
不同抚育措施对衰老火炬树叶片生理的影响
2021-10-14 05:02年晓晨王冲冲贾占稳刘桂林
西北林学院学报 2021年5期
年晓晨,王冲冲,贾占稳,刘桂林
(河北农业大学 园林与旅游学院,河北 保定 071000)
火炬树(Rhustyphina)是优良的观赏绿化树种,具有较强的抗逆性以及良好的保持水土能力[1],对生态环境有改善土壤理化性质的效果[2],主要用于荒山和盐碱荒地绿化[3],火炬树可根蘖繁殖,扩散性极强,但通常生长20多a就会出现衰弱或死亡,是一种典型的“短命树”[4]。
近年来已有对火炬树的生物量特征[5]、克隆分株特性[6]、火炬树与乡土树种生理特性的比较[7]、各种胁迫条件下火炬树的生理响应[8-9]等方面的研究。国内外有关学者都注意到火炬树根蘖扩散能力极强的现象,张明如等[10]对火炬树克隆子株水平侧根的研究发现,火炬树具有克隆子株远端水平根直径大于近端水平根直径的不对称生长特性,火炬树的养分呈现出预先储备的特征,为克隆分株不断向外扩散提供一定的物质基础。目前针对不同抚育措施处理控制克隆分株分取母树养分及对衰老母树回缩修剪的研究鲜有报道。
本研究以12年生衰老火炬树为对象,分析不同抚育措施处理对衰老火炬树叶片质膜透性、渗透调节物质以及有关保护酶活性的的影响,探讨不同抚育措施对火炬树叶片生理的影响,为火炬树延长寿命、科学的养护管理提供理论依据。
1 材料与方法
1.1 试验材料与方法
2019年6月10日于保定市植物园选取长势均匀、树龄为12 a(生长锥法测定)的衰老火炬树20株为研究对象。分别于抚育措施处理的第0、30、60、90、120天共5次进行取样,每组选取5株长势、胸径、生长环境相差不大的树,选择树冠外层生长状态相近、无病害损伤的当年生成熟叶片作为试验材料。采集的叶片用密封袋密封,立即放入液氮中再放到-80℃冰箱保存待测,测定时间为每次取样的第2天。
试验共设置4组处理,CK为空白对照,D1为除萌蘖处理组,将主树2 m范围内萌蘖苗全部剪除,之后每隔15 d将新萌蘖苗剪除;D2为修剪处理组,将冗杂枝、干枯枝、过密枝疏除,树冠回缩;D3为断根处理组,距树干50 cm处画圆,沿弧线分6段,每隔1段断根处理,共断1/2的根;每组5株,每株3次重复。
1.2 测定指标
叶绿素含量、超氧化物歧化酶 (SOD) 活性、过氧化物酶 (POD) 活性、过氧化氢酶(CAT)活性、丙二醛(MDA)含量、可溶性蛋白含量、可溶性糖含量测定参照[11]。
1.3 数据处理与分析
试验数据用Excel2019进行初步计算处理并作图,用SPSS21.0进行方差分析和隶属函数分析。
2 结果与分析
2.1 不同抚育措施对衰老火炬树叶片生理的影响
2.1.1 不同抚育措施对叶绿素含量的影响 叶绿素含量的高低可在一定程度上反映火炬树光合作用强弱的变化[12]。衰老火炬树经过不同抚育措施处理后,叶绿素含量变化见图1。各组叶绿素含量随处理时间增加呈现先增高后降低的变化趋势,3组处理叶绿素含量均>CK,在处理60 d达到最高值,D1叶绿素含量始终显著高于CK和其他处理(P<0.05)。

注:不同小写字母和大写字母分别表示不同处理间和不同处理时间上差异显著(P<0.05)。下同。图1 抚育措施后不同时间叶片叶绿素含量变化Fig.1 The variation of chlorophyll content in the leaves at different time after tending
处理前期,在30 d时,D1、D2、D3叶绿素含量分别比CK显著增加了43.45%、19.76%和6.51%(P<0.05)。随着时间的延长,处理中期60 d时,D1、D2、D3叶绿素含量显著>CK(P<0.05),分别比CK增加了44.78%、27.07%、16.33%。处理后期90~120 d,D1、D3叶绿素含量始终显著高于CK(P<0.05),分别比CK增加60.52%/10.27%和77.74%/14.76%,说明在处理后期D2对叶绿素含量的影响较小,D1、D3可以显著提高衰老火炬树叶片的叶绿素含量,提高光合效率。
2.1.2 不同抚育措施对超氧化物歧化酶(SOD)活性影响 超氧物歧化酶(SOD)可消除自由基,维持活性氧代谢平衡[13],其含量越高,表明植物抗氧化胁迫能力越强。超氧化物歧化酶SOD活性在不同抚育措施处理下变化见图2。随着处理时间增加各组SOD活性先升高后降低,在处理60 d时含量最高,差异显著性检验结果表明,各处理间均达到了差异显著水平(P<0.05)。 处理后,3组处理的SOD活性均显著>CK,D1的SOD活性始终显著高于CK和其他处理。处理前期,在30 d时,D1、D2、D3的SOD活性均显著>CK(P<0.05),分别比CK提高了41.43%、39.88%和10.81%。到处理中期60 d时,D1、D2、D3 SOD活性显著>CK(P<0.05),分别比CK提高了22.10%、9.00%、6.54%,此时各处理SOD活性到达最高值。处理后期90~120 d时,D1比CK显著提高了36.54%和12.58%(P<0.05),较其他2组增幅较大。说明D1、D2、D3均可以显著提高衰老火炬树叶片的SOD活性,增强树体抗氧化胁迫能力,相较于D2、D3,D1对SOD活性影响更大。
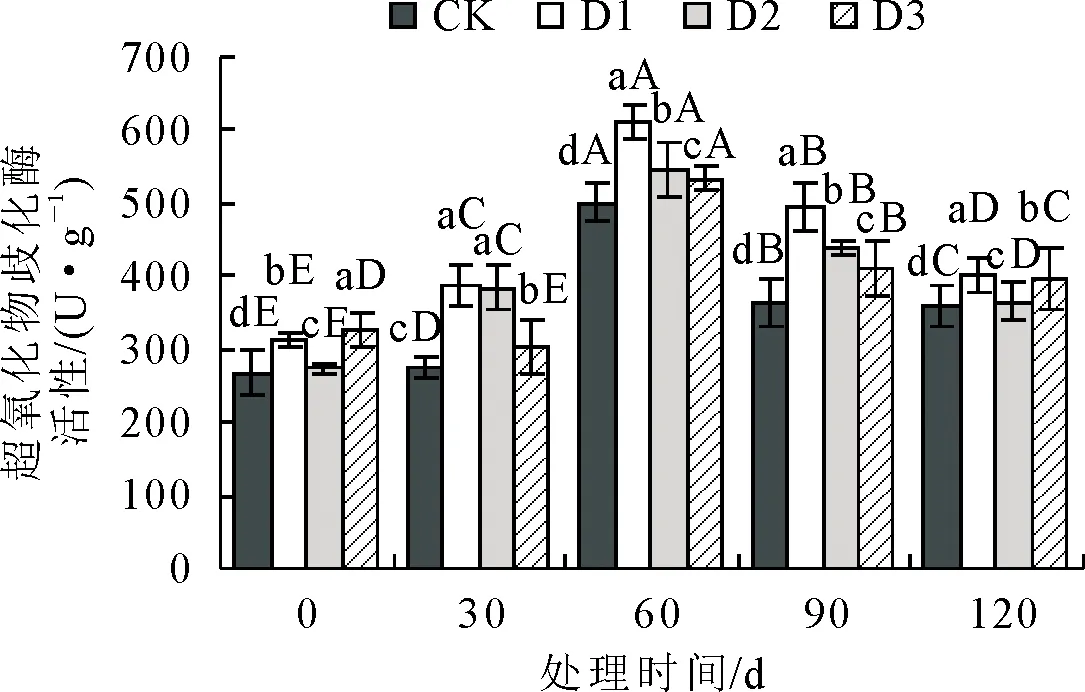
图2 抚育措施后不同时间叶片超氧化物歧化酶(SOD)活性比较Fig.2 The variation of super oxide dismutase (SOD) activity in the leaves at different time after tending
2.1.3 不同抚育措施对过氧化物酶(POD)活性影响 过氧化物酶(POD)活性在不同抚育措施处理下变化见图3。随着处理时间增加POD活性先升高后降低,在处理后,差异显著性检验结果表明,各处理间均达到了差异显著水平(P<0.05),D1的POD活性始终高于CK和其他处理(P<0.05)。处理前期,在30 d时,D1、D2、D3分别比CK显著增加了39.92%、11.07%、27.62%(P<0.05)。各处理POD活性在处理60 d时最高,分别比CK显著增加了73.27%、31.41%、44.09%(P<0.05)。处理后期90~120 d时,D1、D2、D3 POD活性较处理60 d时有所降低,仍显著>CK(P<0.05)。说明处理30 d起,D1、D2、D3均可以显著提高衰老火炬树叶片的POD活性,增强树体抗氧化胁迫能力,其中D1对POD活性影响更大。

图3 抚育措施后不同时间叶片过氧化物酶(POD)活性比较Fig.3 Comparison of peroxidase (POD) activity in the leaves at different time after tending
2.1.4 不同抚育措施对过氧化氢酶活性(CAT)影响 过氧化氢酶(CAT)活性在不同抚育措施处理下变化见图4。各组植株CAT活性变化随处理时间增加先升高后降低,通过D1、D2、D3处理后,CAT活性均比对照组显著增大(P<0.05),说明3组处理可不同程度增强衰老火炬树叶片CAT活性。在处理前期30 d时,D1、D2、D3的CAT活性均显著>CK(P<0.05),分别比CK提高了53.67%、61.48%、67.95%。从处理60 d到120 d,各处理的CAT活性均显著高于CK(P<0.05),其中D1的 CAT活性始终显著高于D2、D3和CK,分别比CK提高了141.31%、113.60%和106.16%。表明3组处理均可显著增强衰老火炬树叶片CAT活性,相较于D2、D3,D1对衰老火炬树叶片CAT活性影响更大。

图4 抚育措施后不同时间叶片过氧化氢酶(CAT)活性比较Fig.4 Comparison of catalase (CAT) activity in the leaves at different time after tending
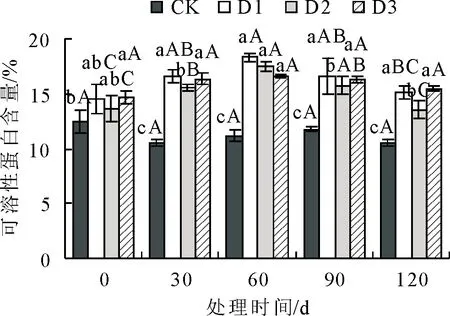
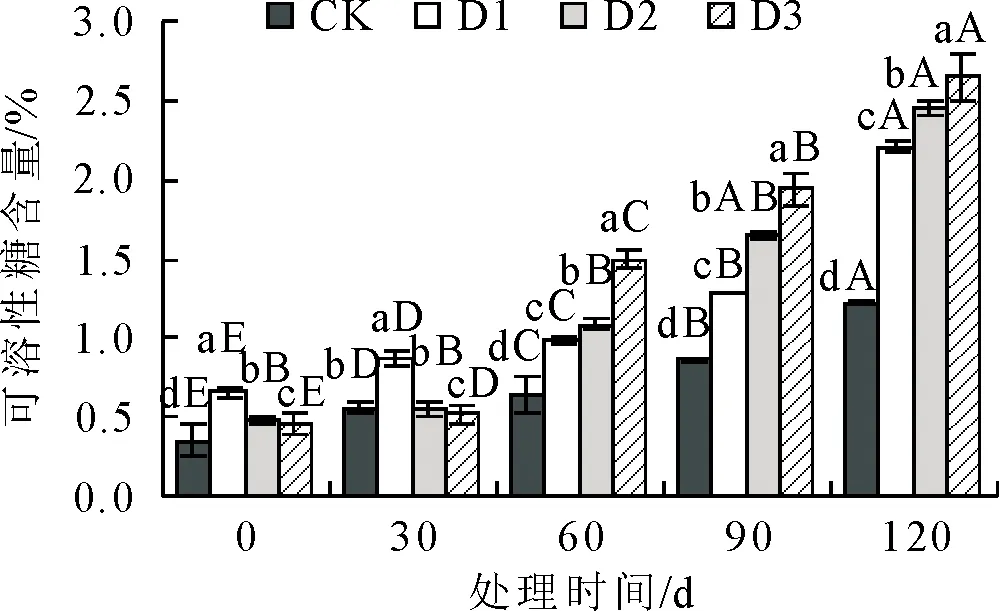
2.1.5 不同抚育措施对丙二醛(MDA)含量影响 丙二醛(MDA)含量在不同抚育措施处理下变化见图5。各组MDA含量随处理时间增加先降低后升高,通过D1、D2、D3处理后,MDA含量均比对照组有所减少。在处理初期30 d时,D1、D2、D3的MDA含量较CK均显著降低(P<0.05)。在处理60 d时,D2的 MDA含量到达最低值,较CK低25.20%,显著 图5 抚育措施后不同时间叶片丙二醛(MDA)含量比较Fig.5 Comparison of malondialdehyde (MDA) content in the leaves at different time after tending 2.1.6 不同抚育措施对可溶性蛋白含量影响 可溶性蛋白含量在不同抚育措施处理下变化见图6。可溶性蛋白含量随处理时间呈先升高后降低变化趋势,3组处理可溶性蛋白含量均比CK显著增多(P<0.05)。从处理初期30 d起,D1、D2、D3的可溶性蛋白含量均显著>CK(P<0.05)。处理中期60 d时,3组处理可溶性蛋白含量达到最高值,分别比对照增加了39.05%、36.12%、32.77%。到处理后期90 d时,各组可溶性蛋白含量较60 d无明显变化,处理120 d时D1、D3可溶性蛋白含量仍处于较高水平,D2可溶性蛋白含量较处理中前期显著降低(P<0.05)。综上所述,D1、D3对衰老火炬树叶片可溶性蛋白含量影响效果较D2更为明显。 图6 抚育措施后不同时间叶片可溶性蛋白含量比较Fig.6 Comparison of soluble protein contents in the leaves at different time after tending 2.1.7 不同抚育措施对可溶性糖含量影响 可溶性糖含量在不同抚育措施处理下变化见图7,总体呈现升高趋势,从处理60 d起D1、D2、D3植株可溶性糖含量均比对照组显著增多(P<0.05)。在处理前期,处理30 d时,D1可溶性糖含量显著高于D2、D3和CK,较CK增加了35.99%(P<0.05)。从处理中期60~120 d,D1、D2、D3的可溶性糖含量均显著>对照(P<0.05);D3可溶性糖含量显著高于D1、D2和CK,各处理阶段分别比CK增多了56.85%、55.50%、53.70%(P<0.05)。表明3组处理均可显著增加衰老火炬树叶片可溶性糖含量,相较于D1、D2,D3对衰老火炬树可溶性糖含量影响更大。 图7 抚育措施后不同时间叶片可溶性糖含量比较Fig.7 Comparison of soluble sugar contents in the leaves at different time after tending 由表1可知,各类生理指标之间存在一定的相关性,如SOD活性与POD活性、CAT活性等同一类别抗氧化酶活性指标,以及同可溶性蛋白含量、MDA含量等渗透调节物质或膜脂过氧化指标均存在显著或极显著相关性,其他生理指标与SOD活性存在相似相关性现象。 表1 火炬树生理指标的相关性分析Table 1 Correlation analysis of resistance physiological indexes of R.typhina 用单一指标来评价不同抚育措施处理效果比较片面,因此采用主成分分析法对表征火炬树衰老程度相关的7个生理指标进行综合分析。由表 2 可知,在前3个主成分中的方差占总方差比例为88.990%,特征值分别为4.316、1.336、0.547,方差贡献率分别为61.664%、19.508%、7.819%,对应较大向量分别为POD活性、可溶性糖含量、SOD活性。因此,选择这3个指标作为不同抚育措施处理对衰老火炬树的生长状态的综合评价指标。 表2 火炬树叶片生理指标主成分分析Table 2 Principal component analysis of physiological indexes in the leaves of R.typhina 运用隶属函数法对3个指标进行隶属函数值计算。将处理第30、60、90 d和120 d各指标隶属函数值累加,求其平均值,以评价各抚育措施处理对衰老火炬树叶片生理的影响效果。由表3可知,3种抚育措施处理隶属函数平均值排序为D1(0.897)>D3(0.698)>D2(0.584)>CK(0.000),隶属函数值越高,说明抚育措施处理效果越好。表明3种抚育措施中,除萌蘖处理效果最好,断根处理次之,修剪处理效果相对较差。 表3 火炬树叶片生理指标的综合评价Table 3 Comprehensive evaluation results of physiology indexes for the leaves of R.typhina 研究结果表明,除萌蘖处理对衰老火炬树叶片生理特性影响效果最好,其次为断根处理,表明分蘖是造成火炬树衰弱的主要原因。研究结果为火炬树早期预防衰老以及衰老期改善生长状态、延缓火炬树衰老以及今后培育不分蘖的火炬树提供了理论依据。 树木在衰老过程中叶绿体数量、组成成分发生变化,叶绿素发生自我降解或液泡中的降解酶在衰老阶段被运输到叶绿体内部,导致叶绿体被降解,光合效率降低[14]。研究结果显示适当的抚育措施可有效降低叶绿体的分解,提高叶绿素含量,除萌蘖处理下,叶片中叶绿素含量显著>CK,很大程度提高了火炬树的光合能力。 在火炬树衰老进程中,不仅外部形态发生明显变化,如叶片形态变化,树体内部各器官生理特性也发生很大变化[15],例如活性氧积累以及细胞膜质损伤等生理生化反应。随着树体衰老,体内SOD、POD、CAT等抗氧化酶活性降低,自由基产生和清除平衡被打破[16-17],导致自由基过剩,细胞膜脂发生过氧化,细胞结构破坏。对衰老火炬树采用除萌蘖、修剪、断根3种抚育措施可显著提高抗氧化酶活性,强化SOD等保护酶的防御系统,降低MDA等膜质过氧化物对细胞代谢的侵害,有效缓解树体衰老,对延缓火炬树衰老具有实用性。 树体内可溶性糖和可溶性蛋白可以缓解树木衰老进程中膜系统的损伤,是重要的渗透调节物质[18]。研究结果显示,除萌蘖、修剪、断根3种抚育措施均可显著提高衰老火炬树体内可溶性糖和可溶性蛋白含量,表明这3种抚育措施均能促使火炬树积累渗透调节物质,有效缓解火炬树衰老进程中叶片膜系统的损伤。 在本研究中运用模糊数学中的隶属函数法能够较全面地对3种抚育措施对衰老火炬树叶片生理影响效果进行综合评价[19-20]。综合评价结果表明,除萌蘖处理对衰老火炬树叶片生理影响效果最好,这可能是由于除萌蘖处理能有效控制无性繁殖的萌蘖苗与母树竞争养分,从而提高母树生长量,增强树势,延缓了母树衰老进程。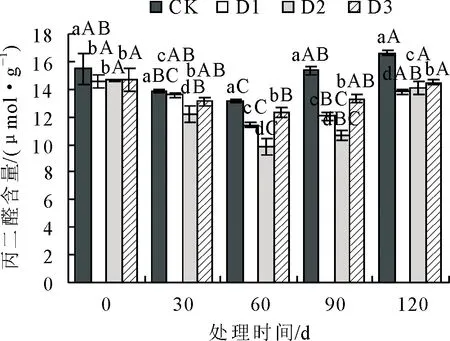
2.2 生理指标间的相关性分析

2.3 主成分和隶属函数分析


3 结论与讨论
3.1 抚育措施对衰老火炬树叶片生理特性的影响
3.2 抚育措施对衰老火炬树生理特性影响效果评价
猜你喜欢
音乐天地(音乐创作版)(2021年7期)2021-10-13
阅读(科学探秘)(2020年8期)2020-11-06
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
Coco薇(2017年5期)2017-06-05
中国火炬(2015年3期)2015-07-31
中国火炬(2015年2期)2015-07-25
中国火炬(2015年2期)2015-07-25
