关系,是优良的属间、种间杂交材料[3-6]。目前已有‘Johnstone’,‘Perny’,‘南农一号’三个羊茅-黑麦草品种通过国审[7]。
然而,种质资源评价体系不明确、选择周期长,阻碍了育种工作的顺利进行。与传统的形态学评价相比,DNA标记受环境影响较小,能够反映真实的遗传多样性。微卫星或简单序列重复(Simple sequence repeats,SSRs)具有高多态性、基因组广泛分布、共显性和可重复性等特点,常被用于遗传分析[8],主要包括基因组SSR和表达区SSR两种。其中表达序列标签(Experssed sequence tags,EST)SSR来自于相对保守的转录组数据,故在物种间有较好的通用性[9]。Ram等[10]采用了一套由甘蔗(Saccharumofficinarum)EST序列开发的60个SSR标记来研究它们在不同草类基因组的转移能力,发现上述标记在蔗茅属(Erianthus)、斑茅(Sclerostchya)、芒属(Miscanthus)转移率高达93.3%,86.3%和83.3%。张燕梅等[11]研究表明,剑麻(AaavesisalanaPerr)中的100对EST-SSR引物在龙舌兰属(Agave)、丝兰麻属(Yucca)、中美麻属(Furcraeavent)中分别有68,52和52对引物扩增出目标产物条带,扩增产物所占比例分别为68%,52%和52%。
黑麦草属(Lolium)和羊茅属(Festuca)杂交范围很宽,种、属间均可杂交,且存在较大变异性,中间类型多,种的界限不清晰,加大了育种研究的难度。而且,目前针对草甸羊茅的研究较少,开发标记不足。鉴于多年生黑麦草和草甸羊茅市场价值大,育种周期长,亟需开发出更有效的EST-SSR用于分子标记辅助选择育种。因此本研究选用40对EST-SSR引物对18份多年生黑麦草和18份草甸羊茅进行通用性分析,以期为两种种质鉴定、遗传多样性分析、分子标记辅助育种及种质资源合理利用提供更丰富的标记资源。
1 材料与方法
1.1 种质资源的收集
供试36份俄罗斯野生种质(表1)均由全国畜牧总站提供,其中包含了18份多年生黑麦草和18份草甸羊茅。此外,为了方便处理,本研究对36份种质进行了重新编号。
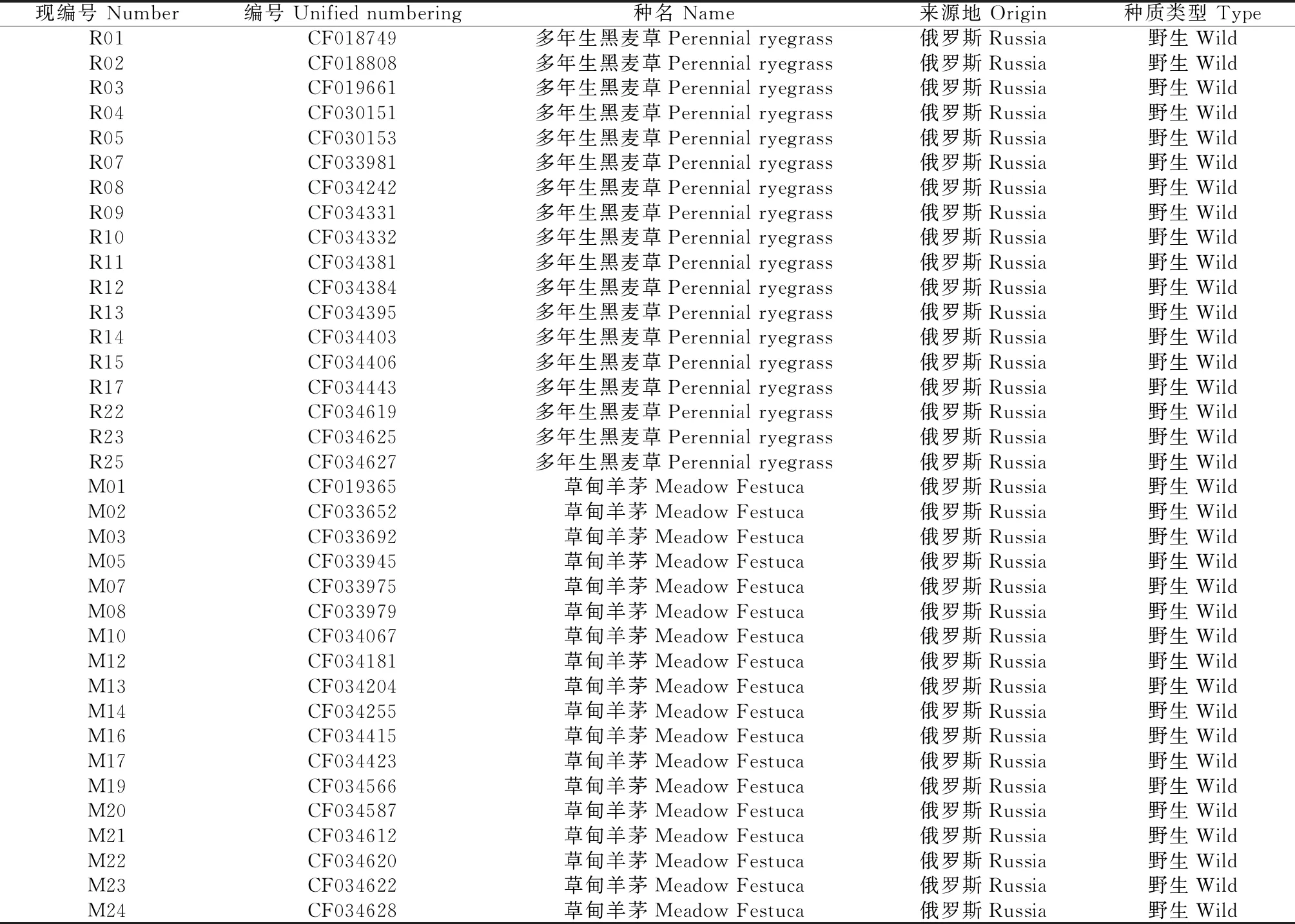
表1 36份种质信息表
1.2 基因组DNA的提取和EST-SSR标记
田间每份种质随机选择5株,采集新鲜叶片样品于—20℃保存[12]。混合样品经液氮研磨后使用Omega试剂盒提取DNA,其OD260/OD280均在1.7~2.0之间。将提取的DNA用2%琼脂糖凝胶电泳检测。最后,将试样浓度稀释至30 ng·μL-1,在三个离心管中各分装100 μL液体,—20℃保存,用于PCR反应。
EST-SSR引物由Saha等[13]设计的157条引物中随机挑选40条(表2),由北京睿博兴科生物技术有限公司合成。使用10 μL PCR反应体系,含有5 μL 2×Taq Master Mix(KT205,北京天根有限公司),0.2 μmol·L-1各引物,2 μL样品DNA和2.6 μL无菌水[14]。利用Bio-Rad T100 PCR仪进行PCR扩增反应,PCR扩增程序为:94℃预变性3 min;94℃变性50 s,59℃~64℃退火30 s,72℃延伸60 s,34个循环;72℃延伸10 min。将PCR产物在8.0%聚丙烯酰胺非变性凝胶上分离并通过银染法显现,使用凝胶成像分析系统拍照保存,用于后续分析。
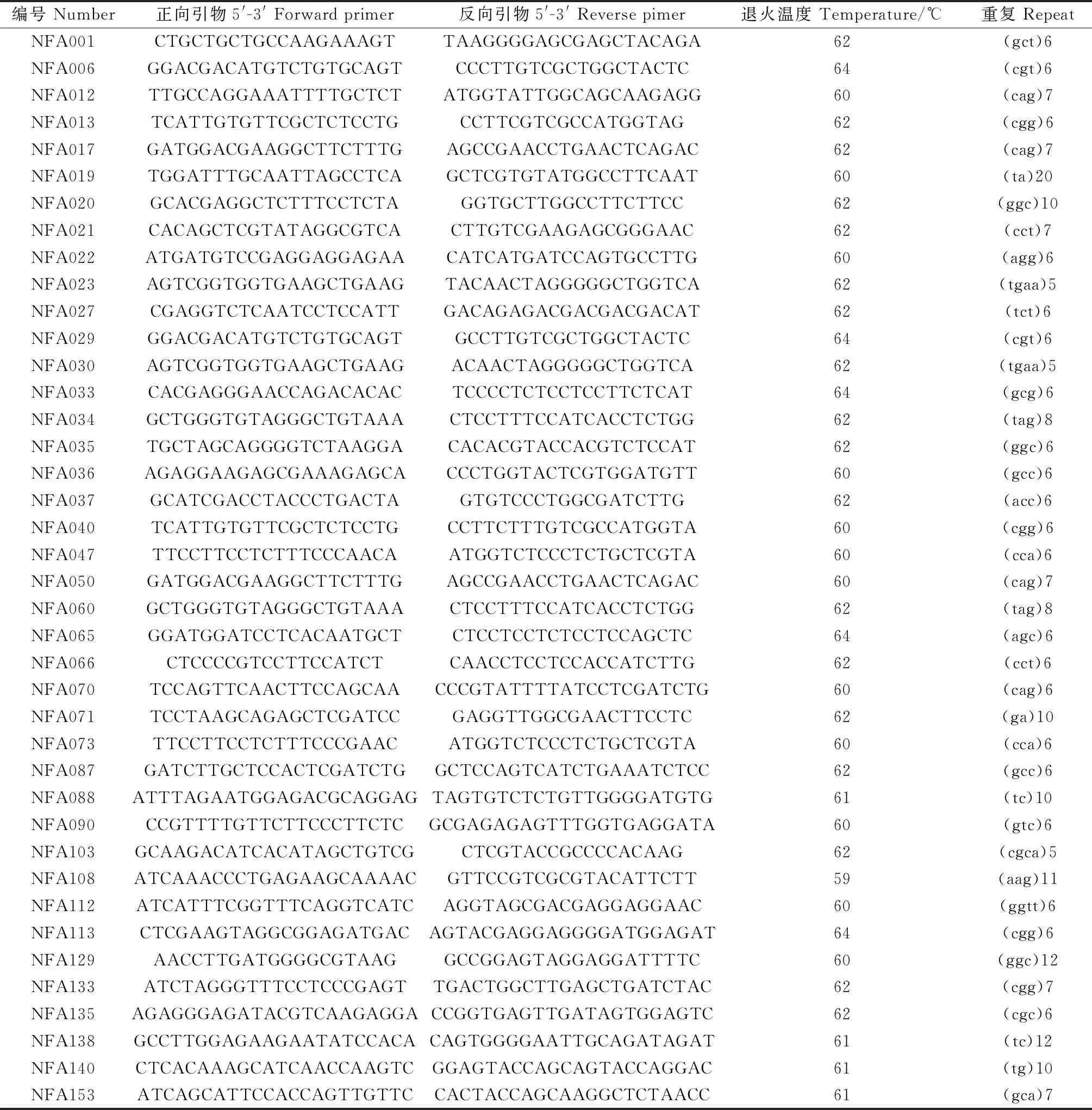
表2 40条苇状羊茅EST-SSR标记信息
1.3 数据统计与分析
PCR扩增产物按“有”或“无”条带进行统计,有条带的标记为“1”,无条带记为“0”[15]。遗传距离及聚类分析采用NTSYS软件分析;等位基因数(Observed number of alleles,Na)、有效等位基因数(The number of effective alleles,Ne)、Nei基因多样性指数(Nei’s gene diversity,H)、Shannon信息指数(The Shannon information index,I)采用POPGEN 32计算[16];引物的多态性信息含量(The polymorphic information content,PIC)值采用公式:PICi=2fi(1—fi)计算,其中PICi为标记i的多态信息量,fi为第i种等位基因占总基因数的比率,1—fi是缺少的基因频率[17]。
2 结果与分析
2.1 苇状羊茅EST-SSR标记在多年生黑麦草和草甸羊茅中的通用性
选取40 对EST-SSR引物对来自俄罗斯的18份野生多年生黑麦草和18份草甸羊茅进行PCR扩增。其中20对引物能在多年生黑麦草中扩增出清晰、有多态性位点的条带,22对引物在草甸羊茅中扩增出清晰、有多态性位点的条带。结果表明,苇状羊茅中开发的SSR引物在草甸羊茅和多年生黑麦草这两个近缘属中是可以通用的,且引物通用性分别为55%和50%(表3)。

表3 EST-SSR标记在两个草种中的通用性信息
2.2 EST-SSR扩增所得条带分析
2.2.1草甸羊茅扩增条带分析 结果表明22对引物在草甸羊茅中共扩增出212个条带,多态性条带总数为190条,多态带百分率为89.62%;平均每对引物扩增出9.6个条带,22对引物所扩增出的多态条带数在4(NFA047)~13(NFA036)之间,平均多态性条带数为8.6。

图1 18份草甸羊茅种质的扩增条带
2.2.2多年生黑麦草扩增条带分析 结果表明20对引物在18份多年生黑麦草中共扩增出203个条带,多态性条带总数为182条,多态性条带百分率89.67%,其百分率与草甸羊茅中的相似;平均每对引物扩增出10个条带,20对引物所扩增出的多态性条带数在4(NFA047)~16(NFA129)之间,平均多态性条带数为9.1。

图2 18份多年生黑麦草种质的扩增条带
2.3 UPGMA聚类分析
2.3.1草甸羊茅UPGMA聚类分析 聚类图显示,18份草甸羊茅的遗传距离在0.59~0.86之间,说明其遗传多样性处于较高水平。对结果进行相关性检验发现相关性系数为r=0.873,说明聚类结果可靠。在遗传距离为0.66时,M18被单独聚为一类,其余17份草种被聚为一类。在遗传距离0.688处18份草甸羊茅被聚为三类,M10,M18被分别单独聚为一类。另外M14和M15的遗传距离相同。

图3 18份草甸羊茅的UPGMA聚类图
2.3.2多年生黑麦草UPGMA聚类分析 聚类图显示,18份多年生黑麦草的遗传距离在0.65~0.84之间。对结果进行相关性检验发现,相关性系数为r=0.796,说明聚类结果可靠。在遗传距离为0.66时,R18被单独聚为一类,其他17份草种被聚为另一类。在遗传距离0.70处25份黑麦草被聚为三类,R7,R8,R9被聚为一类,R18被单独聚为一类。另外在遗传距离为0.84时R1和R3的遗传距离相同,该种质的同源性较高。总的来说草甸羊茅种质较多年生黑麦草遗传多样性高,且UPGMA聚类图准确性更高。

图4 18份多年生黑麦草的UPGMA聚类图
2.4 不同材料间的遗传参数
2.4.1草甸羊茅的遗传参数 18份参试草甸羊茅资源的观测等位基因数(Na)、有效等位基因数(Ne)、基因多样性指数(H)以及Shannon信息指数(I)平均值分别为1.845,1.337,0.212,0.336。PIC值用于比较各种标记的多态性潜力,草甸羊茅中PIC值范围在0.132~0.340之间,平均值为0.212。

表4 18份草甸羊茅的遗传参数
2.4.2多年生黑麦草的遗传参数 18份参试黑麦草资源的观测等位基因数(Na)、有效等位基因数(Ne)、基因多样性指数(H)以及Shannon信息指数(I)平均值分别为1.865,1.339,0.214,0.341。在多年生黑麦草中PIC值最大为0.316,平均值为0.214,该值略高于在草甸羊茅中的参数。

表5 18份多年生黑麦草的遗传参数
2.5 指纹图谱
扩增出的条带按“0”,“1”规则读数,并用不同颜色表示(其中黑色代表“1”,白色代表“0”),每个样品的引物扩增数据组合即为该样品的DNA指纹图谱。36份种质对比后发现,引物中NFA103,NFA140可以直接区别出草甸羊茅和黑麦草(图5)。其中引物NFA103在多年生黑麦草中同时存在Loc8,Loc9和Loc11三个位点,与草甸羊茅不同,引物NFA140在草甸羊茅中存在一条特异性条带LocD,可以直接通过这些等位基因区别供试草甸羊茅和多年生黑麦草。

图5 引物NFA103和FA140对于36份种质的指纹图谱
3 讨论
从不同物种获得的EST在SSR基序的侧翼序列中显示出基因组间的高度同源性[18]。因此,根据从一个作物物种获得的EST序列设计的标记,可用于没有或有限SSR信息的相关植物物种的遗传改良。黑麦草属和羊茅属曾同被划分为羊茅族[19],苇状羊茅、多年生黑麦草、草甸羊茅有着极近的亲缘关系。虽然Alm等[20]使用限制性片段长度多态性(Restriction fragment length polymorphisms,RFLPs)和黑麦草SSRs为草甸羊茅构建了遗传连锁图,但是依据草甸羊茅自身EST序列开发的EST-SSR标记罕见报道,草甸羊茅中的可用标记十分有限。从苇状羊茅EST-SSR标记中寻找适用于草甸羊茅和多年生黑麦草的引物既可以节约引物开发成本,又能提高引物的利用效率。本试验利用苇状羊茅引物的通用性成功的为草甸羊茅开发了22条新标记资源。40对引物在多年生黑麦草和草甸羊茅中的通用性比例为50%和55%,两个比例比林榕燕、迟恩惠、秦雪等人的研究低[21-23],其原因可能是不同植物基因组中编码序列保守程度以及突变频率不同。此外,在植物中,一般核心单元三核苷酸碱基重复类型的SSR通用性比例最高,其次是二核苷酸碱基重复类型[24]。本文所用引物两种重复类型占比为90%,而上述三人的供试引物均由二核苷酸和三核苷酸重复组成。
种质资源是开展育种工作的材料基础,对种质资源的鉴定、评价工作是推动资源进一步开发利用的前提[25]。在多年生黑麦草中,Faville等[26]对14 767个单基因分析,开发了310对有效SSR引物,构建两个亲本的遗传图谱。蒙宇等[27]用23对SSR引物对45份来自于24个不同国家的多年生黑麦草资源的遗传多样性进行分析,共扩增出78条清晰可识别条带,其中多态性条带有54条,占比69.2%,PIC值为0.082~0.499,平均值为0.324。本研究中多态性条带的占比较高,但是PIC均值略低,不过PIC均值与刘凌云等[14]在苔草中的研究数据相近。其原因可能是本研究中供试材料的遗传多样性不高,本试验中,供试材料18份多年生黑麦草的遗传距离在0.65~0.84之间,18份草甸羊茅的遗传距离在0.59~0.86之间,与陈志祥、伍越等[28-29]的研究相比,这36份俄罗斯野生种质遗传多样性处于中等水平。因此,就育种工作而言,一个地区的少量资源是不够的,还需要注重种质资源地理来源的跨度,增加育种亲本遗传的多样性。聚类结果显示草甸羊茅中的M10和M18,多年生黑麦草中的R18,与其他供试草种遗传距离最远,说明这3个材料遗传背景差异较大,具有较强的遗传潜力。而M14和M15,R1和R3具有极高的亲缘关系,说明这些资源具有相似的起源或者经过更多的基因交流[30]。
最后,本研究发现引物NFA103和引物NFA140分别存在3条和1条特异性条带,能够极其迅速的区别开两个草种。该引物可能对羊茅属间、羊茅属和黑麦草属间的分类具有重要意义,同时在实际应用中具有很大价值,但是受样本数限制,该引物的鉴定效果有待进一步确认。其他引物结果并不能显著区别两个植物种,种间鉴别的概率不高。而且,当涉及基因组某个DNA序列或一个染色体的变化时,SSR标记是不适用的[31]。目前羊茅-黑麦草品种已经开始广泛使用,单独使用分子标记进行遗传分析是片面的。因此,在后续羊茅属、黑麦草属草种的遗传分析中,还需要从解剖学特征、表观性状、质量性状等多角度进行探究。
4 结论
40对苇状羊茅EST-SSR引物在18份草甸羊茅和18份多年生黑麦草中的通用性比例分别为55%和50%;36份种质遗传多样性处于中等水平,但仍是优良的育种材料,其中草甸羊茅种质比多年生黑麦草遗传多样性高;引物NFA103和引物NFA140分别存在3条和1条特异性条带,能够极其迅速的区别两个草种,但是受样本数限制,该引物的鉴定效果有待进一步确认。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
奥秘(创新大赛)(2022年8期)2022-09-14
青海草业(2022年2期)2022-07-23
中国果业信息(2021年10期)2021-12-07
养殖与饲料(2020年6期)2020-02-18
中国种业(2019年8期)2019-08-26
活力(2019年21期)2019-04-01
山西地震(2019年1期)2019-03-20
河北林业科技(2016年5期)2016-11-08
新农业(2016年23期)2016-08-16
