
甜菜NADP-ME基因家族的生物信息学分析
2021-10-17 01:19李思琪迪娜尔叶尔兰周婉婷汪曼李佳佳刘大丽兴旺常广玉
中国糖料 2021年4期
李思琪,迪娜尔·叶尔兰,周婉婷,汪曼,李佳佳,刘大丽,兴旺,常广玉
(1.国家甜菜种质中期库/黑龙江大学,哈尔滨 150080;2.黑龙江大学现代农业与生态环境学院/黑龙江省普通高等学校甜菜遗传育种重点实验室,哈尔滨 150080;3.穆棱市农业技术推广中心,黑龙江牡丹江 157500)
0 引言
苹果酸参与多种代谢途径,有几种酶对其进行代谢。其中一种重要的苹果酸代谢酶是NADP-苹果酸酶(NADP-ME)。NADP-ME 是一类广泛存在于原核和真核生物中参与生物体生命活动中多种代谢过程的酶。NADP-ME 主要存在于叶绿体、线粒体和胞质中,并通过二价金属离子催化苹果酸氧化脱羧,产生丙酮酸、CO2和NADPH。同时,产生的NADPH也可以作为其它合成反应的还原剂[1]。
NADP-ME普遍存在于植物的不同组织中,因此被认为具有不同的体内调控功能。NADP-ME在植物的光合作用中发挥着重要作用,通过释放二氧化碳以用于RuBisCo酶(加氧酶)的固碳,从而减弱植物的光呼吸作用,减少氧气的消耗,使植物保持较高的光合效率[2]。除此之外,植物受伤或暴露于UV-B 辐射可诱导NADP-ME 的表达[3]。通过这种方式,该酶为木质素生物合成中两个NADPH 依赖的还原步骤提供NADPH,并参与木质素相关的生物防御。另外,NADP-ME 也可为类黄酮生物合成途径提供还原力。丙酮酸是NADP-ME 反应的产物之一,它可在线粒体中通过呼吸作用获得ATP,并可作为磷酸烯醇式丙酮酸(PEP)合成的前体。PEP 促进苯丙氨酸等芳香族氨基酸的合成,是木质素和类黄酮合成的共同底物。此外,NADPME可参与活性氧的合成机制[4],借以破坏病原体而产生。因此,NADP-ME水平的增加可为防御物质的生物合成提供基础和能量,这也进一步说明苹果酸代谢在植物逆境防御中发挥了重要作用。
根据光合作用中的碳同化途径,植物分为C3、C3-C4、C4和CAM 四类[5]。根据功能和亚细胞定位的不同,植物体内的NADP-ME 被分为光合型和非光合型两种[6]。结合基因定位及功能[7],目前NADP-ME 被分为以下几类:C4植物维管束鞘细胞叶绿体型、C4植物质体型、C4植物细胞质型;CAM植物光合型(细胞质型)、CAM植物非光合型(细胞质型);C3植物质体型、C3植物细胞质型。其中光合型为C4植物维管束鞘细胞叶绿体型和CAM 植物光合型,其主要功能是脱羧产生CO2用于RuBisCo 酶的碳固定及类似的作用[8]。除这两类外,其它NADP-ME 均属于非光合型,通过提供CO2、NADPH 和苹果酸等来调节植物渗透压、细胞内pH、促进果实成熟、气孔运动以及脂肪酸的合成[9],影响逆境相关的细胞生理功能。
在植物体中,NADP-ME基因家族较小,家族成员的数量因植物不同而异[10]。虽然NADP-ME基因家族各成员间的同源性较高,但其基因表达模式和生理生化特性差异较大[11],因而在植物体内也发挥着不同的调节功能。其中,植物胞质型NADP-ME 主要参与其逆境防御及相关代谢活动[12]。甜菜(Beta vulgarisL.)属于C3作物,在我国东北、西北及华北地区种植,是我国北方重要的经济作物和糖料作物之一。近年来人们对不同碳同化途径植物的NADP-ME基因家族进行了一定的功能分析[13],但对于甜菜的NADP-ME基因家族信息及其功能了解甚少。本文以全基因组信息已解读的甜菜为对象,利用生物信息学对甜菜NADP-MEs家族成员的序列特征、蛋白理化性质、亚细胞定位、跨膜结构域、蛋白质基序及进化关系等进行预测,为研究BvNADP-ME基因家族的生理功能提供基础信息,并为进一步提高甜菜对逆境的耐性和品质提供新途径。
1 试验方法
1.1 甜菜NADP-MEs基因的挖掘和蛋白的理化性质分析
拟南芥种质信息数据库(http://www.arabidopsis.org/)检索拟南芥NADP-ME基因家族蛋白质序列。利用拟南芥NADP-ME(At1g79750)氨基酸序列进行Pfam(http://pfam.sanger.ac.uk)分析。在NCBI(https://www.ncbi.nlm.nih.gov/)数据库中进行BLASTp 同源检索,最终筛选并获得甜菜NADP-ME家族的基因序列和蛋白序列。使用pI/Mw(http://web.expasy.org/protparam/)对所获得的蛋白序列理化性质进行分析。
1.2 甜菜NADP-MEs基因结构及其染色体定位分析
Ensemblgenomes(http://ensemblgenomes.org/)分析获得甜菜NADP-MEs家族的基因组DNA 及其CDS 序列,并利用GSDS(http://gsds.cbi.pku.edu.cn/)绘制基因结构图。利用Ensemblgenomes 数据库获得基因所在染色体的位置、染色体长度等信息;通过MG2C在线网站获得甜菜NADP-ME基因家族的染色体定位。
1.3 甜菜NADP-ME家族基因蛋白的二、三级结构预测
利用SOPAM在线网站,对蛋白质的二级结构进行预测,并利用SWISS-MODEL(https://swissmodel.expasy.org/interactive)构建甜菜NADP-ME家族基因蛋白的三级结构[14]。
1.4 甜菜NADP-MEs的跨膜预测和导肽分析
通过PSORT(https://www.genscript.com/)对甜菜NADP-MEs序列所编码的蛋白进行亚细胞定位;并用TMHMM 在线网站对家族成员进行跨膜结构域分析;最后用TargetP1.1 Server对甜菜NADP-MEs基因所编码的蛋白进行导肽分析。
1.5 甜菜NADP-ME家族蛋白的结构序列比对及进化树构建
利用DNAMAN 对获得的甜菜NADP-ME 家族蛋白的氨基酸序列进行同源序列比对和保守结构域的分析。利用MEGA7 对拟南芥、菠菜、藜麦以及甜菜的NADP-ME 蛋白质序列进行多序列比对,绘制无根进化树,构建邻接进化树。
1.6 甜菜NADP-ME家族基因蛋白的基序分析
将已分析获得的BvNADP-MEs氨基酸序列,利用MEME分析和预测保守基序,基序设定值为14个。
1.7 甜菜NADP-MEs基因启动子顺式作用元件分析
以甜菜基因组数据库以及GFF3 文件为基础,利用TBTOOLS 软件获取甜菜NADP-MEs基因上游2 000 bp的启动子序列,提交至Plantcare在线进行顺式作用调控元件的统计分析。
2 结果与分析
2.1 甜菜NADP-ME家族成员基本理化特性
通过NCBI 数据库Blast 分析,共获得5 个甜菜NADP-ME家族成员(表1)。这5 个基因编码了584~641个氨基酸,蛋白质相对分子量为64 505.83 Da(BvNADP-ME1)~70 432.69 Da(BvNADP-ME3)。家族成员中仅有BvNADP-ME5等电点大于7,为碱性蛋白,其余均为酸性蛋白。

表1 甜菜NADP-MEs 蛋白理化性质分析Table 1 Analysis of physicochemical properties of NADP-MEs protein in sugarbeet
甜菜NADP-ME 蛋白的平均亲疏水性都为负值,说明该蛋白家族都属于亲水性蛋白;这些蛋白中,只有BvNADP-ME1和BvNADP-ME5的不稳定系数小于40,为稳定蛋白,其余的均为不稳定蛋白。亚细胞定位预测结果显示,除BvNADP-ME1 和BvNADP-ME2 分别位于内质网、细胞膜和细胞核、细胞质上以外,甜菜NADP-ME家族的大部分蛋白位于线粒体。
2.2 甜菜NADP-ME基因家族的结构分析
如图1 所示,甜菜NADP-MEs成员的基因长度介于6 537 bp(BvNADP-ME2)~19 759 bp(BvNADPME4)。它们均为断裂基因,并且内含子数目较多。其中BvNADP-ME3内含子数目高达19个,其余成员均在16~18个。

图1 甜菜NADP-ME家族的基因结构Fig.1 Gene structure of NADP-ME family in sugar beet
2.3 甜菜NADP-MEs的二、三级结构预测分析
α 螺旋和无规则卷曲是构成甜菜NADP-ME 蛋白二级结构的主要元件,β 折叠和延伸链则较少(表2)。其中只有BvNADP-ME3 的α 螺旋少于40%,而少于40%的都是质体型NADP-ME,导肽分析也证明了这一点。利用SWISS-MODEL 建立了甜菜NADP-ME家族蛋白序列的三级结构。由图2 发现,甜菜NADP-ME家族蛋白的三级结构多是由α-螺旋和无规则卷曲组成的,与二级结构预测(表2)结果一致。其中BvNADPME1、3、5的三级结构非常接近。因此,可以推测这3个蛋白在甜菜体内的功能可能是类似的。
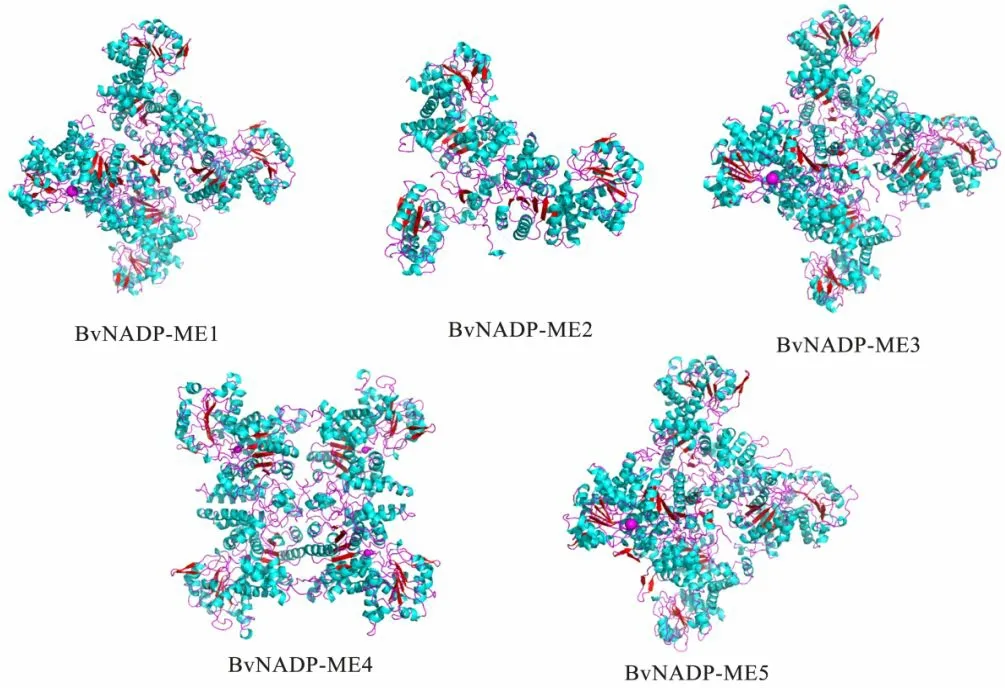
图2 甜菜NADP-MEs蛋白三级结构预测Fig.2 Prediction of tertiary structure of NADP-MEs protein in sugar beet
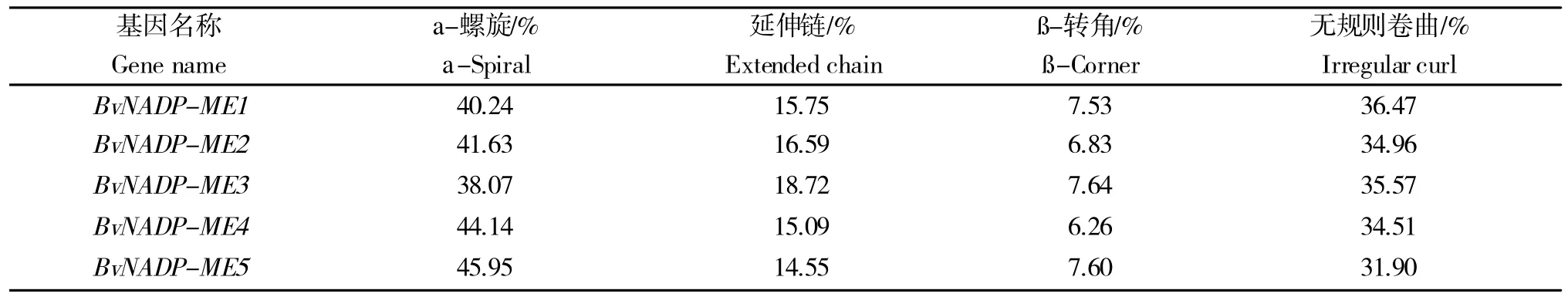
表2 甜菜NADP-MEs 蛋白二级结构分析Table 2 Secondary structure of NADP-MEs protein in sugarbeet
2.4 甜菜NADP-ME基因家族的染色体定位及结构分析
已知甜菜全基因组测序已被测定出的染色体是9 条,通过对所获得的甜菜5 个BvNADP-MEs基因进行染色体定位分析发现,甜菜NADP-ME家族成员(BvNADP-ME5、2、1)主要分布在1、2、8 号染色体上,还有两个基因(BvNADP-ME3、4)分别位于两个零散的片段上,其中BvNADP-ME3位于9 号染色体上的scaffold,BvNADP-ME4位于1号染色体的scaffold,可能是全基因测序未能填补的gap区域(图3)。甜菜NADP-ME基因家族在有限的染色体上分布,也进一步地证明了该家族是一个相对较小的基因家族,其遗传较为稳定。
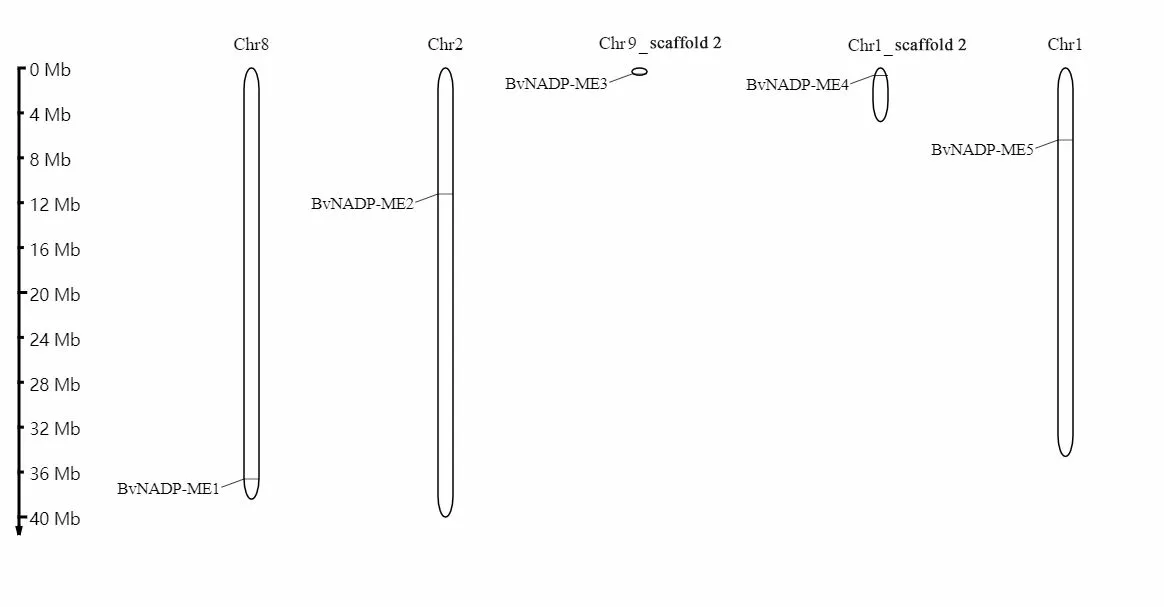
图3 甜菜NADP-ME基因家族的染色体定位Fig.3 Chromosome location of NADP-ME gene family in sugar beet
另外,跨膜结构与分析结果表明BvNADP-ME 家族的5 个蛋白均不存在跨膜结构域,不属于跨膜蛋白。导肽分析结果表明,BvNADP-ME3 含有叶绿体转运肽,BvNADP-ME4、5 均含有线粒体靶向肽。BvNADPME1、2则分别定位在内质网、细胞膜和细胞核、细胞质当中。信号肽预测的结果中5个蛋白均未发现存在信号肽。
2.5 甜菜NADP-MEs序列及进化分析
氨基酸同源性、保守区域及进化关系显示,甜菜NADP-ME 蛋白家族成员之间的氨基酸序列一致性为61.06%,每个成员均有5个保守的NADP-ME氨基酸区域(图4)。其中,氨基酸保守区域I、II、V 是NADP的结合区域,但III和IV 区域的功能尚不清楚[15]。

图4 甜菜NADP-ME蛋白序列同源性及保守结构域分析Fig.4 Homology and conserved domain analysis of NADP-ME protein in sugar beet
如图5所示,为了分析甜菜NADP-ME蛋白家族与拟南芥、菠菜和藜麦NADP-MEs的进化关系,对这4种植物的21 个NADP-MEs 构建了6 个亚组(I~VI)系统发育树。研究发现所有甜菜NADP-MEs 均属于I、II、IV、VI 这4个亚组,其中VI亚组是甜菜所特有的一个亚组,而亚组III是属于拟南芥所特有的一个类群。根据聚类显示,BvNADP-ME1与SoNADP-ME1 同源关系最近。由此可以推测,甜菜与菠菜NADP-MEs亲缘关系相对较近,其次是藜麦,最远是拟南芥。

图5 NADP-ME家族蛋白的系统进化树分析Fig.5 Phylogenetic tree analysis of NADP-ME family protein
2.6 甜菜NADP-ME成员蛋白保守基序分析
甜菜NADP-MEs蛋白包含了14个保守基序,分别命名为Motif 1~14(图6)。其中BvNADP-ME1、2、3包含所有14 个保守基序,BvNADP-ME4、5 则缺少Motif 7、8、13、14。将Motif 的分析结果与保守结构域结果进行比较,发现Motif 2构成I区域,Motif 1构成II区域,这两个区域是NADP的结合区域[15]。Motif3构成III、IV区域,Motif12构成V 区域,并在Pfam数据库中查到Motif1、2、3和motif 9、10都分布在N端,暗示该家族蛋白N端可能具有重要的生物学功能。

图6 甜菜NADP-MEs家族蛋白基序分析Fig.6 Analysis of NADP-MEs family protein motifs in sugar beet
在14 个基序中,Motif4、Motif6、7、8 和Motif 11、12 可能与NAD 的结合有关。但在保守序列分析中提到motif12 构成的V 区域是NADP 的结合区,与Pfam 数据库中分析出的结论有所差异,推断可能是不同碳同化类型的植物其功能区的作用有所不同。另外,Motif 5、13、14 是未知的,在Pfam 数据库未能找到相对应结构域,这3个基序的功能还有待进一步试验证明。
2.7 甜菜NADP-ME基因家族启动子顺式作用元件分析
分析BvNADP-ME基因家族启动子区域的顺式作用元件,结果(表3)显示:各成员启动子均含有植物激素响应元件和非生物胁迫响应元件,其中光响应元件最多,有17 个位点,占总位点数的45.95%,这证明了甜菜NADP-ME基因在光合作用中的重要地位。
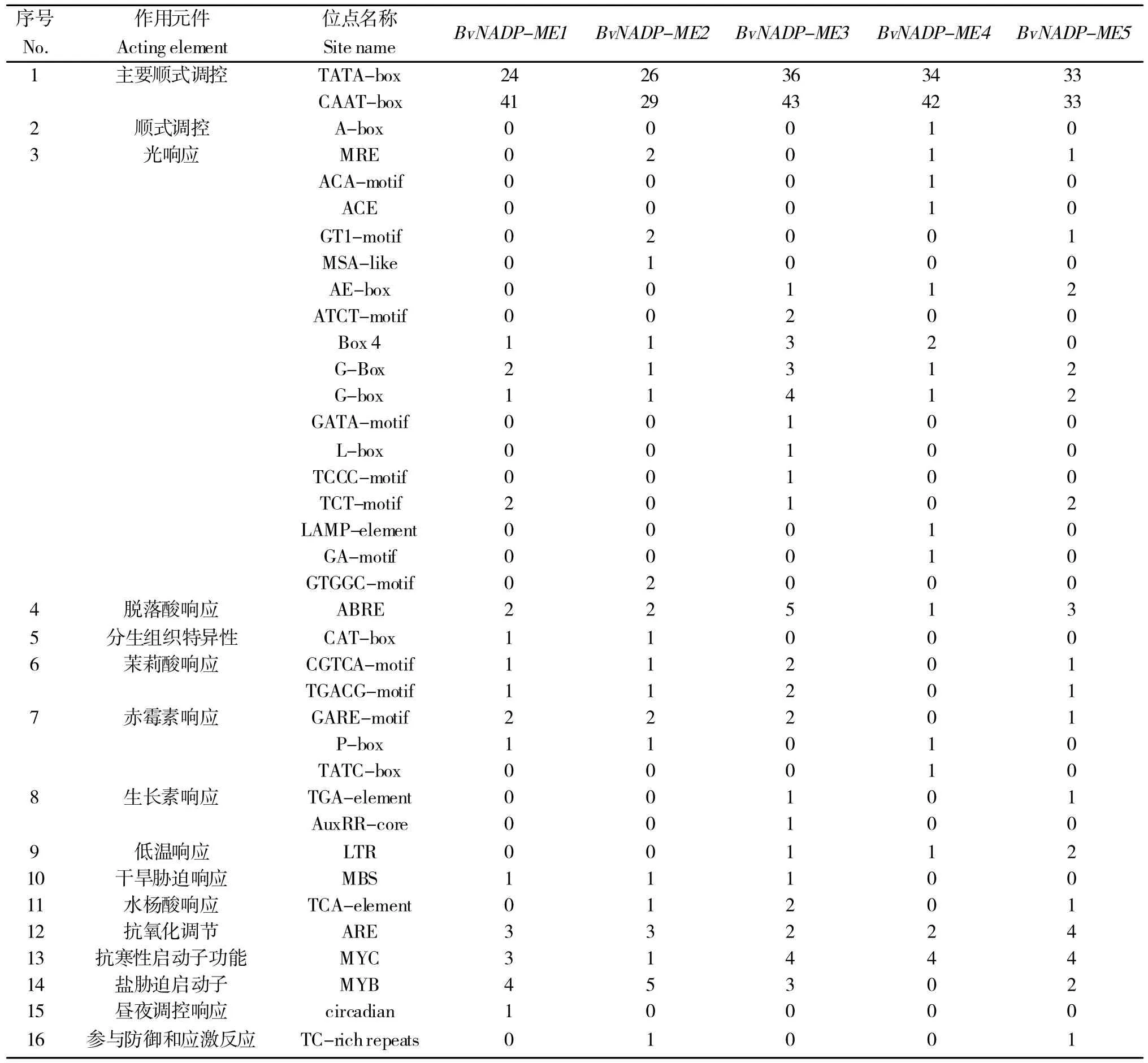
表3 甜菜NADP-ME 基因家族启动子顺式作用元件分析Table 3 Analysis of cis-acting elements of NADP-ME gene family promoter in sugarbeet
在植物生长发育方面,BvNADP-ME基因家族成员启动子区域都含有脱落酸响应元件(Abscisic acid re‐sponse element,ABRE);在非生物胁迫方面,很值得一提的是BvNADP-ME3含有绝大部分的非生物胁迫响应元件,其中包括干旱胁迫响应元件MBS(1 个)、抗氧化调节元件ARE(2 个)、抗寒性启动子功能元件MYC(4个)、盐胁迫启动子元件MYB(3 个)。另外,BvNADP-ME2、5都含有参与防御和应激反应元件。这进一步说明了BvNADP-ME基因家族在植物逆境防御中起到一定的作用。上述结果表明BvNADP-ME家族基因可能参与植物生长发育调节活动并在应对环境胁迫中发挥调控作用。
3 讨论与结论
在自然条件下,植物不断遭遇各种环境的逆境胁迫,这些胁迫可能会限制植物的生长发育,甚至产量。NADP-MEs被认为在植物的不同胁迫响应中发挥重要作用[16]。其生物学功能在多种植物中得到了广泛挖掘,但是该基因家族在甜菜中的信息较为缺乏。因此,利用生物信息学对甜菜NADP-ME基因家族的理化性质、跨膜结构域及同源性、蛋白质基序等进行分析,可为进一步研究BvNADP-MEs的功能表达提供基础信息。
本研究共鉴定得到5 个甜菜NADP-ME基因家族成员。同源检索到与甜菜NADP-ME家族成员同源性较高的菠菜、藜麦和拟南芥,并构建了系统进化树。结果显示,进行比对的21个NADP-MEs基因可分为6个亚组,通过进化分支来看,甜菜NADP-ME家族与拟南芥的亲缘关系较远,与菠菜的亲缘关系较近,这与它们在植物分类上的亲缘关系远近相同(同属藜科二年生草本植物)。另外在系统发育学中存在的几个不同物种的同源对,表明它们可能由共同祖先进化而来,也暗示其在某些信号通路中可能具有相似结构和功能。
亚细胞定位结果表明,BvNADP-ME3、4、5 与跨膜结构域分析结果一致,即BvNADP-ME3 含有叶绿体转运肽,BvNADP-ME4、5均含有线粒体靶向肽;而BvNADP-ME1、2 则分别定位在内质网、细胞膜和细胞核、细胞质当中,但在其信号肽预测的结果中却并未发现存在信号肽,其作用机制及相关功能还有待进一步研究。根据跨膜结构域分析的结果可将BvNADP-ME3定义为非光合型的质体型NADP-ME基因,而根据其亚细胞定位可以将BvNADP-ME2定义为非光合型的胞质型NADP-ME基因。这预示着不同的BvNADP-MEs可能参与或调控不同的信号途径,包括植物发育和逆境响应[16]。
在桃果实发育过程中,发现NADP-ME1和NADP-ME2的表达均随着果实发育而增加。果实硬化期NADP-ME活性的增加为木质素、苯丙素和类黄酮的合成提供了大量的NADPH[17]。另外,氧化应激被认为是导致贮藏种子劣化的主要因素,研究表明拟南芥NADP-ME1功能缺失突变体的种子表现出比野生型更高的蛋白质羰基化水平,与野生型相比,NADP-ME1功能缺失突变体在种子吸胀时,老化种子胚胎中积累了苹果酸和氨基酸[18]。因此,认为NADP-ME1 活性是种子干燥贮藏过程中保护种子不被氧化所必需的。在BvNADP-MEs启动子顺式作用元件分析结果中,我们观察到家族所有成员都具有抗氧化调节元件,这进一步证明了甜菜NADP-ME基因对于种子活力的保持有一定的作用。
以往的研究发现,经过长期的低温诱导,黑麦苹果酸含量或NADP-ME 活性会有所增加;当低温胁迫解除后,苹果酸含量和酶活性均下降[19]。这与甜菜NADP-ME基因家族启动子区域顺式作用元件分析结果一致,试验观察到甜菜NADP-MEs所有成员都含有抗寒性启动子功能元件(MYC)。此结果表明,NADP-ME不仅是一种良好的渗透调节物质,而且还能响应低温造成的渗透胁迫。近年来,越来越多关于NADP-ME蛋白参与植物应激反应的研究被报道:干旱胁迫下,烟草叶片NADP-ME的合成显著增强;但在恢复期间,酶活性又恢复到基础水平[20]。通过启动子顺式作用元件我们确实看到甜菜NADP-ME1、2、3含有干旱胁迫响应元件(MBS)。究其原因,可能是NADP-ME通过在光合碳同化环节催化释放CO2参与干旱胁迫响应。在干旱胁迫下,植物关闭气孔,导致CO2吸收减少,此时增加的NADP-ME可以弥补CO2的不足[21],从而维持碳同化的反应。此外,高盐和脱水诱导了芦荟NADP-ME基因(AvME)的表达,在高盐胁迫诱导48 h 后,AvME 蛋白的表达显著增加,并且随着处理时间的延长,其表达水平逐渐升高[22]。这与顺式作用表达元件分析结果也一致,整个NADP-ME基因家族中,只有NADP-ME4不含有盐胁迫启动子元件。
本研究中,BvNADP-ME1、2的亚细胞定位结果与信号肽结果不符,蛋白质基序Motif12 的功能与在Pfam数据库里查询到的结果有一定差异,这些都需进行进一步的研究鉴定。此外,根据蛋白质基序分析表明,BvNADP-ME 蛋白存在与NAD 结合相关的基序,这暗示着BvNADP-MEs可能参与了糖酵解,糖异生和三羧酸循环等过程,与植物体的代谢途径密不可分。本研究结果不仅为分子抗性育种的发展提供基础,还为利用基因编辑技术增强甜菜抗逆性,进一步提高其产量和品质提供基础信息和研究思路。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
南方医科大学学报(2022年3期)2022-04-13
中国糖料(2022年2期)2022-04-06
中国医科大学学报(2021年8期)2021-09-02
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
实用肿瘤学杂志(2020年4期)2020-12-08
三农资讯半月报(2020年15期)2020-08-25
