基于热溶解膜过滤法的太岁古菌菌群结构研究
2021-09-08 02:45:18王朝江王世清信艳杰高春燕
生命科学研究 2021年4期
王朝江,王世清,李 慧,信艳杰,高春燕
(河北省农林科学院遗传生理研究所,中国河北 石家庄 050051)
太岁是一种在中国古籍有记载[1],现代不断发现[2],科学归属正在探索的神秘物体[3~4]。1992年发现的现代首例太岁被鉴定为“以黏菌为主体的特大型黏菌复合体”[5],但该结论又于2010年被作者自我否定[6]。于是,研究者不得不转向探索太岁中存在的其他菌类作为主体菌的可能性。
采用培养方法,研究者在太岁中分离出了丝状真菌、酵母、细菌[7~9];采用非培养方法,特别是高通量测序技术,研究人员获知了太岁细菌和古菌的菌群结构,不仅证明太岁中细菌种群多样性丰富[10],还发现有11个属的古菌存在[11],但在不同太岁间检测到的细菌和古菌的优势属差别巨大。细菌中检出的优势属有Edaphobacter、梭菌属(Clostridium)、芽孢杆菌属(Bacillus)[10]、盐单胞菌属(Halomonas)[12],其间没有相同的属,即使在两个形态近似、产地相同的太岁间也是如此。古菌有相同的优势属,分别是甲烷杆菌属(Methanobacterium)、甲烷短杆菌属(Methanobrevibacter)和甲烷球菌属(Methanosphaera),但它们在不同太岁间的相对丰度差别很大。这种差异化的结果不能在不同形态太岁间建立起以共有菌群为纽带的联系,也不能为分离太岁的主体菌准确提供目标信息。
高通量菌群测定结果的差别,无疑由所用DNA间的差别决定,而DNA又受其提取方法的影响,对此,已不乏研究报道[13]。本文作者用热溶解膜过滤方法提取的DNA,重测同一太岁的细菌菌群结构,结果证明:之前采用的常规材料处理方法,未考虑太岁质地如胶、不溶于冷水并含有大量多糖类物质(包括聚乙烯醇)的特性,所提取的DNA包含的细菌信息片面,导致测知的菌群结构严重失真,真正优势菌群的相对丰度降低或漏检,非优势菌群则以优势菌群面目出现;同时,本文作者还通过设置不同取样位置和重复的方法,从极富多样性的细菌群落中确定出了优势菌群[14]。目前,尚未有该方法在太岁古菌菌群结构研究中应用的报道。
古菌通常存在于高温、高盐、缺氧、强酸、强碱等极端环境,分离培养的条件苛刻,且生长慢不易观察[15]。因此,古菌优势菌的分离较细菌更加依赖于高通量测序技术,只有通过多次测定,特别是建立在适合太岁材料特性的DNA提取方法基础上的测定,才能获知太岁准确的古菌优势菌群信息,同时也可进一步证实热溶解膜过滤方法对已有结果偏差的修正效果。
1 材料与方法
1.1 供试材料
太岁样本发现于河南省三门峡市黄河流域,呈红褐色和尖圆柱体状,质感如低弹性橡胶,2007年由笔者购回收藏。
1.2 试剂和仪器
十六烷基三甲基溴化铵(hexadecyl trimethyl ammonium bromide,CTAB)(分析纯,天津市大茂化学试剂厂);2×Taq Master Mix(北京全式金生物技术有限公司);DNA检测试剂盒(Qubit 2.0,美国Life公司)。PCR仪(T100,美国Bio-Rad公司);MiSeq高通量测序平台(2×300,美国Illumina公司)。
1.3 方法
1.3.1 取样与DNA提取
在太岁的表里层不同部位做两次重复取样。表层取样部位位于表皮,深约0.8 cm,取样前用无菌水和滤纸对外表做清洁处理,样品编号为SAO2和SAO3;里层取样部位位于表皮下2~3 cm处,样品分别编号为SAI2和SAI3。每份样品湿重0.20 g,切碎后加水水浴溶化,用针头滤器过滤溶化液,即热溶解膜过滤法[16],之后切取滤膜用作DNA提取。DNA经1%琼脂糖凝胶电泳检验后,送生工生物工程(上海)有限公司进行测序。
1.3.2 高通量测序
采用DNA检测试剂盒测定DNA含量后,使用融合有MiSeq平台接头序列和标签(barcode)序列的古菌特异性引物349F和806R,扩增16S rDNA的V3~V4区。扩增体系和程序见文献[11]。回收4个样品DNA的扩增产物,用DNA检测试剂盒精确定量,各取10 ng混合后测序。
1.3.3 测序数据处理与深度验证
用Cutadapt软件去除测序原始数据的接头,用Pear软件[17]完成序列拼接,然后按照barcode识别得到各样本的原始序列(raw reads)。原始序列经Prinseq软件[18]质控,去除嵌合体、非扩增区域序列、低于阈值的BLASTn比对序列[19],得到优质序列(seq reads)。使用Usearch(version 5.2.236)软件[20]在97%的相似度水平下将优质序列聚类为不同的操作分类单元(operational taxonomic unit,OTU)。
利用覆盖率(Coverage)和香农(Shannon)指数稀疏曲线评价测序数据量的合理性。覆盖率计算公式:C=1-n1/N,式中:n1为只含有1条序列的OTU数目,N为序列总数。
1.3.4 古菌多样性及OTU分布分析
采用α多样性评价古菌的多样性及丰度。其中,香农指数、辛普森(Simpson)指数反映群落物种多样性,前者数值越高,或后者数值越低,则多样性越高;ACE指数、Chao1指数反映物种丰度,值越大,则丰富度越高。
使用R的韦恩(Venn)图软件统计样品中共有的和独有的OTU数目,分析其在样品间的重叠情况[14,16]。
1.3.5 古菌物种分类及优势菌群构成分析
选择OTU中丰度最高的序列作为代表性序列,用 Naïve Bayesian assignment算法,对其进行门、纲、目、科、属等不同水平的分类[21]。相对丰度是各个类群的序列数占总序列数的百分比[22]。相对丰度高且样品间共有的类群被初步确定为优势菌群。为准确反映物种间的相对丰度差别,该类数值保留4位小数。
1.3.6 OTU对优势属的支撑分析
分析优势属中OTU的数目构成及其相对丰度,并根据不同样品间是否存在共有且占比高的OTU,判定优势属是否是太岁中古菌的真正优势菌群[14,16]。
2 结果与分析
2.1 样品DNA的提取和扩增
SAO2、SAO3、SAI2、SAI3 等4 个样品中提取的DNA 的质量浓度分别是 7.20 ng/μL、10.36 ng/μL、0.27 ng/μL、2.20 ng/μL,均高于高通量测序需要的最低质量浓度(0.05 ng/μL)。样品DNA及其扩增产物的电泳结果见图1。
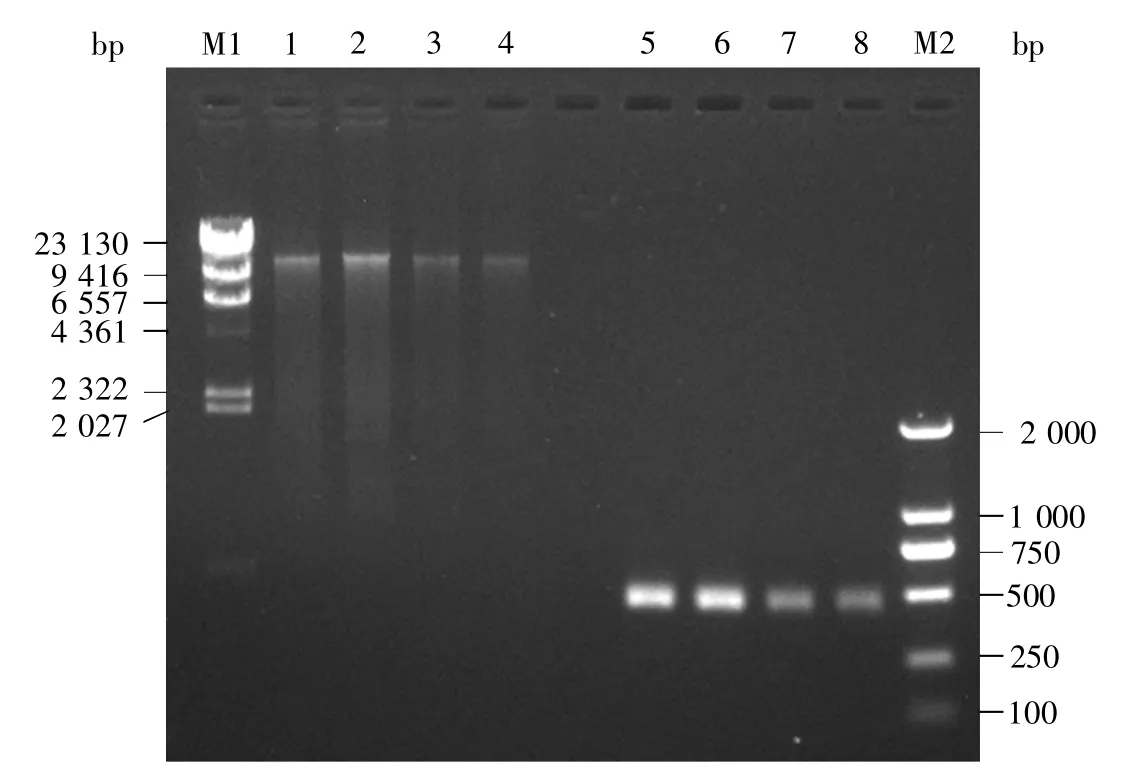
图1 4个样品中提取的DNA及其扩增产物电泳图M1:λDNA/HindⅢ单切DNA分子量标准;M2:DNA分子量标准 DL2000;1~4:SAO2、SAO3、SAI2、SAI3样品中提取的DNA;5~8:SAO2、SAO3、SAI2、SAI3 的 DNA 扩增产物。Fig.1 Electrophoresis of sample DNAs and amplification productsM1:λDNA/Hind Ⅲ DNA marker;M2:DL2000 DNA marker;1~4:Extracted DNAs from four samples;5~8:DNA amplification products of four samples.
2.2 测序数据及深度验证
高通量测序后获得的4个样品的原始序列、质控序列(clean reads)和优质序列的数目,以及优质序列聚类为OTU的数目与文库覆盖率见表1。从表1可以看出,里层样品所含OTU数目明显低于表层样品。如此,若假定太岁为表层生长,可推知其由表至里存在种群消失现象;若为里层生长,则外部的多数菌对太岁生长非必需,为环境黏附冗余。覆盖率指数均接近1,表明样本中所有的序列基本被测出,测序结果能够代表样本的真实情况。

表1 4个样品的序列及OTU数目信息Table 1 Sequence and OTU numbers of 4 samples
另外,4个样品的香农稀疏曲线趋于平稳,这也说明测序数据量合理,更多的数据量只会产生少量的新物种(图2)。
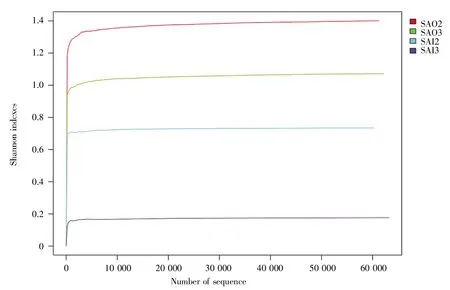
图2 4个样品的香农指数稀疏曲线Fig.2 The Shannon rarefaction plot of 4 samples
2.3 古菌α多样性及OTU分布
4个样品的α多样性指数见表2。从表中可以看出,多样性指数在样品间的变化幅度很大,说明各样品在物种多样性和丰度上存在巨大差异,多个重复样品对获得正确菌群结构是必需的。此外,SAO2和SAO3的香农指数、ACE指数、Chao1指数均高于SAI2和SAI3,SAI3的香农指数最低,说明表层样品的物种多样性远高于里层样品,同时也说明太岁里层的古菌种类单一,且分布极不均衡。
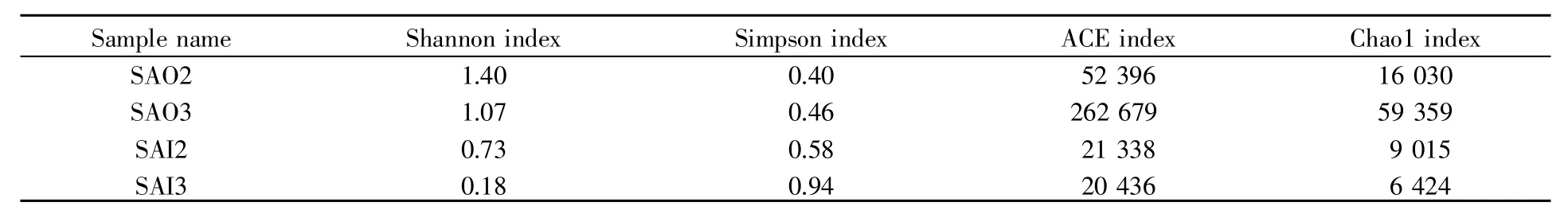
表2 4个样品的α多样性指数Table 2 Alpha diversity indexes of 4 samples
OTU在样本间的重叠和独有情况见图3。从图中可以直观看出,4个样品共有OTU只占每个样品OTU的很少一部分,说明其共有种类少且分布极不均衡。通过核对相对丰度数据,我们发现共有的 3 个 OTU(OTU2、OTU63、OTU107)存在超高占比,其占比之和在SAO2、SAO3、SAI2和SAI3样品中依次为89.950 3%、94.561 7%、71.842 6%、99.588 5%,特别是OTU2,在SAI3中独占96.811 0%,提示供试太岁中虽然菌群种类多但优势种类突出。
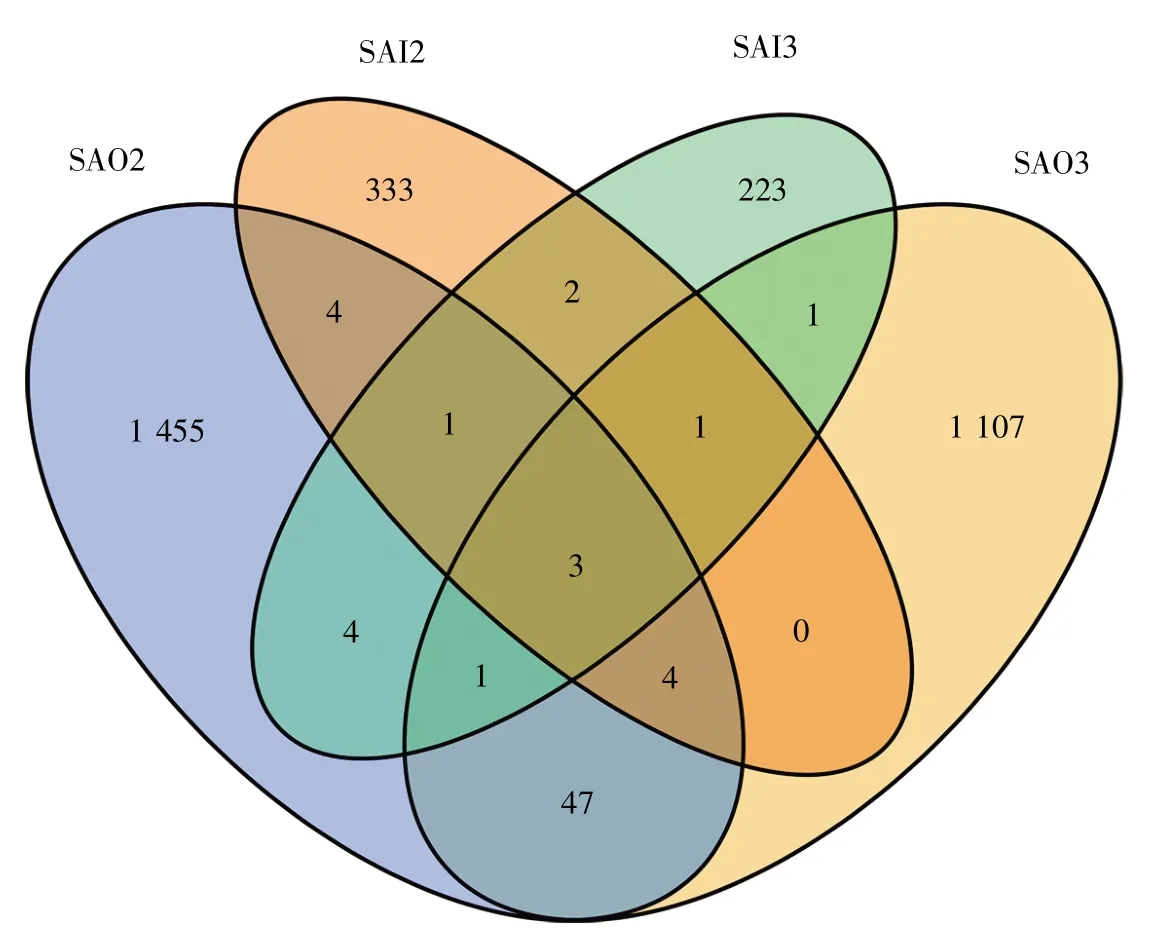
图3 4个样品的OTU分布韦恩图Fig.3 The OTU Venn diagram of 4 samples
2.4 太岁古菌的物种分类及优势类群解析
4个样品在各分类水平上所含分类单元总数和百分占比≥0.01%的数目情况见表3。从表3中可以看出,样品在各分类水平上的古菌总数,在表里层位置间是表层多于内部,重复间亦有差异;百分占比≥0.01%的古菌数目在各样品间的差别,远小于总数间的差别,说明供试太岁中有大量极低相对丰度的古菌菌群存在。

表3 4个样品在门、纲、目、属分类水平上的古菌数目分析Table 3 The archaea number in 4 samples at different taxonomy levels
门分类水平上,共检出16个门。低于0.01%占比的门数在样品间相差1~7个,差别最大的是SAO2样品。SAO2和SAO3共检出16个门,SAO2中全含,SAO3中仅有8个,存在差异的8个门在SAO2中的占比为0.073 5%,二者共有的8个门在SAO2和SAO3中的占比分别为99.926 5%和100.000 0%。SAI2和SAI3共检出9个门,表面相差2个,但由于SAI2独有4个,SAI3独有2个,所以两个样品实际相差6个门;二者共有的3个门在SAI2和SAI3中的占比分别为99.951 9%和99.993 7%。
相对丰度大于1%的门共检出4个,分别是广古菌门(Euryarchaeota)、奇古菌门(Thaumarchaeota)、疣微菌门(Verrucomicrobia)和乌斯古菌门(Woesearchaeota)。SAO2、SAO3、SAI2 和 SAI3 所涉及的数目分别为4、3、2、2。广古菌门和奇古菌门为共有门,它们在 SAO2、SAO3、SAI2和 SAI3中的占比之和依次为95.216 9%、97.684 0%、99.935 3%、99.971 5%。广古菌门在表层中的占比基本与奇古菌门持平,但在里层SAI2和SAI3中的占比分别为72.229 1%、97.181 3%,优势突出。以上信息说明,样品间共有门的占比具绝对优势,且里层更加突出。
纲水平上共检出27个纲,低于0.01%占比的纲在SAO2中最多,占总数的一半。SAO2和SAO3共检出27个纲,相差17个,均为SAO2独有,占比之和为0.205 8%;二者共有的10个纲在各自的占比分别为99.794 2%和100.000 0%。SAI2和SAI3共检出10个纲,表面相差4个纲,但由于SAI2独有6个,SAI3独有2个,所以两个样品实际相差8个纲;二者共有的4个纲在各自的占比分别为99.927 1%和99.993 7%。
相对丰度大于1%的纲共检出5个,分别是甲烷杆菌纲(Methanobacteria)、甲烷微菌纲(Methanomicrobia)、奇古菌门未分类纲、未分类纲(unclassified)和乌斯古菌门未分类纲。SAO2、SAO3、SAI2和SAI3所涉及的数目分别为5、3、3、2,其中,甲烷杆菌纲和奇古菌门未分类纲为共有纲,二者占比之和依次为93.585 0%、97.315 4%、98.500 3%、99.949 4%。另外,甲烷杆菌纲在SAI3中的占比高达97.159 1%,同样是内部占比高于外部。
目分类水平上,共检出39个目。低于0.01%占比的目在SAO2样品中最多,为24个。SAO2和SAO3共检出39个目,表面相差25个目,但由于SAO2独有26个,SAO3独有1个,所以两个样品实际相差27个目;二者共有的12个目在各自的占比分别为99.790 9%和99.995 2%。SAI2和SAI3共检出15个目,表面相差7个目,但由于SAI2独有8个,SAI3独有1个,所以两个样品实际相差9个目;二者共有的6个目在各自的占比分别为99.877 2%和99.996 8%。
相对丰度大于1%的目有6个,分别是甲烷杆菌目(Methanobacteriales)、甲烷八叠球菌目(Methanosarcinales)、Nitrosopumilales、Nitrososphaerales、未分类目(unclassified)和乌斯古菌纲未分类目。SAO2、SAO3、SAI2 和 SAI3 所包含的数目分别为 5、3、3、2,其中,甲烷杆菌目和Nitrososphaerales为共有目,二者占比之和依次为91.835 5%、96.346 6%、98.490 4%、99.949 4%。SAO2样品中共有目占比较低的原因是其单独含有占比之和为5.668 5%的乌斯古菌纲未分类目等3个目。
属分类水平上,共检出64个属。低于0.01%占比的属在SAO3和SAI2样品中占近半,在SAO2中占2/3。SAO2和SAO3共检出62个属,表面相差43个属,但由于SAO2独有44个,SAO3独有1个,所以两个样品实际相差45个属;二者共有的17个属在各自的占比分别为99.683 1%和99.998 4%。SAI2和SAI3共检出19个属,表面相差13个属,但因SAI2独有14个,SAI3独有1个,所以两个样品实际相差15个属;二者共有的4个属在各自的占比分别为99.865 6%和99.998 4%。以上数据说明表层和里层中非共有属的占比极低。
相对丰度大于1%的属共检出5个,分别是甲烷杆菌属、甲烷丝菌属(Methanothrix)、Nitrosopumilus、亚硝化球菌属(Nitrososphaera)和未分类属(unclassified),SAO2、SAO3、SAI2 和 SAI3 所涉及的数目分别为4、3、3、2。5个属占比之和最低的仍有98.279 9%,其中,甲烷杆菌属和亚硝化球菌属为共有属,二者在SAO2、SAO3、SAI2和SAI3中的占比之和依次为91.654 1%、96.312 8%、98.480 4%、99.944 6%,占绝对优势。此外,甲烷杆菌属和亚硝化球菌属的占比在表里层不均衡。前者在SAO2、SAO3、SAI2、SAI3 样品中的占比分别为44.504 7%、55.526 1%、70.784 2%、97.154 4%,里层明显高于表层;后者则是表层高于里层(图4)。
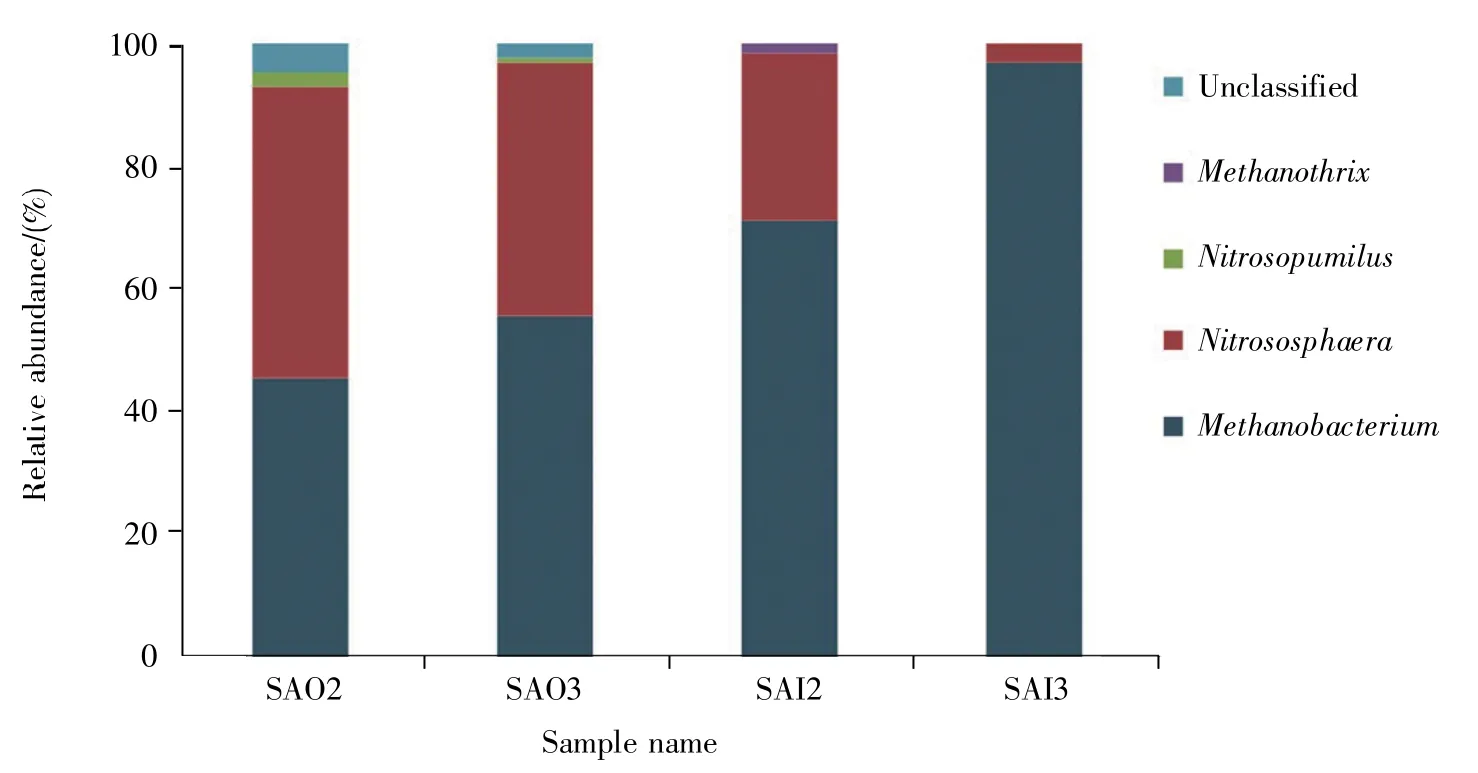
图4 4个样品中相对丰度大于1%的属结构分布图Fig.4 The column chart of 5 dominant genera with relative abundance higher than 1%
2.5 OTU对优势属的支撑能力
4个样品中甲烷杆菌属和亚硝化球菌属所包含的OTU数目、相对丰度位于前两位的OTU及其占比情况见表4。从表中可以看出,两个属均含有众多OTU,其数目均是表层多于里层;在每个属中第一、二位OTU的相对丰度差别极大,第一位OTU的占比近乎等于其所在属的占比;OTU2在4个样品的甲烷杆菌属中的占比均位列第一,同时也是4个样品的共有OTU;在亚硝化球菌属中,占比高的OTU63分布在表层,内部极少且不共有。上述结果说明,仅甲烷杆菌属是有高丰度OTU支撑的真正优势属,其中优势种明显,可代表太岁所含古菌的优势菌群。
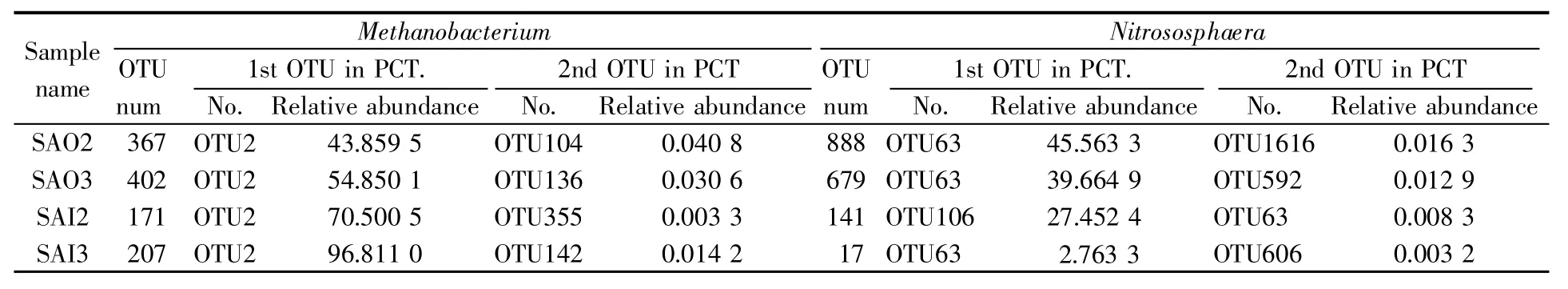
表4 甲烷杆菌属和亚硝化球菌属的OTU分析Table 4 OTU analysis of Methanobacterium and Nitrososphaera in 4 samples
3 结论
供试太岁所含古菌分属16个门、27个纲、39个目、64个属。优势菌群是甲烷杆菌属,相对丰度在43.859 5%~96.811 0%。
太岁所含古菌的多样性是表层高于里层,而优势菌群的相对丰度是里层显著高于表层。
各样品的菌群在种类和数量上均存在差别,有大量非共有菌群,其可判定为机会性进入太岁组织中的杂菌。
4 讨论
采用热溶解膜过滤法提取DNA所测得的种群结构,较之前的测定结果[11]变化巨大。大于1%的属,本次内部样品测得4个,而之前的内部样品测得10个,其中仅甲烷杆菌属相同,但本次所测的丰度大幅升高。甲烷杆菌属占比在SAI2和SAI3样品中分别为70.784 2%、97.154 4%,比之前结果分别上升39.554 2%和65.924 4%。对于之前测定的占比最高和第三的两个属——甲烷球菌属(34.15%)与甲烷短杆菌属(17.25%),前者在本次仅测得1条序列(read),占比0.001 6%,后者却未检测到。此外,和之前结果相比,本次测定的丰度次高的亚硝化球菌属的占比也大幅上升,其在SAI2和SAI3样品中的占比分别为27.696 2%和2.790 2%,而之前的结果则为0.28%[11]。以上数据说明,采用热溶解膜过滤法提取的DNA,含有太岁中古菌主体菌群的更多信息。主体菌群信息的增加,相对挤占和压减了非优势菌群的占比,凸显了正确的菌群结构。另外,本次研究中设置重复样品和着重分析其间共有菌群的方法,非常有助于排除机会性菌的干扰,从而解析出真正的优势菌群。
甲烷杆菌属作为两次测定的古菌优势菌群,结果可信度高,其在形态和代谢特性上均具有助于太岁形成的可能性。形态上,甲烷杆菌属菌体弯曲、钩形到长杆状,可以形成长的丝状体到类球状体,具备结团能力[23~25];代谢上,其能产生很多胞外聚合物,电镜下可观察到该聚合物从一个细胞的外表面延伸黏附至另一细胞,具备使菌体团块不易散开的物质基础[26],可以形成絮状沉淀[24,27]。另外,剖开刚采自池塘污泥的其他柱状太岁,能观察到类似气腔和呈膜片状的结构(图5),说明太岁中有厌氧产气的菌类存在。

图5 一柱状太岁切面出现的气腔和膜片结构Fig.5 Air cavities and membranes in the section of a columnar Taisui
甲烷杆菌属和亚硝化球菌属在太岁中并存,且丰度占比在表里层样品中呈相反的趋势,这至少反映二者对氧气的生理需求有差异。资料显示,甲烷杆菌属是严格厌氧的,亚硝化球菌属在低氧环境下则具有一定耐受能力[28~29],这可使表层组织中亚硝化球菌属占比较高获得合理解释,同时也提示从太岁的表层到内部,氧气浓度呈现出从低氧到无氧的变化。另外,亚硝化球菌还是无泡曝气生物反应器生物膜的组成成分之一[29],而生物膜是一种菌胶团,一定程度上具有太岁状物质的特征。
猜你喜欢
课堂内外·小学版(低年级)(2023年5期)2023-04-29 16:01:41
河南医学研究(2022年19期)2022-10-19 00:44:18
土壤学报(2022年3期)2022-08-26 12:15:26
大自然探索(2022年5期)2022-07-11 03:10:33
知识就是力量(2022年6期)2022-06-16 20:19:36
生态学报(2019年11期)2019-07-08 06:18:58
文史博览·文史(2017年2期)2017-03-01 18:10:33
应用海洋学学报(2014年4期)2014-11-22 07:43:56
食品工业科技(2014年23期)2014-03-11 18:19:08
现代检验医学杂志(2014年6期)2014-02-02 03:01:54