
盐碱胁迫对盐地碱蓬生长、有机酸等溶质积累及其生理功能的影响
2021-10-19 13:41张洪嘉库都斯阿布都沙拉木耿淑娟薛傲然赵翰韦菅柏涵
草地学报 2021年9期
麻 莹, 张洪嘉, 库都斯·阿布都沙拉木, 耿淑娟, 薛傲然, 赵翰韦, 菅柏涵
(1.吉林大学植物科学学院, 吉林 长春 130062; 2.六盘水市第三中学, 贵州 六盘水 553000)
土壤盐碱化已成为限制我国畜牧业生产的最大障碍因素[1]。土壤盐碱化既有中性盐又有碱性盐,而植物所面对的胁迫既有盐胁迫又有碱胁迫,盐胁迫和碱胁迫是2种不同性质的胁迫;二者的胁迫作用机理及植物对其生理响应也不同[1]。盐胁迫主要有渗透胁迫和离子胁迫;而碱胁迫除上述胁迫外,还涉及高pH胁迫[1]。高pH直接干扰Na+、K+选择性吸收,抑制阴离子吸收,打破植物体内已经形成的离子稳衡态、电荷平衡、pH稳定[2]。可见,碱胁迫对植物的破坏力更为严重[2]。
一直以来,植物抗盐生理多集中在以中性盐NaCl为主的研究[3-6]。近年来,虽然有一些有关植物对盐碱胁迫的生理响应[1-2]、种子萌发[7],抗碱性与有机酸代谢[1],碱胁迫下的基因表达[8]等研究报道,但所涉及植物盐碱胁迫的研究都集中于对植株整体或是植物器官的研究,而以碱蓬根或叶细胞液为切入点,探讨其溶质响应盐、碱胁迫的特点并对其溶质进行功能分析的研究却鲜见报道。许多东北碱化草原土壤pH值已高达10以上,虽然碱化土壤不利于植物生长,但是有一些抗碱性极强的植物能够生存[9],如盐地碱蓬(Suaedasalsa(L.) Pall.)就是珍贵的天然高抗碱牧草,它是研究植物抗盐碱机制的理想材料。
本文分别用中性盐(NaCl,Na2SO4)和碱性盐(NaHCO3,Na2CO3)模拟盐胁迫或碱胁迫条件,并以此对碱蓬幼苗进行胁迫处理。通过比较根系、叶片细胞液的有机酸等溶质应答盐碱胁迫的变化,进一步探讨体内溶质的积累及其功能特点,为探明碱蓬适应盐碱生境特殊的生理机制提供理论依据。
1 材料与方法
1.1 材料培养
在中国吉林省西部天然草原采集碱蓬(S.salsa)种子,采用砂培法培养[1],具体方法为:种子播种于洗净细砂的塑料花盆中(高度14 cm,直径17 cm),每盆10 孔穴,每孔播种4粒种子,出苗后间苗2次,最终每孔保留1株幼苗,即每盆10 株幼苗。幼苗每天晚5∶00—6∶00用 500 ml营养液(Hoagland)透灌。整个试验在室外进行,处理期间人工遮雨。
1.2 试验设计
将2种中性盐NaCl和Na2SO4混合(1∶1的摩尔比)作为盐胁迫(A组),将2种碱性盐NaHCO3和Na2CO3混合(同上)为碱胁迫(B组)。A,B组处理液均用Hoagland作为溶剂配制,盐浓度均为100 mmol·L-1;以Hoagland溶液作为对照组(CK)。盐胁迫(A)、碱胁迫(B)和对照组(CK)溶液的pH值分别是6.55,9.90,6.67;其中碱胁迫的pH值与典型的碱蓬生境的pH值相近[9]。
苗龄6周后,选取12 盆长势一致的幼苗,随机分成3组,每组4盆为4次生物学重复,其中1组为对照组,其余2组为盐、碱胁迫组。盐胁迫或碱胁迫每天分别用100 mmol·L-1盐或碱处理液浇灌1次(晚5∶00—6∶00);对照组只浇Hoagland营养液。胁迫处理7 d。
1.3 取样、植物细胞液的收集
处理7 d后,随机选取5株植株在子叶节处分成地上部(Shoot)和根部(Root)2部分,依次用自来水、蒸馏水冲洗干净,吸干表面水分。分别称取茎叶和根鲜重(FW),然后将其真空冷冻干燥,冻干,分别称取干重(DW),计算植株含水量(WC)及其生物量。另外5株植株分别用蒸馏水清洗1次,去离子水清洗2次,用滤纸将其表面多余水分吸干。将植株的全部叶片和根部剪下,采用离心法[10](2 500 g离心15 min,4℃)去除根、叶质外体汁液后再去除其老根和叶的主脉,将植物材料分别装入封口袋,排净空气后在-20℃冷冻24 h,在干燥器中解冻,分别用注射器收集根和叶的细胞汁液[11],其汁液用于各种溶质含量的测定。
1.4 生长、生理指标测定
生物量为每株植物干重(g)。含水量(WC)= (鲜重-干重)/鲜重×100%[12]。用离子色谱法测定无机阴离子、草酸(DX-300离子色谱系统,CDM-II电导检测器,AS4A-SC离子交换色谱柱,流动相为Na2CO3/NaHCO3=1.7/1.8 mmol·L-1)和其他有机酸含量(DX-300离子色谱系统,AMMS-ICE II消声器,ICE-AS6离子排斥柱,流动相为0.4 mmol·L-1全氟丁酸)[13]。以上色谱试验均在室温25℃进行。
K+和Na+含量用火焰光度计测定[12]。可溶性糖和脯氨酸含量分别采用蒽酮显色法和茚三酮比色法测定[1]。之前压榨挤出的根、叶的汁液(细胞液)用数字酸度计测定pH值(图5),根据其pH值用Geochem软件计算各离子的游离电荷数量(meq·L-1组织水),分别计算各种游离态离子所贡献的正电荷或负电荷数量占总正或负电荷的百分比,即表示各种离子对离子平衡的贡献率。
阳(阴)离子的电荷贡献率计算公式:
阳(阴)离子的电荷贡献率(%)=游离态阳(阴)离子所贡献的正(负)电荷/各种游离阳(阴)离子的正(负)电荷的总和×100%。
溶质渗透势计算公式:
ΨS=-nRT/V,其中n表示溶质摩尔浓度,R和T为常数。因此,本试验用某一溶质的摩尔浓度(mmol·L-1组织水)占各种溶质摩尔浓度的总和的百分比,表示此溶质对渗透调节贡献率[13]。
1.5 数据处理及统计学分析
数据整理和作图采用Excel 2010;SPSS20.0软件对试验数据进行单因素方差分析,检验水平为0.05;多重比较采用LSD法,数据用平均值±标准误(±SE)表示。
2 结果与分析
2.1 盐碱胁迫对碱蓬幼苗生长的影响
由图1A可知,盐胁迫下碱蓬生物量有所降低,但未达到显著水平,而碱胁迫下其生物量显著下降(P<0.05)。由图1B所示,对照(CK)碱蓬植株含水量达到90%以上,盐胁迫对碱蓬含水量影响不显著,在碱胁迫下其含水量有所降低,但仍保持较高的含水量(88.26%)。
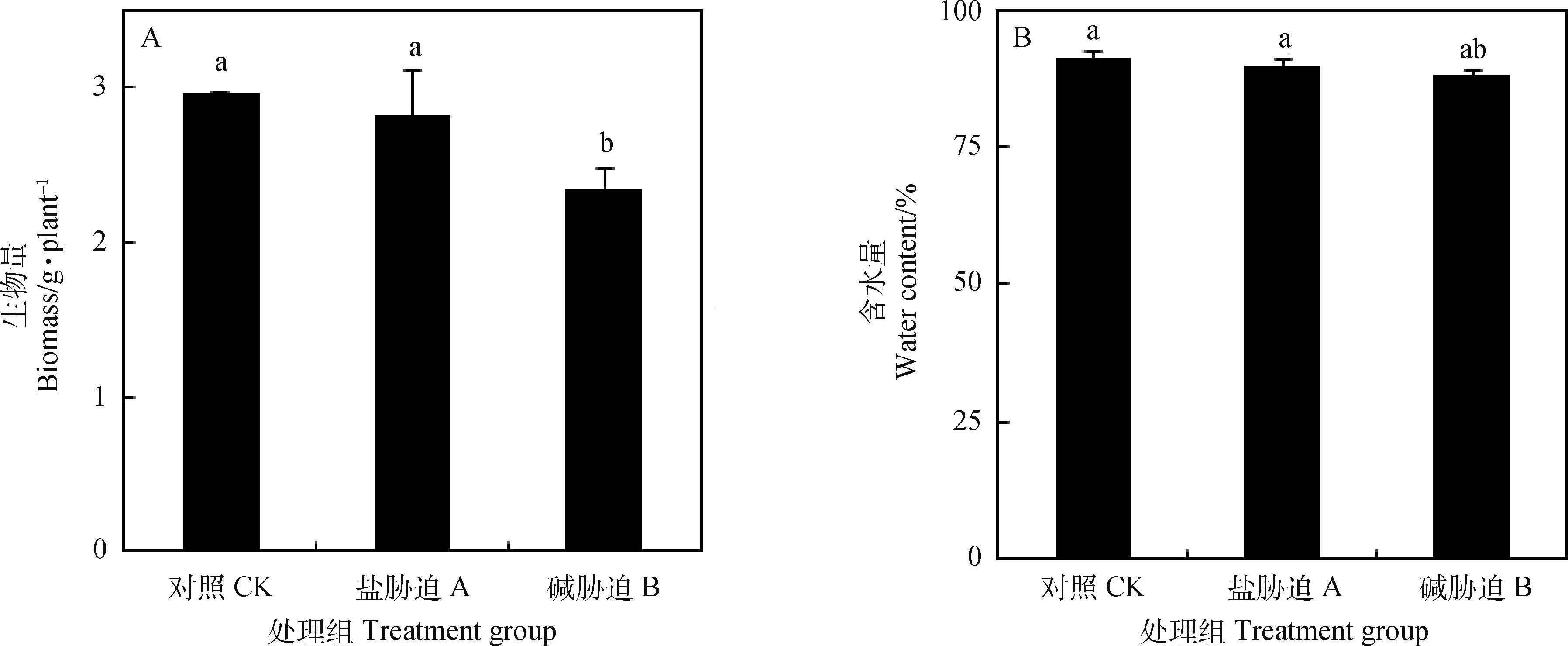
图1 盐、碱胁迫对碱蓬生物量和含水量的影响
2.2 碱蓬根、叶的阳离子对盐碱胁迫的响应
盐、碱胁迫均会引起叶片、根部细胞液Na+含量的显著增加(P<0.05),其中盐胁迫下叶片Na+含量极显著增加(P<0.01)(图2A)。
根部细胞液的K+含量在碱胁迫下略有下降;在盐胁迫下,叶片细胞液K+含量显著低于对照和碱胁迫组(P<0.01)(图2B)。
由图2C可见,与对照相比,根部细胞液的Na+/K+在盐、碱胁迫下比值显著升高(P<0.05),而盐、碱胁迫对其作用差别不大。叶片细胞液Na+/K+在盐、碱胁迫下也显著升高(P<0.05),而碱胁迫下其比值低于盐胁迫。
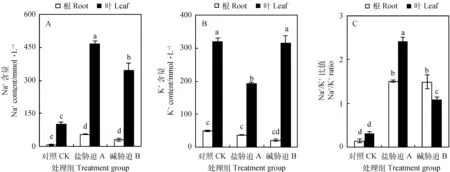
图2 盐、碱胁迫对碱蓬根和叶片中阳离子的影响
2.3 碱蓬根、叶的阴离子对盐碱胁迫的响应
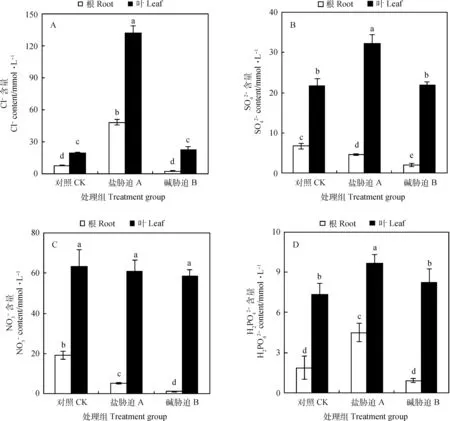
图3 盐、碱胁迫对碱蓬根和叶片中阴离子的影响
2.4 碱蓬根、叶的有机溶质对盐碱胁迫的响应
由图4所示,叶片脯氨酸、可溶性糖、有机酸等有机溶质明显高于根部其含量,而且根部的有机溶质含量在盐、碱胁迫作用下,其含量几乎不积累。在叶片中脯氨酸、可溶性糖、有机酸等有机溶质含量在盐、碱胁迫下均显著增加(P<0.01)。但从叶片的最大积累量上看,脯氨酸积累量最少,有机酸最多(其最大积累量是脯氨酸的157.06倍)。
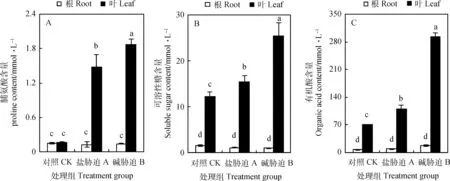
图4 盐、碱胁迫对碱蓬根和叶片中有机溶质的影响
2.5 盐、碱胁迫对根和叶片组织液pH的影响
由图5可知,盐碱胁迫下碱蓬根、叶片的细胞汁液pH都在正常生理pH范围内(pH<7),盐碱胁迫下碱蓬根部的细胞汁液pH的变化显著(P<0.05),而其叶片细胞汁液pH差异并不显著。
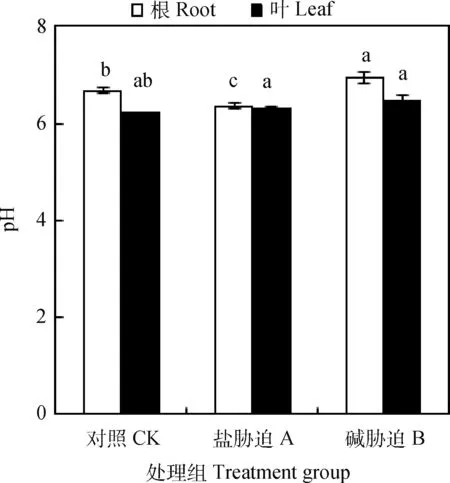
图5 盐、碱胁迫下碱蓬根和叶片的细胞汁液pH值
2.6 各种溶质对渗透调节的贡献
由表1可知,对照K+在根和叶片的渗透调节占据主导地位。盐、碱胁迫后Na+对渗透调节的贡献率有所升高,但是K+的贡献依然较高(仅次于Na+);与盐胁迫相比,碱胁迫下叶片的K+的贡献率提高,而Na+的比例下降。根和叶片的有机酸对总有机溶质的贡献占据绝对优势(84.76%~97.25%),盐、碱胁迫,特别是碱胁迫下有机酸的贡献率达到最大(表1),而脯氨酸对渗透调节的贡献甚微(不足0.20%)。
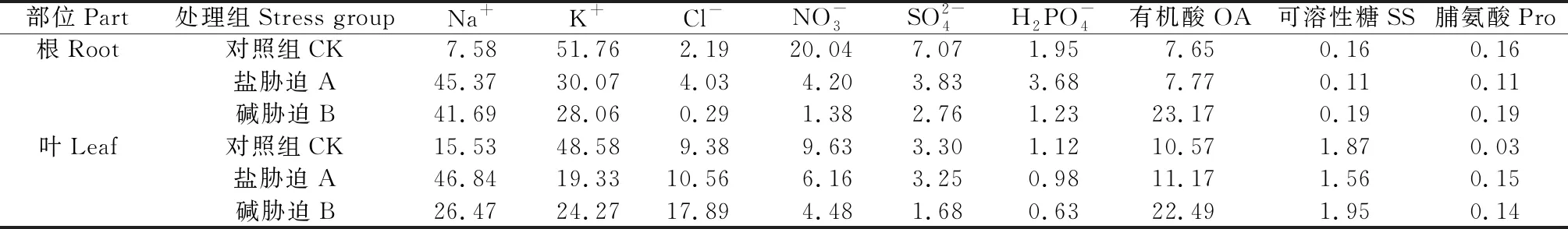
表1 盐碱胁迫下碱蓬根、叶细胞液中各溶质对渗透调节贡献率
2.7 阳(阴)离子对总正(负)电荷的贡献
对照植株根和叶片中K+对正电荷的贡献率最大;盐碱胁迫后,K+对正电荷的贡献率降低,Na+达到最大(表2),碱蓬根和叶片在碱胁迫下K+贡献率均高于盐胁迫,特别是叶片在碱胁迫下K+贡献率(47.83%)高于盐胁迫下(29.22%)。
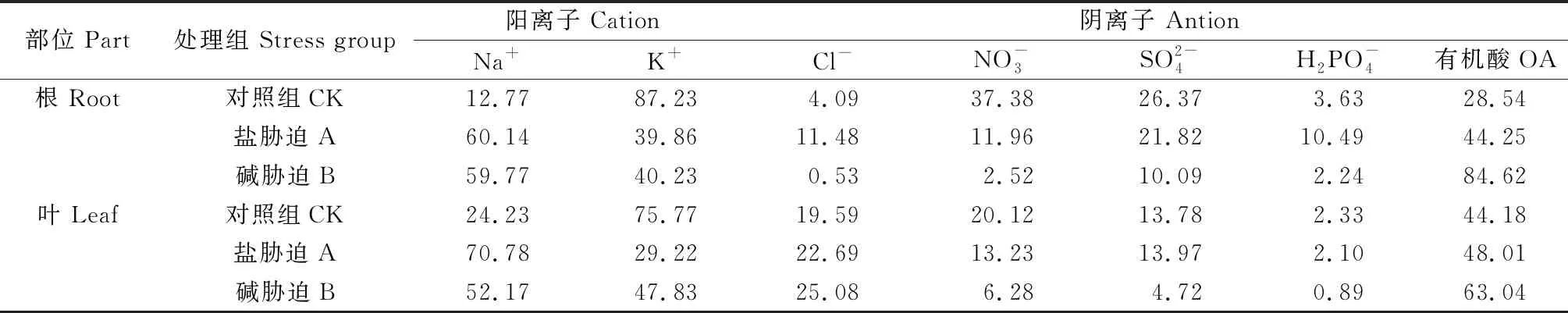
表2 盐、碱胁迫下碱蓬根、叶中各种游离态离子对总正(负)电荷的贡献率
3 讨论与结论
3.1 盐碱胁迫对碱蓬生长的影响
生物量降低是植物生长受到抑制的直接表现。本研究中,碱蓬只在碱胁迫下其生物量才显著下降。可见,盐胁迫对碱蓬生长影响不大,而只有高pH的碱胁迫才抑制其生长。说明碱蓬具有较强的抗盐能力,同时也证实碱胁迫对植物具有更大的破坏性[14-15]。
植物通过降低含水量来降低细胞渗透势是既迅速又经济的方式[14]。盐胁迫对碱蓬含水量影响不大,碱胁迫下其含水量下降,但仍保持较高水平(88.26%)。碱蓬作为稀盐植物,其植株高度的肉质化可以最大限度地稀释液泡中有害离子(Na+,Cl-)的浓度[16]。这也许是决定碱蓬抗盐碱的关键生理特性之一。
3.2 盐碱胁迫对碱蓬体内离子积累的影响
盐碱胁迫下,Na+毒害是影响植物生长的主要原因之一。Na+区域化到液泡是减少细胞质中过多Na+的有效方法之一[17]。Na+区域化主要依靠液泡膜Na+/H+逆向蛋白(Na+/H+antiporter protein,NHX)实现[18]。本试验盐、碱胁迫引起碱蓬叶片和根部细胞液Na+激增,特别是盐胁迫的叶片Na+含量达到最大值(其含量是对照的4.55倍),而此时叶片的高浓度Na+并没有影响碱蓬植株的生长。可以推测碱蓬细胞内可能存在着高效的NHX将Na+区域化到液泡,而其具体的转运机制还需进一步探究。此外,碱蓬作为真盐生植物,其肉质化的叶片具有稀盐作用。根从盐碱土壤吸收的Na+等离子输送至叶片,并将离子区域化于肉质叶片的大液泡中,其较高含水量具有稀释离子浓度的作用,以此保护胞质内酶类等活性物质免受毒害,保证了植株的正常生理代谢。
植物体内维持较低Na+/K+对其抵抗胁迫至关重要[19]。碱胁迫下叶片依然保持低Na+高K+,其Na+/K+比值低于盐胁迫,而且低于根部比值。这可能是在碱胁迫下,碱蓬叶片具有更好的Na+/K+稳态调控能力。
3.3 有机酸等溶质的渗透调节、离子平衡作用
植物保持低Na+高K+是维持渗透调节的主要机制[20]。虽然盐、碱胁迫Na+对渗透调节的贡献变化大,但是K+的贡献依然保持较高水平(仅次于Na+),碱胁迫下叶片K+对渗透调节的贡献提高,Na+的贡献下降。可见,碱蓬对Na+和K+的吸收机制是独特的[21],这可能与其具有较强的抗盐碱能力有关。
脯氨酸对渗透调节的贡献很小,这与碱地肤(Kochiasieversiana)的情况一致[13]。而与盐柳(Salixpsammophila)[22]、酸枣(Ziziphusjujubavar.spionsa)[23]等植物的报道不同,碱蓬体内脯氨酸不是对渗透胁迫的简单反应,它可能是由于盐碱胁迫对其代谢的干扰或是其起到保护响应的作用[20]。
盐、碱胁迫下根和叶片的有机酸贡献提高(仅次于Na+,K+),特别是高pH的碱胁迫下叶片其贡献率增加幅度更大。可见,有机酸的代谢与pH密切相关。考虑到有机酸在细胞中的定位(存在液泡中),可以认为,盐、碱胁迫下有机酸为根、叶片液泡中主要的有机渗透调节物。而多数双子叶盐生植物主要吸收Na+,Cl-等无机离子进行渗透调节[13,24],这可能是因为碱胁迫(高pH)对无机阴离子的吸收有抑制作用[3],碱蓬不得已选择更为耗能的合成有机酸的方式进行渗透调节,同时以调节细胞内pH。
猜你喜欢
土壤学报(2022年4期)2022-10-22
初中生学习指导·中考版(2022年1期)2022-02-09
金桥(2021年11期)2021-11-20
现代畜牧科技(2021年4期)2021-07-21
课程教育研究(2017年20期)2017-06-14
中学生数理化·中考版(2017年12期)2017-04-18
西藏科技(2016年8期)2016-09-26
中国麻业科学(2015年5期)2015-12-28
烟台果树(2015年3期)2015-12-08
创新作文(3-4年级)(2015年11期)2015-11-28
