
作物株型驯化研究进展
2021-10-15 13:33:34王泉李广丁照华庄洁怡李欣怡赵迪程文
山东农业科学 2021年9期
王泉,李广,丁照华,庄洁怡,李欣怡,赵迪,程文
(1.山东省农业科学院玉米研究所/小麦玉米国家工程实验室/农业部黄淮海北部玉米生物学与遗传育种重点实验室,山东 济南 250100;2.深圳大学生命科学与海洋学院,广东 深圳 518055)
农作物不仅为我们提供食物和能源供应,还为生产活动提供各种原材料,可以说农作物是人类生活和社会发展的最基本要素。目前普遍认为,人类长期对野生植物的驯化形成了当今的作物类群。驯化是一种特殊的、多世代的、长时间的驯化者和被驯化对象之间互利共生、共同进化的关系。在这个关系中驯化者通常对被驯化对象的生长和繁殖产生重大影响以确保其满足驯化者自身的各种需求[1]。很明显,驯化者是人类祖先,被驯化对象则是同一时期的各种野生植物。
在驯化过程中,为了满足人类的需求,被驯化的植物不可避免地产生出一系列区别于其野生祖先的形态和生理特征,这些特征被统称为驯化综合症(Domestication syndrome)[2,3]。植物驯化综合症通常包括植株形态、种子和果实大小、种子传播途径、生育期、种子成熟和休眠等[4]。在人为驯化植物的同时,仍然生长在自然环境中的野生植物也经受着大气、水文、土壤、太阳辐射、岩石矿物以及其它生物等环境因素的影响,自然地为适应新环境而发生改变,最终使得群体的表型发生改变,这个过程被称为自然选择[5]。无论是人为驯化还是自然选择都不可避免地对物种基因型产生影响[6]。由于人工驯化和自然选择的共同作用,绝大多数作物和其祖先品种以及拥有共同祖先的亲缘品种表型迥异、差别明显。而在众多差异表型中最普遍、区分度最显著、对产量影响深远的一个差异是株型差异[7,8]。相关研究表明株型差异往往可以由一些关键基因的变化引起。本文将依次对最重要的三大粮食作物水稻、玉米、小麦的起源和主要株型驯化关键基因(表1)进行综述。同时对如何利用这些驯化基因为分子设计育种提供思路进行展望[9,10]。
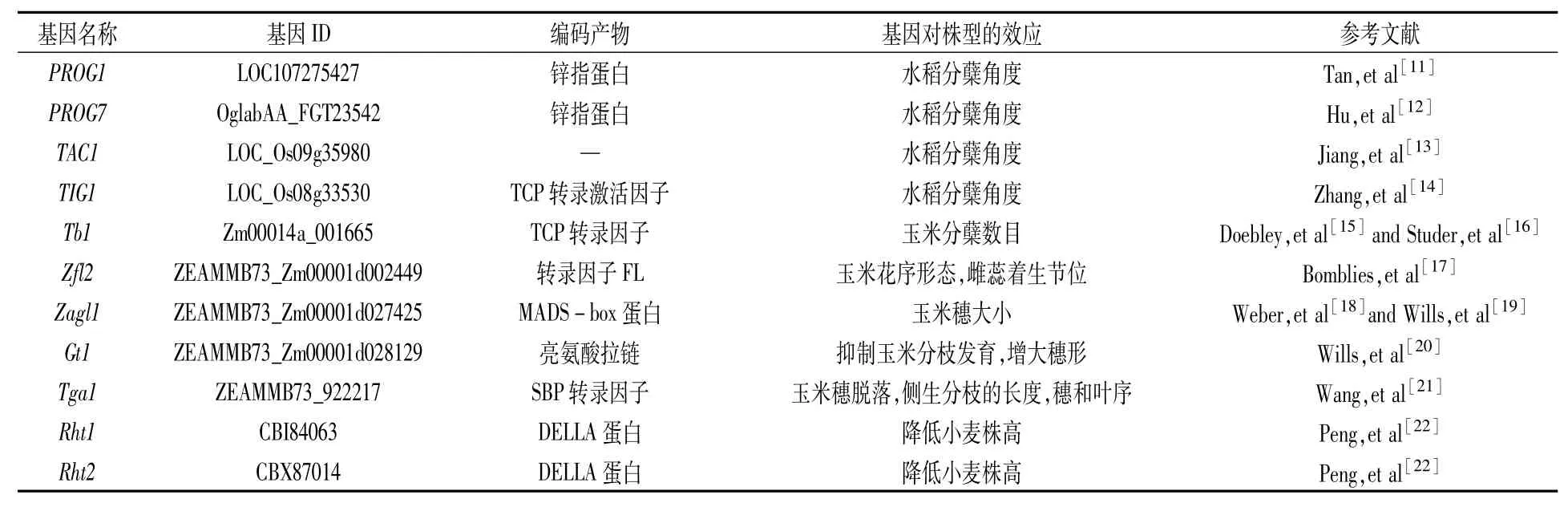
表1 主要农作物株型驯化基因
1 水稻株型驯化研究进展
水稻(OryzasativaL.)是最重要的粮食作物之一,世界范围内超过一半人口以水稻为主食。水稻主要包括亚洲栽培稻(Oryzasativa)和非洲栽培稻(Oryzaglaberrima)两个栽培种[23]。其中非洲栽培稻种植范围相对集中,主要在非洲西部,其起源也比较明确,即由非洲中西部的一年生巴蒂野生稻驯化而来(Oryzabarthii)[24]。而亚洲栽培稻在全球范围内均有分布,可以分为粳稻(Oryza sativassp.japonica)和籼稻(OryzaSativassp.indica)两个亚种[25,26]。关于亚洲稻起源问题目前也是众说纷纭,大体上可分为单一起源说和多起源说两类[26-34]。单一起源说认为粳稻和籼稻都是从野生稻(Oryzarufipogon)驯化而来。多起源说认为粳稻和籼稻分别从亚洲不同地方的野生稻驯化而来,其中粳稻为起源于我国的野生稻[31],籼稻为起源于东南亚地区的普通野生稻或者是尼瓦拉野生稻(Oryzanivara)[25,35,36]。无论是单一起源说还是多起源说都支持普通野生稻是亚洲栽培稻的祖先。
普通野生稻和栽培稻株型上有明显差异。野生稻地上部分匍匐生长,而栽培稻则是直立生长。这种株型上的变化能够有效提高种植密度,增加光截获从而大大提高产量。研究表明从野生稻到栽培稻的这种变化主要是通过改变PROG1基因型引起[11]。该基因编码一个Cys2-His2锌指蛋白,在分蘖基部旺盛表达。改变该基因的基因型可以改变水稻茎秆近地侧细胞数目从而改变分蘖角度。研究者检测了182种直立生长的栽培稻(包括87种籼稻和95种粳稻),表明所有的栽培稻都是prog1基因型;另外检测了30个野生稻,证明大部分匍匐生长的野生稻都是PROG1基因型。prog1和PROG1有15个SNP位点差异和6个indel位点差异。因此,当前普遍认为株型驯化可能是通过对prog1基因型的选择实现的[11]。此外非洲野生稻到非洲栽培稻的株型直立驯化也被证明,是由一个编码锌指蛋白转录因子的PROG7基因的改变引起的。通过分析113个非洲栽培稻和94个非洲野生稻的PROG7基因序列表明,该基因在非洲稻驯化过程中受到强选择。PROG7基因启动子区的4个SNP和2个indel位点改变导致其表达丰度在非洲栽培稻中显著下调,此外编码区也存在30个SNP和5个indel位点差异,这些差异共同导致栽培稻分蘖角度改变,对非洲栽培稻的直立驯化起到重要影响[12]。
另外水稻中还报道了另外两个对分蘖角度和植株形态产生重大影响的基因TAC1和TIG1。其中TAC1在113个栽培稻和48个野生稻中的核酸多态性分析表明,该基因在野生稻、栽培稻品种及籼稻品种中基因型十分保守,核酸多态性几乎不发生变化。但是在粳稻品种中的功能性核苷酸多态性分析表明,该基因在粳稻品种中受到选择压力,从而表明粳稻的株型直立驯化是通过选择TAC1基因实现的[13]。而最近的一项研究表明,籼稻也存在一个特异受到人工选择的且与分蘖角度和植株形态密切相关的基因TIG1。TIG1基因特异地在茎秆远地端高表达,促进EXPA3、EXPB5和SAUR39等下游基因的表达,引起该部位的细胞长度增加,从而形成较大的分蘖角度,改变株型。选择压力分析看出,该基因仅仅在籼稻中受到选择,表明该基因只与籼稻的直立驯化相关[14]。
总的来说,水稻直立株型驯化与4个关键基因的选择相关,其中普通野生稻到亚洲栽培稻的株型改变与PROG1基因相关;非洲野生稻到非洲栽培稻的株型改变与PROG7基因相关;粳稻和籼稻的株型驯化分别与TAC1和TIG1基因相关。
2 玉米株型驯化研究进展
玉米(ZeamaysL.)是世界三大主要农作物之一,不仅是主要的粮食作物,也是重要的饲料和能源作物,同时在工业生产上也占据重要地位。从分类上看,玉米是玉蜀黍属(GemusZea)的一个亚种。玉蜀黍属由玉米黍亚属(SectionZea)和繁茂亚属(SectionLuxuriantes)两个亚属共同组成。玉米黍亚属只有一个种群即玉米种(Zeamays)。但玉米种包含栽培玉米亚种(Zeamaysssp.mays)、墨西哥玉米亚种(Zeamaysssp.mexicana)、委委特南戈类玉米亚种(Zeahuehuetenangensis)以及小颖玉米亚种(Zeamaysssp.parviglumis)等多个亚种。繁茂亚属主要包括繁茂玉米种(Zealuxurians)、多年生玉米种(Zeaperennis)以及二倍体多年生玉米种(Zeadiploperennis)等种群[36]。除了栽培玉米外,其它种群亚群被统称为大刍草(Teosinte)。遗传学证据表明,小颖玉米和栽培玉米有很高的等位基因相似度[37-39],可能是其直系祖先。对不同地区材料分析和遗址考古证明,玉米起源于墨西哥中部,由约9 000年前的一次独立驯化事件而来[39]。但是在玉米扩张传播的过程中墨西哥玉米(Zeamaysssp.mexicana)的许多适应性基因被引入栽培玉米中,表明小颖玉米和墨西哥玉米对现代栽培玉米的基因组都有贡献[40]。因此在提及玉米的祖先时往往用大刍草指代,玉米和大刍草相比有明显的株型差异。
大刍草有分蘖性,具有多个侧生分枝,并且每个侧生分枝都长有一个小穗,穗上既有雌花也有雄花,种子排列通常是二列式;栽培玉米一般不分蘖,顶端只有一个雄穗,侧生分枝数目通常只有一个并且很短,分枝顶端长着雌穗,种子排列通常是多列式[41]。玉米这一系列的株型差异主要与5个关键基因的驯化有关。其中最早被克隆的是tb1基因[15],该基因编码一个含TCP结构域的转录因子,对腋芽的生长起到负向调控作用,从而减少植株分蘖的产生。对大刍草和玉米tb1基因进行遗传分析表明,玉米中该基因的表达量升高了一倍左右。这种表达量的升高是由于约10 000年前tb1基因上游65 kb处插入了一个Hopscotch转座子,并且起到了增强子的作用引起的[16]。另外一个重要基因是zfl2,它是一个受到强选择的一因多效的基因,对玉米的花序和株型都产生重要影响:一方面它能调控花序分生组织形成多级花序(形成大的雄穗),另一方面也影响最高和最低能着生雌穗的节位,同时对侧生分枝数也产生影响[17]。zagl1基因编码一个MADS-box转录因子,能调控玉米开花时间,还表明该基因在玉米驯化过程中被选择用来增加玉米穗的大小和种子的数目[18,19]。gt1基因编码一个亮氨酸拉链转录因子,在玉米驯化过程中受到强选择,它在玉米的一级分枝中高表达,抑制侧枝的生长从而使得玉米的分枝变短[20]。进一步的驯化分析表明该基因在驯化过程中被选择用来减少大刍草的穗数目和增大玉米穗的大小[19]。另外还有一个tga1基因,其主要功能与玉米籽粒的稃壳相关,在驯化过程中受到强选择,在大刍草与栽培玉米中有7个位点差异[21]。但是最新研究表明,该基因也是一个一因多效的基因,对玉米的穗脱落、侧生分枝的长度、穗和叶序的发生都产生影响,并且在驯化过程中被用来选择理想株型[42]。
3 小麦株型驯化研究进展
小麦对保障世界粮食安全起着重要作用。它起源于西南亚地区,最早的栽培遗址可以追溯到7 000年前,栽培于西南亚地区的“新月形沃地”[43]。早期人类在小麦栽培过程中培育出一粒小麦(Enikorn,基因组AA)并且向附近地区传播。一粒小麦在传播过程中与田间杂草(染色体组BB)发生天然杂交,然后再经历染色体加倍后形成野生二粒小麦(Emmer,染色体组AABB)。随后二粒小麦在传播过程中与粗山羊草(Aegilops tauschii,染色体组DD)发生天然杂交,然后经历染色体加倍后形成异源六倍体的普通小麦(染色体组AABBDD)[44,45]。从基因组上来看,六倍体普通小麦应该有三个二倍体祖先,其中两个已经得到确认:乌拉尔图小麦Triticum urartu(基因组AA)和粗山羊草(基因组DD)被认为是A、D基因组的起源物种,但B基因组的起源仍存在争议。由于基因组来源多种,因此本文讨论的小麦株型驯化并不是与其祖先品种进行对比的株型差异,而是主要指20世纪60年代以来的半矮秆化驯化。20世纪60年代,由于人口骤涨,粮食压力增大,育种界掀起一场绿色革命。当时的栽培小麦植株普遍较高,这一方面导致营养生长占用过多的生物量,同时也容易发生倒伏,大大影响作物产量。育种学家们通过杂交方法引入矮秆基因,降低了小麦株高,产量大幅提高[22,46]。
小麦矮秆驯化的关键基因是Rht基因。虽然迄今为止小麦中已经鉴定并且统一命名的Rht基因共有25个(表2),即Rht1至Rht25[47,48],但20世纪60年代的矮秆驯化主要利用的是Rht1和Rht2,这两个基因分别被定位到染色体4B和4D上并且重新命名为Rht-B1b和Rht-D1b[22]。这两个基因都编码DELLA蛋白,该蛋白作为转录抑制因子调控赤霉素信号。在矮化植株中Rht-B1b和Rht-D1b的等位基因型变为赤霉素不敏感型从而抑制植株生长。随后在不同地区的不同小麦品种中鉴定出更多的降秆基因,这些基因对小麦植株的矮化都起到一定作用,但是同时也对穗型、产量、种子营养成分等其它性状产生不同影响,生产上也经常是组合使用来适应不同的育种背景和生长环境。Rht3的矮化等位基因型能降低株高和缩短叶长50%以上[49]。Rht4、Rht5、Rht6、Rht7这4个基因虽然都能降低株高但会降低产量,曾经认为生产应用价值不大,但最近的研究表明Rht4与Rht8共同使用时,与亲本相比能进一步降低株高和提高产量[50]。Rht5能降低株高23%,但会延迟成熟、降低分蘖,严重影响产量[51]。导入Rht8基因能降低株高11%,并且在不同地区不同灌溉条件下不会降低产量,所以也是除Rht1和Rht2之外应用最广泛的矮化基因[52]。Rht9来源于日本赤小麦,能降低株高,同时也能增加11.25%的分蘖数,增产10.11%[53]。Rht10是降秆能力最强的矮化基因,调控小麦株高不到40 cm,但导致地上部分的生物量极低,极大地影响产量[54]。Rht11是一个类似于Rht1的赤霉素敏感型隐性基因,矮化能力也十分出色[55]。Rht12能降低株高43%左右,是一个赤霉素敏感型隐性基因,会延长抽穗时间[56]。Rht13能降低株高24%左右,同时也能提高单株粒数和产量[57]。Rht14、Rht16和Rht18三个基因也是赤霉素敏感型,分别能降低株高47.3%、39.9%和36.0%,并且这三个基因都是通过降低穗下第一节间长度来降低株高[58]。Rht15基因是从Durox品种中通过EMS诱变出来的一个半显性矮化等位基因[47]。Rht17是一个DELLA蛋白编码基因的等位基因,在178个核苷酸处发生C-to-T的碱基替换导致编码提前终止,该基因型能使面包小麦和硬粒小麦株高分别降低约30%和50%[59]。Rht19是一个通过EMS诱变出来的赤霉素反应敏感型矮化等位基因[60]。Rht20是通过γ-射线诱变而来的赤霉素敏感型矮化等位基因[61]。Rht21是一个半显性、能降低株高13.8%且不会引起其它不利于产量的表型发生并具有良好育种潜力的矮化等位基因[62]。Rht22是一个能降低株高约42%左右的隐性基因,它通过减少节间细胞数目实现矮秆[63]。Rht23是一个通过EMS诱变NAUH164品种而产生且具有降低株高、缩短穗长等优点的矮化等位基因[64]。Rht24是一个广泛存在于小麦中的矮化基因,能降低株高8.0%~10.4%,同时对产量起到积极作用[65]。Rht25是最新鉴定的矮化基因,矮化效应约为广泛利用的Rht1和Rht2的一半左右,是一个对胚轴长度、抽穗期、穗长、小穗数、小穗密度和粒重有显著多效性影响的矮化基因[66,67]。
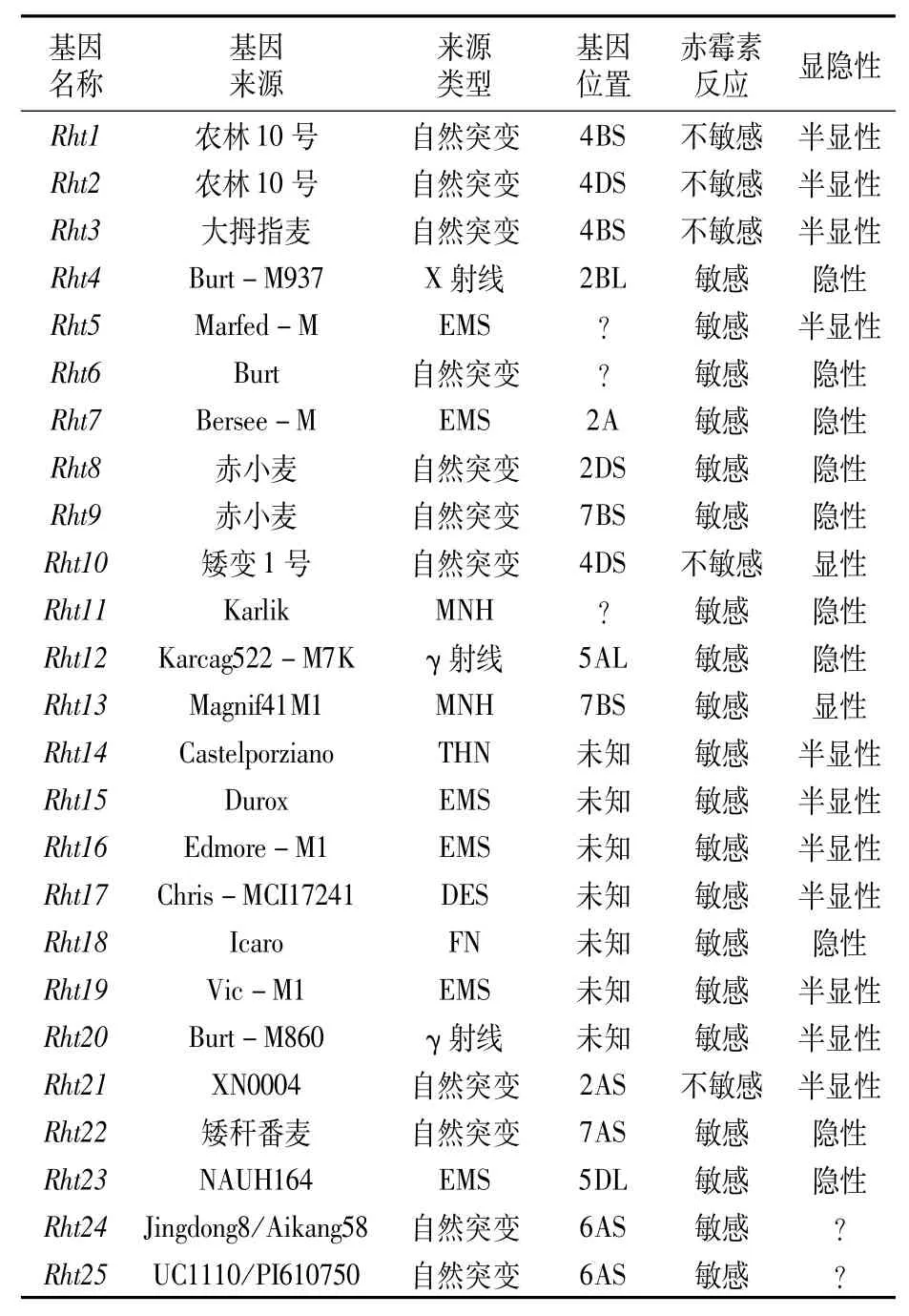
表2 小麦矮秆基因Rht1到Rht25
总的来说,小麦矮化株型的驯化是通过对Rht基因的选择实现,20世纪60年代绿色革命的基因主要是指Rht1和Rht2。到目前为止小麦中共鉴定出25个矮秆基因,它们都能对小麦株高起到抑制作用,具有育种应用潜力。但是目前这些基因在育种上的应用并不普遍,主要用的是Rht1、Rht2、Rht8和Rht11,但对小麦矮化基因进行总结和归纳有利于为后续小麦设计育种提供株型设计思路。
4 展望
植物株型是人类辨识、区别和分类植物的主要标准,也是植物长期适应环境和获取竞争优势的生长策略,更重要的作用是帮助植物更好地繁衍和传播种子。植物独特的外观形态和结构特征是植物按特定程序表达内源基因、遵循特有模式发育的结果。在作物中原本也是如此,但是由于人类活动的介入,一些野生植物的栽培环境发生变化,然后结合人类长期的选择这些野生植物内源基因原有的表达程序也发生了改变,使得它们能更好地适应人类创造的栽培环境、耕作技术并且收获足够的食物和物质材料。这些植物也就逐渐变为人类广泛种植的作物,这个过程也就是作物驯化。
人类祖先的驯化活动改变了作物地上部分的外观形态和结构特征,这些改变为他们挑选出了符合当时耕地环境、耕作技术并且能提供足够食物的作物。但时至今日耕地环境、耕作技术、粮食及有关物质材料的压力一直发生着改变,比如耕地面积的显著减少、土壤肥力的下降、盐碱程度增加、农业机械化的推进、人口增长速度远大于粮食增长速度等。这就迫使我们必须进一步改造现有作物株型以适应现阶段的各种需求。随着1968年澳大利亚育种家C.M.Donald首次提出理想株型(ideotype)概念后,世界各地研究机构和育种家们相继就各种作物提出理想株型标准。比如:国际水稻研究所(International Rice Research Institute,IRRI)提出水稻的理想株型应该是几乎不存在无效分蘖、每穗籽粒较多以及茎较粗抗倒伏等;玉米的理想株型应该为塔型,穗上叶片直立上冲夹角20°~25°,穗下叶角度为40°~45°,且保证所有叶片都能直接接受光照,也不阻碍授粉;小麦的理想株高应该是70 cm左右,且保证地上部分能提供足够的生物量但又不浪费生物量、不倒伏等。通过遗传学和分子生物手段,我们已经发现并且克隆了众多人类祖先驯化选择株型时使用的关键驯化位点和基因。通过总结和归纳这些基因的功能和对表型的影响可以为培养理想株型作物提供参考。并且随着多基因聚合、基因编辑等技术的发展,分子设计育种已逐渐成为主流的育种方法。通过综述株型驯化基因和有效单倍型可以直接通过分子设计育种提供基因组合思路,提升株型育种效率。
猜你喜欢
作物学报(2022年7期)2022-05-11 12:58:02
河北果树(2021年4期)2021-12-02 01:14:52
河北果树(2020年1期)2020-02-09 12:31:18
农村百事通(2019年17期)2019-10-08 02:24:55
中国农业信息(2017年12期)2017-09-03 04:33:01
老友(2017年7期)2017-08-22 02:36:36
上海农业学报(2017年4期)2017-04-10 12:40:28
现代农业科技(2017年1期)2017-03-06 23:08:18
江苏农业科学(2016年8期)2017-02-15 19:19:29
江苏农业科学(2016年8期)2017-02-15 19:19:15
