
定向栽培连作淮山根际土壤细菌群落的多样性分析
2021-09-25 02:49周灵芝劳承英申章佑李艳英黄渝岚韦本辉
西南农业学报 2021年8期
周灵芝,劳承英,申章佑,李艳英,黄渝岚,韦本辉,胡 泊,周 佳
(广西农业科学院经济作物研究所,广西 南宁 530007)
【研究意义】淮山又称山药,是薯蓣科薯蓣属(Dioscorea)一年生或多年生草质藤本植物[1],是我国著名的补益中药,也是菜粮药兼用的高效经济作物[2-4],市场需求旺盛。但淮山连作极易形成连作障碍,造成淮山长势衰弱、病害严重,极大影响淮山的产量和品质,是制约淮山产业健康发展的重要因素,研究解决淮山连作障碍已成为淮山生产上亟需解决的问题。在已有对淮山连作的研究中,较多采用平板培养法统计土壤微生物的菌落数量,但此法获得纯培养土壤微生物菌株的种类数量仅占微生物总数的0.1%~1.0%,难以从根本上揭示淮山连作土壤微生物的群落结构及其变化特征。而高通量测序技术具有高通量、高灵敏度、高准确性和低运行成本等特点,可更全面、更准确地揭示土壤微生物群落的复杂性和多样性[5]。因此,利用高通量测序分析连作淮山根际土壤细菌群落结构,对研究淮山连作障碍具有重要意义。【前人研究进展】在土壤中生存数量巨大、种类繁多的微生物是土壤生态系统的主要组成部分[6],亦是缓解作物连作障碍及其他土壤障碍因子的重要因素[7]。土壤中的有益微生物可促进作物生长发育,有害微生物则会抑制作物正常生长发育[8]。孙凯宁等[9]采用稀释涂抹平板法研究发现,淮山连作破坏了土壤耕层微生物种群结构,其细菌数量随着淮山连作年限的延长呈递减趋势,轮作模式则促进土壤细菌数量增加。马莹[10]研究认为,随着淮山连作年限的延长,土壤微生物的组成和数量排序表现为连作两年土壤>对照土壤>耕作一年土壤。土壤微生物在总体数量上均以细菌为主,随着连作年限的增加,可培养的微生物总量呈上升趋势。王文庆[11]研究表明,在淮山连作时其根际微生物区系的微生物菌群结构发生了由“细菌主导型”向“真菌主导型”转化。乔卿梅等[12]研究显示,根际土壤细菌、放线菌和真菌均随着淮山药生长期的延长呈显著上升趋势,三者在11月的绝对数量分别是7月的2.11、1.70和26.19倍,尤其以真菌的增长速度更快。张红霞等[13]利用高通量测序发现变形菌、绿弯菌、酸杆菌和放线菌是连作山药根际土壤细菌的优势种群,其相对丰度均大于5.0%,占总序列的85.1%。【本研究切入点】淮山定向栽培技术在南方已大面积推广应用[14],但目前利用高通量测序技术研究定向栽培淮山连作对其根际土壤细菌多样性影响的研究鲜见报道。【拟解决的关键问题】通过Illumina HiSeq高通量测序平台探究定向栽培淮山根际土壤细菌群落的结构特征,探明定向栽培淮山连作的细菌群落变化情况,为进一步解决淮山连作障碍提供参考依据。
1 材料与方法
1.1 试验地概况
试验于2018年3月至2020年3月在广西南宁市隆安县那桐镇大滕村试验基地(东经107°52′22″,北纬22°58′11″)进行,土壤类型为红壤,年均气温21.8 ℃,年均降水量约1300 mm,日照充足。
1.2 试验材料
田间种植的淮山品种为桂淮2号,采集其根际土壤为试验材料。桂淮2号由广西农业科学院经济作物研究所选育提供。该品种属于小叶型中晚熟品种,薯块长圆柱形,长50~100 cm,表皮棕褐色、光滑,肉质细腻;单株薯重0.6~1.0 kg,产量约22 500 kg/hm2。
1.3 试验方法
设连作淮山(T)(定向栽培2年淮山)和空白露地(位于T处理旁边,第1年种植淮山而第2年未种植淮山)(对照,CK)两个处理。
1.3.1 土壤样品采集 2020年3月T处理收获期进行土壤样品采集。在试验地内选取20个取样点,拨开淮山定向管的表土层,用手将淮山轻轻取出,抖掉根系上松散的大土块,然后用刷子将与根系紧密结合的土壤刷下作为1个根际土壤样品。土样在田间充分混匀,并剔除根系、杂草等杂物,每个土样分成2份:1份装于无菌离心管,放入装有冰袋的取样箱带回实验室,存放于-80 ℃冰箱,用于土壤细菌群落结构多样性分析;另1份置于室内风干,用于测定土壤理化性质。CK取样时先清除厚约1.0 cm的表层土壤,再挖取第1年淮山种植带中的耕层土壤,去除杂物等操作与T处理相同。T处理和CK各3个重复,共6个样品。
1.3.2 高通量测序 土壤样本委托上海美吉生物医药科技有限公司进行测序,以细菌V3-V4区特异性引物(338F-806R)进行PCR扩增。
1.3.3 测序数据生物信息学分析 测序数据通过Flash和Trimmomatic进行过滤优化和双端序列连接,优化的序列利用Usearch在97%相似水平进行聚类,得到各样品可操作分类单元(OTU)的代表序列,采用Mothur对OTU进行Alpha多样性和β多样性分析,计算Chao l、Ace、Shannon和Simpson指数;利用R语言工具分析和制图(Venn图和群落柱形图)。参考Silva细菌数据库利用RDP classifier进行物种注释和分类。
1.3.4 土壤理化性质测定 参照鲁如坤[15]方法,土壤pH采用酸度计测定,水解性氮含量采用碱解扩散皿滴定法测定,有效磷含量采用钼锑抗比色法测定,速效钾含量采用乙酸铵浸提—火焰光度计法测定,有机质含量采用重铬酸钾硫酸氧化—滴定法测定。
1.4 统计分析
试验数据采用Excel 2010和SPSS 17.0进行统计分析,以Duncan,s新复极差法进行多重比较,以Pearson系数进行相关分析。
2 结果与分析
2.1 连作淮山根际土壤细菌群落高通量文库结果评价
基于细菌16S rRNA基因分析,对6个土壤样品进行Illumina MiSeq高通量测序,经过滤优化,共得到有效序列230961条,序列长度分布在234~531 bp,在97%相似水平上聚类获得3423个OTUs,分属于33门95纲242目395科780属1524种。其中,T处理根际土壤细菌群落获得3076个OTUs,分属于33门93纲237目387科767属1471种;CK根际土壤细菌群落获得2035个OTUs,分属于27门75纲180目273科496属905种(图1)。
由表1可知,T处理和CK土壤样品文库的覆盖率均较高,分别为98.42%和98.91%,说明土壤样品的测序文库已达饱和状态,所构建文库已包含样品中的大部分细菌种类,能较好地反映土壤的细菌群落结构。
2.2 连作淮山根际土壤细菌群落的物种Venn图分析
从图1可看出,T处理和CK根际土壤共有的OTUs数为1688个,特有的OTUs数分别为1388和347个。经方差分析,二者的OTUs数差异极显著(P<0.01,下同)。说明连作淮山根际土壤中细菌群落的OTUs分布与CK相比既有共性又存在极大差异。
2.3 连作淮山根际土壤细菌群落的多样性分析
2.3.1 Alpha多样性分析结果 分别对细菌菌群丰富度指数(ACE指数和Chao l指数)和细菌群落多样性指数(Shannon指数和Simpson指数)进行分析。由表1可知,与CK相比,T处理土壤细菌的Shannon指数极显著上升,升幅达12.8%,而Simpson指数极显著下降,降幅为57.1%。说明连作淮山根际土壤细菌群落的多样性极显著高于CK。
从表1还可看出,T处理土壤细菌的ACE指数和Chao l指数均大幅上升,升幅分别为50.8%和49.4%,与CK差异极显著。说明淮山连作后其根际土壤细菌群落的多样性和丰富度都发生了显著变化。
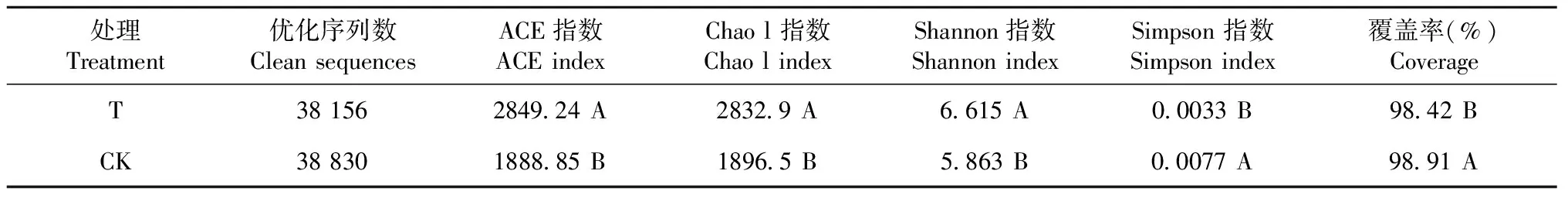
表1 连作淮山根际土壤的细菌多样性指数
2.3.2 β多样性分析结果 从图2可看出,T处理和CK土壤样品分布离散、距离较远,表明二者土壤样品间的细菌群落结构差异较明显;第一主成分(PC1)对细菌群落结构的差异性贡献率为81.44%,T处理土壤样本点与PC1轴距离较近,说明连作淮山土壤根际细菌菌群结构受PC1影响较大;第二主成分(PC2)对细菌群落结构的差异性贡献率为14.49%,CK土壤样本点与PC2轴距离较近,说明CK土壤细菌菌群结构受PC2影响明显。可见,本研究中土壤细菌群落的PC1和PC2能解释95.93%的样品间差异性,连作淮山根际土壤的细菌群落结构与CK存在明显差异。
2.4 连作淮山根际土壤的细菌群落组成分析
2.4.1 细菌优势菌门的分析结果 在门分类水平上连作淮山根际土壤的细菌群落隶属于33门,CK土壤的细菌群落隶属于27门。从图3可看出,放线菌门(Actinobacteria)、变形菌门(Proteobacteria)和厚壁菌门(Firmicutes)等12个已知菌门(相对丰度>1.00%的菌门属于优势菌门)在T处理和CK土壤细菌群落中的占比分别为98.04%和98.55%,其中,T处理土壤细菌群落中相对丰度排在前10位的优势菌门依次为放线菌门(26.86%)、变形菌门(25.46%)、厚壁菌门(12.62%)、绿弯菌门(Chloroflexi)(10.62%)、酸杆菌门(Acidobacteriota)(7.72%)、拟杆菌门(Bacteroidota)(5.17%)、粘球菌门(Myxococcota)(3.59%)、芽单胞菌门(Gemmatimonadota)(1.82%)、浮霉菌门(Planctomycetota)(1.49%)和Patescibacteria(1.36%),而CK土壤细菌群落中相应细菌门的相对丰度分别为30.55%、11.99%、8.69%、31.79%、7.32%、0.27%、0.83%、12.83%、2.34%和0.88%。说明连作淮山根际土壤细菌群落组成在门水平上与CK存在较大差异。
门分类水平上对物种相对丰度进行方差分析,结果(图4)表明,在T处理土壤细菌群落相对丰度排前12位的优势菌门中,变形菌门、绿弯菌门、拟杆菌门、粘球菌门、Patescibacteria、疣微菌门(Verrucomicrobiota)和WPS-2等7个优势细菌门的相对丰度与CK存在显著差异(P<0.05,下同)。其中,变形菌门、拟杆菌门、粘球菌门、Patescibacteria和疣微菌门等5门的相对丰度显著大于CK,绿弯菌门和WPS-2显著小于CK。说明连作后淮山根际土壤优势细菌在门水平上的相对丰度与CK存在较大差异。
2.4.2 细菌优势菌属的分析结果 在属分类水平上连作淮山根际土壤的细菌群落隶属于767属,其中,相对丰度>1.00%的有20属,而CK土壤的细菌群落隶属于496属,相对丰度>1.00%的有25属;相对丰度>1.00%的T处理和CK土壤细菌群落的优势菌属共有37个,其中有18属在分类学数据库中为分类学谱系的中间等级,无科学名称,因此用norank或unclassified标记。由表2可知,T处理排名前20位的优势菌属和丰度与CK存在较大差异,其中,仅有芽孢杆菌属(Bacillus)、慢生根瘤菌属(Bradyrhizobium)、norank_JG30-KF-CM45、热酸菌属(Acidothermus)和Bryobacter同为T处理和CK土壤细菌群落共有的优势菌属,T处理排名前10位的优势菌属均未出现在CK排名前10位的优势菌属中;而T处理根际土壤细菌中出现了有益菌属链霉菌属(Streptomyces)和鞘氨醇单胞菌属(Sphingomonas)。说明淮山连作的优势菌属与CK差异较明显。

表2 连作淮山根际土壤细菌群落排名前20位优势细菌属的相对丰度比较
2.5 淮山连作根际土壤细菌主要群落与土壤理化性质的关系
2.5.1 连作淮山根际土壤的理化性质 由表3可知,T处理和CK的土壤均偏酸性,但T处理土壤的pH为6.33,极显著高于CK(4.83);T处理土壤的水解氮、有效磷和速效钾含量分别为176.23、88.77和1010.70 mg/kg,有机质含量为54.47 g/kg,均极显著高于CK。可见,淮山连作后其根际土壤的理化性质发生了极明显的变化。

表3 连作淮山根际土壤的理化性质
2.5.2 连作淮山根际土壤主要细菌菌群与土壤理化性质的相关分析 对T处理土壤在门水平相对丰度>1.00%的主要细菌群落与土壤理化因子进行Pearson相关分析,结果(表4)表明,在相对丰度排名前12位的主要细菌菌门中,有7个细菌门与土壤理化性质呈显著(P<0.05,下同)或极显著相关。其中,变形菌门与pH及水解氮、速效钾和有机质含量呈极显著正相关,与有效磷含量呈显著正相关;绿弯菌门与pH及水解氮、速效钾和有机质含量呈极显著负相关;拟杆菌门与pH及水解氮、速效钾和有机质含量呈极显著正相关,与有效磷含量呈显著正相关;粘球菌门与pH及水解氮、速效钾和有机质含量呈极显著正相关,与有效磷含量呈显著正相关;Patescibacteria与pH及水解氮、速效钾和有机质含量呈显著正相关;疣微菌门与有效磷含量呈极显著正相关,与水解氮、速效钾和有机质含量呈显著正相关;WPS-2与pH、水解氮、有机质含量呈极显著负相关,与速效钾含量呈极显著正相关。说明pH及水解氮、速效钾和有机质含量是影响连作淮山根际土壤细菌群落组成的重要因子,而速效磷含量对细菌群落组成的影响较小。

表4 连作淮山根际土壤主要细菌菌群与土壤理化性质的相关分析结果
3 讨 论
淮山属于极易发生连作障碍的作物,有些地方淮山连作两年即出现不同程度的烂根现象[11],严重制约淮山产业的可持续发展。前人对连作马铃薯[16]、西瓜[17]、草莓[18]等的研究发现,连作导致土壤微生物环境和微生物群落结构发生改变是其发生连作障碍的原因之一。本研究通过高通量测序对定向栽培连作淮山根际土壤和CK土壤样品进行细菌序列测定,结果发现该2种土壤除存在共有的细菌类群外,还存在具有极大差异的细菌群落,与胡彦婷等[19]对连作胡萝卜的研究结果相似。
土壤样品中的微生物多样性通常以Shannon指数和Simpson指数表示,前者数值越大,表明群落多样性越高,后者数值越大,表明群落多样性越低。细菌菌群的丰富度常用ACE指数和Chao l指数表示,其数值越大表明细菌群落越丰富。本研究中,连作淮山土壤细菌的Shannon指数、ACE指数和Chao l指数与CK相比均显著上升,升幅分别为12.8%、50.8%和49.4%;连作淮山土壤细菌的Simpson指数极显著下降,降幅为57.1%。表明淮山连作可导致其根际土壤细菌群落的多样性和丰富度增加,与前人对连作木薯[20]、烟草[21]、花生[22]等作物的研究结果一致,但与吕昭和等[16]、杨尚东等[23]、周冰谦等[24]研究认为连作会降低作物根际土壤细菌群落多样性的观点不一致,可能与作物种植地的气候、土壤类型、连作年限和田间管理措施不同等有关。
本研究中,连作淮山根际土壤细菌群落的优势菌门有放线菌门、变形菌门、厚壁菌门、绿弯菌门、酸杆菌门和拟杆菌门等,与张红霞等[13]对山药的研究结果相似。其中,变形菌门、拟杆菌门、粘球菌门、Patescibacteria和疣微菌门等5门的相对丰度显著大于CK;连作淮山和CK根际土壤细菌群落的优势菌属(相对丰度>1.00%)共有37个,但仅芽孢杆菌属、慢生根瘤菌属、norank_JG30-KF-CM45、热酸菌属和Bryobacter等5属为二者排名前20位优势菌属中的共有菌属,表明在细菌属水平上连作淮山与CK的优势菌属差异较明显;连作淮山根际土壤细菌群落中出现链霉菌属和鞘氨醇单胞菌属等有益菌优势菌属,可能与淮山连作过程中施用有机肥有关。本研究仅为2年的试验数据,为更明确淮山连作后其土壤菌属的变化规律,还需结合淮山连作后病害发生状况及淮山产量和品质等性状等进行跟踪研究,才能最终筛选出引起淮山连作障碍的致病菌株,为解决淮山连作障碍提供精确的科学依据。
本研究结果表明,在连作淮山根际土壤细菌群落的12个主要细菌门中,有7个细菌门与土壤pH及水解氮、速效钾和有机质含量等理化性质呈显著或极显著相关,表明土壤理化性状改变是引起微生物群落组成发生变化的原因之一,与秦红灵等[25]和赵帆等[26]报道的结果一致。
4 结 论
定向栽培连作淮山可导致其根际土壤细菌多样性和丰富度发生变化,增加大量特有细菌,其根际土壤细菌的Shannon指数、ACE指数和Chao l指数均显著上升,优势细菌群落主要为放线菌、变形菌、厚壁菌、绿弯菌和酸杆菌。土壤pH及水解氮、速效钾和有机质含量变化是影响连作淮山土壤细菌群落组成发生变化的重要因子。
猜你喜欢
安徽农业科学(2022年20期)2022-11-11
科学技术与工程(2022年26期)2022-11-01
今日农业(2022年12期)2022-07-06
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
中国生态农业学报(中英文)(2021年9期)2021-09-10
中国果业信息(2020年10期)2020-12-15
第二课堂(小学版)(2019年7期)2019-07-16
