
高寒草地凋落物覆盖对狼毒生长微环境及种苗定居的影响
2021-10-19 13:41夏建强李佳欣孙淑范
草地学报 2021年9期
夏建强, 张 勃, 李佳欣, 孙淑范, 汪 睿
(甘肃农业大学草业学院/草地生态系统教育部重点实验室, 甘肃 兰州 730070)
植物的繁殖成功是其种群得以再生补充和保持稳定的前提,是草地群落健康和生物多样性维持的关键[1]。种苗定居是植物通过种子繁殖体的有性再生过程,因此不仅影响植物种群的更新和扩散,而且影响种群的遗传多样性和进化潜力[2]。种苗定居过程包括种子萌发和种苗存活2个阶段,是植物生活史中对环境最为敏感的时期[3-4]。因此,研究植物种苗定居的生境选择对于了解植物种群动态及其适应性具有重要意义。
在高寒草地生态系统中,影响种苗定居成功的因素错综复杂,其中凋落物和种子埋深是2个最直接的因素[5-7]。凋落物是由草地生态系统中生物组分产生并归还到地面的有机物质总称,包括灌木的枯枝落叶、落皮和枯死的草本植物等[8]。凋落物作为植物种子掉落后首先接触到的物理环境,可通过物理屏障[8]、淋溶物化感作用[9]、阻隔光照[10-11]、改变土层温度以及增加土壤含水率[12-13]等环境效应,对植物种子萌发和种苗存活产生显著影响。另外,土壤埋深也是影响植物种子出苗和幼苗成功定居的主要因素[14-15]。不同植物的种子其萌发和出苗对土壤埋深的要求存在显著差异,适当的埋深可以提供种子萌发所需要的温度、湿度、光照以及厌氧微生物数量等生境条件[16],并能增强种苗根系的固着能力,而高于或低于一定阈值的土壤埋深均会对种子出苗产生不利影响[17-18]。
狼毒(Stellerachamaejasme)是广泛分布于亚洲温带草原的一种烈毒性草本植物,是草地发生退化的重要指示物种[19]。在天然草地中,狼毒完全依靠种子进行繁殖[20],因此研究狼毒种子萌发以及种苗定居的生境选择对于揭示该植物种群的入侵和扩散机制具有重要意义。近年来,国内外学者对狼毒种子的萌发特性[21-22]、休眠机理[22]、种群分布格局和散布机制[23]等进行了大量研究。王琳等[21]认为狼毒种子在自然状态下多数需经过休眠才能萌发,种皮对其萌发有一定的抑制作用,种子硬实性[20]是导致狼毒种子萌发率较低的主要原因;邢福等[22]认为狼毒整体萌发率较低,且种子萌发具有“偏高温型”的温度需求等。目前,有关狼毒种子萌发的研究多在实验室控制条件下完成,而在原生境中探究该物种种子萌发和种苗定居生境选择的研究尚未见报道。本试验以采自天祝高寒草地的狼毒种子为试验材料,在野外原生境开展其种苗定居试验,研究凋落物覆盖对狼毒生长土壤温度和含水量的影响、凋落物对狼毒种子的出苗和定居的影响以及狼毒种子在不同土壤埋深的出苗和定居的情况。
1 材料与方法
1.1 试验区概况
试验地设在青藏高原东缘天祝县甘肃农业大学高山草原试验站。该试验区海拔2 940 m左右,年均温在0.2℃~3.6℃之间,1月气温最低为-18.3℃;7月气温最高为12.7℃,>0℃积温为1 380℃;年降水量265~632 mm,年均蒸发量1 590.5~1 703.2 mm,降水分布不均,多集中在7,8,9三个月,无霜期3~4个月[24];属寒温带大陆性高原气候,夏季温凉干旱,冬季寒冷干燥,草地类型为高寒草甸。
1.2 狼毒种子和凋落物的收集与处理
1.2.1种子收集 2019年4月中旬,在狼毒自然居群,收集狼毒母株周围(半径30 cm以内)的浅层土壤(土表3~4 cm),剔除较大杂物后带回试验站;最后,将收集的土样通过网筛(孔径< 1 mm)筛掉细土后,挑选生长饱满、均匀一致的种子,装入尼龙网袋(40目),浅埋于(原生境)试验地备用。
1.2.2凋落物的收集 在试验区周围草地群落中,收集草地枯落物,适当剪碎后装入尼龙网袋,放置于试验地备用。
1.3 试验设计与播种方法
本试验采用双因素裂区设计。主区为覆盖方式,设2个水平:凋落物覆盖和裸露(对照组);裂区为不同埋深,设0.5 cm,2 cm和4 cm三个水平,共6个处理组合。狼毒种子种植于提前埋置的种植罐(直径10 cm、高20 cm的PVC管),每个种植罐播种5粒种子,每个处理24个种植罐(即24个重复),共120粒种子。播种前,首先将种植罐竖直埋入预先翻耕平整的试验区,罐口略高于土表;其次,将原生境土壤混合均匀后填入种植罐中。2019年5月3日播种,用直径3~4 mm的筷子头在种植罐中扎孔(5孔)至特定深度,每孔放入1粒种子,然后在种植孔填入细土。完成播种后,将凋落物均匀覆盖于相应处理的种植罐中。
1.4 土壤温度和含水率的测定
土壤温度用HH147AU型手持式热电偶温度计(OMEGA,美国)测量,土壤水分(%)用土壤墒情测定仪(杭州美农仪器有限公司,型号MN-TYS)测量。为测量不同土层的土壤温度以及含水率,预先在种植罐侧壁距地表0.5 cm,2 cm和4 cm处分别打孔,然后填埋。在种子播种后1周(5月初),选择适宜的天气,分别于9∶00—10∶00,14∶00—15∶00和19∶00—20∶00,完成对各处理土壤温度和含水量的测量。测量前,首先将种植罐打孔一侧刨开,然后迅速将温度和含水量测定仪的探头水平插入罐体进行测量,测量时用多个探头对不同处理同步测量,测量完成后将种植罐填埋复原。
凋落物覆盖的温度效应(℃)=覆盖处理的土壤温度值-对照(裸露)土壤的温度值;
凋落物覆盖的湿度效应(%)=覆盖处理的土壤水分含量-对照(裸露)土壤水分含量。
1.5 狼毒种子出苗、成苗和越冬率的统计
出苗率:在播种后30~45 d,统计各处理种植罐中的狼毒种苗,并计算种子出苗率(G);
式中,n表示种子出苗数,N表示播种的种子总数。
成苗率:在入冬前(9月中旬)统计各处理种植罐中存活的狼毒种苗,干枯或缺苗代表幼苗死亡,最后计算其成苗率;
成苗率(%)=成苗数/出苗数×100%
越冬率:于翌年(2020年)7月初,待狼毒种苗完全返青后统计各处理种植罐中存活的种苗(注意:仅统计二年生幼苗,排除播种当年未发芽而次年出土的幼苗)。
越冬率(%)=越冬存活株数/当年成苗数×100%
1.6 数据统计分析
在同一土层某一时段,凋落物覆盖与对照(裸露)土壤温度和含水量的差异性用成对数据的T检验(The paired)进行分析;凋落物覆盖对温度和含水量的影响效应在不同土层间的差异显著性用R软件的一般线性模型(LM)进行分析。凋落物覆盖和种子埋深对狼毒出苗(每个种植罐出苗数为0~5之间的计数数据)的处理效应用广义线性模型GLM(Poisson分布)进行分析。两处理对狼毒成苗率和越冬率的影响效应,将不同种植罐的数据合并后用卡方检验进行分析。所有的数据均使用R 3.6.1软件进行分析。
2 结果与分析
2.1 凋落物覆盖对不同土层土壤温度的影响效应
凋落物覆盖对土壤温度的影响随不同土层深度存在显著差异,其影响效应在各时段表现不同,结果如图1所示。凋落物覆盖处理下,0.5 cm土层土壤在9∶00—10∶00和14∶00—15∶00时段的温度显著低于对照;2 cm土层土壤温度的影响仅在9∶00—10∶00时具有显著性。在各时段,凋落物覆盖对0.5 cm土层土壤温度的影响均显著高于对2 cm和 4 cm(图1D,E和F)。其中,在9∶00—10∶00和14∶00—15∶00时段,凋落物覆盖处理后0.5 cm土层土壤温度的平均降幅分别为2.58℃和3.49℃,对4 cm土层温度的影响最小,土壤温度平均降幅为0.28℃和0.36℃。凋落物覆盖对各土层在19∶00—20∶00时段温度的影响总体较小;其中,对0.5 cm土层土壤有一定的增温效应,温度增幅为0.29℃,相反,对深层土壤(2 cm和4 cm)表现出不同程度的降温效应,2土层温度的平均降幅分别为0.2℃和0.46℃(图1F)。
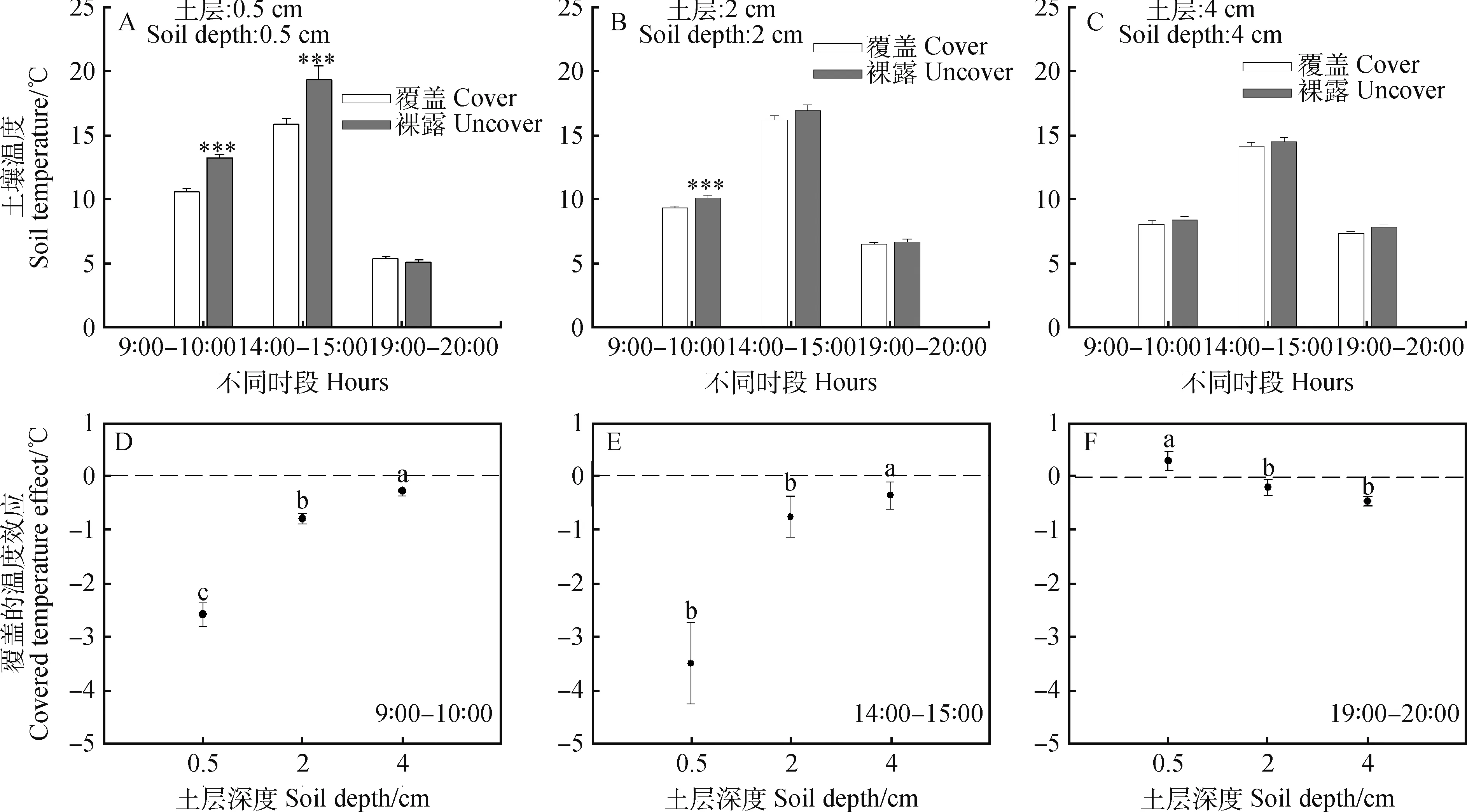
图1 凋落物覆盖对不同土层土壤温度的影响
2.2 凋落物覆盖对不同土层土壤含水量的影响效应
凋落物覆盖对土壤含水量的影响随不同土层深度和不同时段存在显著差异,结果如图2所示。凋落物覆盖后,0.5 cm和2 cm土层的土壤含水量在9∶00—10∶00和14∶00—15∶00两个时段均显著高于对照;在19∶00—20∶00时段,覆盖处理下2 cm和4 cm土层的土壤含水量显著高于对照(图2A,2B和2C)。在9∶00—10∶00和14∶00—15∶00两个时段,凋落物覆盖处理下0.5 cm土层土壤含水量的平均增幅分别为8.46%和8.42%,4 cm土层土壤含水量的平均增幅最小,分别为4.84%和3.94%,均显著低于0.5 cm土层土壤(P< 0.05)。在19∶00—20∶00时段,凋落物覆盖处理对各土层含水量的影响差异不显著(图2F)。
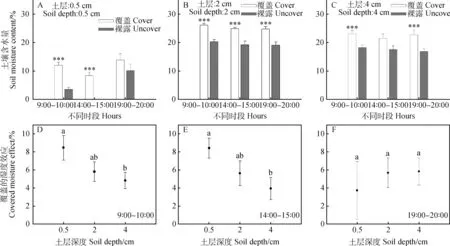
图2 凋落物覆盖对不同土层土壤含水量的影响
2.3 凋落物对狼毒种子出苗和种苗存活(成苗和越冬)的影响
凋落物覆盖对狼毒种子的出苗和种苗存活(即成苗率和越冬率)均有显著的影响,结果如图3所示。狼毒种子在裸露土壤(对照)的平均出苗率为23.61%,在凋落物覆盖土壤(处理组),狼毒种子的出苗率显著低于对照(P<0.001,图3A),其平均出苗率为11.67%。出苗后,对照(裸露)组狼毒种苗的成苗率为94.12%,覆盖处理组的成苗率为83.33%,二者存在显著差异(P<0.05,图3B)。凋落物覆盖处理组与对照(裸露)组的越冬存活率差异不显著(图3C)。

图3 覆盖处理对狼毒种子出苗和种苗存活的影响
2.4 种子埋藏深度对狼毒种子出苗和种苗存活(成苗和越冬)影响
土壤埋藏深度对狼毒种子的出苗和种苗存活(即成苗率和越冬率)有显著影响,结果如图4所示。总体上,狼毒种子的出苗率随土层深度增加而显著下降。种子埋植于0.5 cm土层时,出苗率最高,为37.08%;当埋植于2 cm土层时,种子出苗率显著降低至13.75%(P<0.001);当埋深至4 cm土层时,狼毒种子出苗率仅为2.08%,显著低于0.5 cm土层和2 cm土层种子的出苗率(P<0.001)。种子在土壤的定植深度对狼毒种苗成苗率没有显著的影响(图4B)。土壤埋深对狼毒种苗越冬存活率有显著影响,定植于0.5 cm土层土壤的种苗,其越冬率为97.47%,显著高于深层(2 cm和4 cm)定植狼毒种苗的越冬率(P<0.01,图4C)。

图4 不同埋深对狼毒种子出苗和种苗存活的影响
3 讨论
3.1 凋落物对狼毒种子出苗和种苗存活的影响
种子萌发和幼苗定居是植物生活史中最为脆弱的两个阶段[3,9]。研究表明,在高寒草地群落中,凋落物的存在能显著改变群落的微生境条件,是影响植物种子萌发和幼苗定居的重要因素[2]。凋落物覆盖能通过阻隔太阳辐射,减缓其下层土壤与外界空气的热交换,因此总体上能降低土壤温度。但是,土壤平均温度的降低并非是影响草地植物种子萌发的关键因素[25-26]。潘开文等[27]认为,凋落物主要通过改变土壤温度的日温差对植物种子的萌发产生影响。本研究发现,凋落物覆盖处理对表层土壤温度的影响较为明显,在白天能显著降低土壤温度,相反在晚间(19∶00以后)能起到一定的保温作用,从而导致凋落物覆盖土壤的温度变幅显著低于裸露土壤。相应地,凋落物覆盖处理下,狼毒种子的出苗率也显著低于对照(裸露)土壤。据此推测,凋落物覆盖后狼毒种子较低的萌发率可能与土壤温度日振幅的降低有关,这与邢福等[22]发现较大的温差更有利于打破狼毒种子休眠从而提高其萌发率的结果相类似。
另外,适宜的水分是植物种子萌发和幼苗生长的基础。大量的文献表明,凋落物能通过吸持和拦截水量[28]、减少地表蒸发[29-30]和改善土壤结构[27]等途径增加土壤保水能力,提高土壤含水量。本研究中,凋落物覆盖处理同样能显著提高土壤含水量,但是并没有提高狼毒种子的出苗率,相反表现出明显的抑制作用。有研究表明,种皮限制是抑制狼毒种子萌发的主要因素之一[22]。凋落物覆盖虽然能够增加土壤含水量,但同时也降低了土壤含水量的日变化,这样会使狼毒种子保持在含水量相对稳定的微环境中,不利于种皮的破裂,从而抑制了种子的萌芽。最后,凋落物除了能通过改变土壤温度和含水量影响种子的出苗,还可通过直接的物理屏障作用[8]或通过影响幼苗的光合作用等影响狼毒种苗的存活和定居成功[27]。综合分析表明,尽管凋落物覆盖在高寒草地中可以营造和维持相对稳定的植物生长微生境,但是从狼毒种子萌发所需条件的角度出发,该环境并非是狼毒种苗定居的有利生态条件。因此,在草地管理中,维持草地盖度,并保持一定的凋落物覆盖量可能遏制狼毒种苗的定居和种群扩展。
3.2 土壤埋藏深度对狼毒种子出苗和种苗存活的影响
在草地生态系统中,植物种子会通过多种途径被埋于不同深度的土壤中,例如,啮齿动物储存和草食动物踩踏等[31]。埋藏在土壤中的种子通常有4种结果:一部分种子正常萌发,一部分因不适宜的土壤环境而死亡,一部分进入休眠储藏于土壤种子库,还有一部分种子尽管会萌发,但是因幼苗不能够伸长到地面而死亡[32]。本研究结果显示,当狼毒种子埋植于土壤表面0.5 cm以内时,出苗率最高;当埋深超过2 cm时,其出苗率显著降低;当埋深达4 cm时,出苗率降低了94.38%。这些结果表明,在高寒草地原生境下,狼毒种子埋藏于土壤表层时更有利于其种苗的萌发和定居。依据前人的研究,植物种子埋藏于深层土壤,其出苗率降低的原因主要包括3个方面。首先,当植物种子在土壤中超过适宜的埋藏深度时,因顶土压力增大,种子贮藏的能量不能为种子萌发和出苗提供保障,因此影响其出苗率[33-34]。其次,随着土壤深度的增加,土壤含氧量显著降低,从而能使种子在低氧的环境下进入强迫休眠状态或吸涨后死亡[35]。最后,深层土壤的温度振幅通常低于表层,其温度变幅的降低将不利于需要昼夜变温才能打破休眠的植物种子萌发。本研究结果在一定程度上支持了该假说。Grime等[36]研究表明,土壤表面的大多数种子需要≥10℃的温度振幅就能很好地萌发,而埋藏于土壤深层的种子则需要≥15℃的振幅才能萌发。最后,本研究结果也显示,随土层深度的增加,不仅能抑制狼毒种子的出苗,同时能显著降低种苗在翌年的存活率,即种苗的越冬率。一方面,种植太深致使种苗出土延迟,从而缩短了其生长发育期,导致根系的营养储存不足,从而影响其越冬率;另一方面,种植太深可能不利于狼毒种苗地下芽的发育形成或返青出土,从而导致其越冬率的下降[37],这些假说需要在今后的研究中进一步证实。综合分析表明,在高寒草地群落中,狼毒主要依靠表层土壤中的种子完成其种群更新,被埋藏于深层土壤中的狼毒种子可能会因土层阻隔、含氧量骤降而强迫休眠或因缺少适宜的变温条件,导致其出苗和定居的失败。
4 结论
本研究结果表明,凋落物覆盖总体上能降低土壤温度、增加土壤含水量,且能显著降低表层土壤温度和含水量的日变幅;与裸露土壤相比,凋落物覆盖能显著降低狼毒种子的出苗率和幼苗定居;狼毒种子在土壤的埋植深度显著影响其出苗率和种苗定居,埋植于表层土壤(0.5 cm)的种子相对于埋植在深层土壤(2 cm和4 cm)的种子具有更高的出苗率和越冬存活率。综合分析表明,高寒草地凋落物的存在或土壤种子深埋均不利于狼毒的种苗定居,草地退化后导致的草地裸露或促使土壤种子浅表化的扰动均可促进狼毒种苗定居和种群发展,是导致退化草地狼毒种群扩散的主要生态因素。因此,在高寒草地管理中,合理控制扰动强度(如放牧等),维持草地充分的盖度,并保持一定的凋落物覆盖量,可有效遏制狼毒的入侵扩散和草地退化。
猜你喜欢
辽宁农业科学(2021年5期)2021-11-03
农民致富之友(2020年14期)2020-05-25
农民致富之友(2019年34期)2019-12-20
水土保持研究(2019年6期)2019-10-19
草业学报(2019年3期)2019-03-23
中成药(2018年7期)2018-08-04
新农业(2018年6期)2018-04-18
辽宁林业科技(2017年4期)2017-06-22
浙江大学学报(工学版)(2016年2期)2016-06-05
现代农业(2016年5期)2016-02-28
