
杂交兰‘紫妍氏’叶艺性状的生理生化与细胞结构分析
2021-09-04 03:16:22钟淮钦林榕燕吴建设叶秀仙
西北植物学报 2021年7期
钟淮钦,林榕燕,吴建设,林 兵,叶秀仙
(福建省农业科学院作物研究所/福建省特色花卉工程技术研究中心,福州 350013)
杂交兰(Cymbidiumhybrid)是国兰(ChineseCymbidium)和大花蕙兰(Cymbidiumhybridum)种间杂交培育而成的一类兰花的特称,兼具国兰的植株小巧、幽香、典雅和大花蕙兰的花大、色艳、花期长等优异特性,观赏和经济价值高,极具发展潜力[1-2]。杂交兰在长期的人工选育培养过程中,获得一批稳定的叶色变异新种质,即叶片上不同位置镶嵌着或金黄、或银白、或朱红的条纹或斑块,称之为线艺或叶艺;叶艺品种观赏和商品价值成倍增加,成为了育种的重要目标[3-4]。鉴于杂交兰叶艺性状对其商品性的重要作用,研究其形成机理,对人工诱发叶艺具有重要意义。
叶色突变通常是由于叶绿素的代谢过程直接或间接受到突变基因的影响,从而引起叶绿素的含量和比例发生变化[5-7]。叶色突变体不仅是遗传和育种研究中的重要材料,还是研究光合机制、叶绿素代谢、激素生理等的理想材料[8],同时在观赏植物中能够极大地提高观赏价值。国内外学者已在拟南芥[9]、烟草[10]、水稻[11]、大豆[12]、玉米[13]和番茄[14]等模式植物和大宗作物中获得、鉴定出不同类型的叶色突变体,对其结构、功能、作用等进行了大量研究。而在兰属[15-18]、文心兰[19]、铁皮石斛[20]、菊花[21]和红掌[22]等观赏植物中虽然发现了多种类型的叶色突变体,但对其变异机理研究的相关报道较少,研究结果也不尽相同。目前尚未见有关杂交兰叶色突变体的研究报道。
本研究以杂交兰品种‘紫妍氏’(绿叶)及突变选育的3个稳定叶色变异(叶艺)新品系‘福韵西施’、‘爪艺紫妍氏’和‘中透紫妍氏’为材料,从生理学和细胞学角度分析叶片绿色区域与黄色、银色区域的光合色素、叶绿素合成前体物质、叶绿素合成相关酶活性、叶绿素荧光特性及叶片显微结构、叶绿体超微结构的差异,旨在探讨杂交兰叶色变异的生理基础,为进一步揭示其形成的分子机理奠定基础,也为合理利用这一新种质提供参考依据。
1 材料和方法
1.1 材 料
试验材料为福建省农业科学院作物所花卉研究室种质资源圃保存的长势基本一致且生长状况良好的杂交兰‘紫妍氏’(K21)及其3个叶艺品系‘福韵西施’(K21-1)、‘爪艺紫妍氏’(K21-2)和‘中透紫妍氏’(K21-3)。其中,‘紫妍氏’叶片绿色,‘福韵西施’叶片绿色,叶尖至叶片中部边沿具黄色条纹(爪艺),‘爪艺紫妍氏’叶片绿色,叶尖至叶片中部边沿具银色条纹(爪艺),‘中透紫妍氏’叶片中间具黄白色宽条纹,叶尖至叶片中部边沿为绿色。
试验于2019年9月在福建省特色花卉工程技术研究中心花卉生理实验室进行,每组样品设置3个重复,具体用量见测定指标与方法。文中用编号和字母表示各品系叶片取样测定部位,即‘紫妍氏’绿色叶片为K21-G;‘福韵西施’叶片绿色部位为K21-1G,黄色部位为K21-1Y;‘爪艺紫妍氏’叶片绿色部位为K21-2G,银色部位为K21-2Y;‘中透紫妍氏’叶片绿色部位为K21-3G,黄白部位为K21-3Y。
1.2 测定指标与方法
1.2.1 光合色素含量测定参考彭运生等[23]、王学奎[24]的方法,并做适当修改。用ddH2O清洗叶片样品,晾干后,分别剪取4个供试材料叶片中的绿色、黄色或银色部位,剪碎后称取0.1 g置于20 mL离心管中,加入10 mL萃取液(丙酮∶无水乙醇=2∶1),黑暗放置24 h直至叶片完全失绿。用紫外分光光度计分别在470 nm、645 nm和663 nm波长下测定浸提液的吸光度A470、A645和A663,共3次重复,最后依据以下公式计算各光合色素含量:
叶绿素a 含量(Chl a)=(12.21A663-2.81A645)V/(1 000W);
叶绿素b 含量(Chl b)=(20.13A645-5.03A663)V/(1 000W);
类胡萝卜素含量(Car)=(1 000A470+483.193 3A663-2 084.331 3A645)V/(229 000W);
式中,V为提取液体积,W为样品鲜重。
1.2.2 叶绿素合成前体物质含量参照韩巧红[25]的方法,略作修改,对供试材料的δ-氨基乙酰丙酸(ALA)和胆色素原(PBG)含量进行测定;参照毛晶晶等[26]的方法,略作修改,对供试材料的尿卟啉原 Ⅲ(Urogen Ⅲ)含量和粪卟啉原 Ⅲ(Coprogen Ⅲ)含量进行测定;参照田韦韦[27]的方法,对供试材料的原卟啉Ⅸ(ProtoⅨ)、镁原卟啉Ⅸ(Mg-Proto Ⅸ)和原叶绿素酸酯(Pchlide)含量进行测定。将‘紫妍氏’绿色叶片的7种叶绿素合成前体物质含量设为100%,叶艺品系叶绿素合成前体物质含量以相对于‘紫妍氏’的百分比表示。
1.2.3 叶绿素合成途径相关酶活性称取一定量的样品材料,以1∶9的比例加入浓度为0.01 mol/L 的PBS缓冲液(pH 7.4),迅速将样品匀浆化,2 500 r/min离心20 min后,收集上清夜,用离心管分装后置于-20 ℃保存备用。氨基乙酰脱氢酶(ALAD)、胆色素原脱氨酶(PBGD)、尿卟啉原Ⅲ合成酶(UROS)、尿卟啉原Ⅲ脱羧酶(UROD)、粪卟啉原Ⅲ氧化酶(CPOX)和原卟啉原氧化酶(PPOX)活性的测定使用对应的测定试剂盒(均购置于上海晶抗生物工程有限公司),操作步骤参照各试剂盒说明书。
1.2.4 叶绿素荧光参数参照GFS-3000光合仪(德国WALZ公司)使用说明书,选择在上午9:00-11:00于室内测定4个供试材料叶片绿色、黄色或银色部位的叶绿素荧光参数PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率Y(Ⅱ)、光化学淬灭系数(qP)和非化学淬灭系数(NPQ)。其中Fv/Fm值是在叶片经20 min暗处理后于室温下测定,其余参数值则直接在室温下测定,共3次重复。
1.2.5 叶片显微结构参照翁伯琦等[28]的方法,对4个供试材料叶片绿色、黄色或银色部位的显微结构进行观察。取样后,经固定、漂洗、脱水、干燥、喷涂等步骤,在扫描电子显微镜(日本电子株式会社)下观察,并采集图像进行分析。
1.2.6 叶绿体超微结构取4个供试材料叶片绿色、黄色或银色部位,经电镜固定液4 ℃固定,离心,电镜固定液4 ℃再固定,PBS缓冲液(pH 7.4)漂洗,锇酸-磷酸缓冲液PBS(pH 7.4) 20 ℃固定,PBS缓冲液(pH 7.4)漂洗等步骤完成材料的固定程序。而后将材料脱水、渗透、包埋、切片、染色,最后再用透射电子显微镜(美国FEI公司)观察并采集图像进行分析。
1.3 数据分析
采用Excel 2007软件对文中涉及的数据进行统计和分析,利用SPSS 20.0软件中的Duncan’s multiple range test方法对统计后的数据进行差异显著性分析。
2 结果与分析
2.1 杂交兰‘紫妍氏’及其叶艺品系叶片光合色素含量比较
由表1可见,3个叶艺品系叶片绿色区域(K21-1G、K21-2G和K21-3G)的光合色素含量均不同程度地低于‘紫妍氏’绿叶(K21-G);三者的类胡萝卜素(Car)含量均与K21-G差异不显著(P>0.05),而它们的叶绿素b(Chl b)和总叶绿素(Chl)含量以及K21-2G和K21-3G的叶绿素a(Chl a)含量降幅均达到显著水平(P<0.05); 3个叶艺品系叶片绿色区域的Car/Chl值均与K21-G差异不显著,而其Chla/b值均明显高于K21-G,且K21-3G的增幅还达到显著水平。说明杂交兰叶艺品系叶片绿色区域的叶绿素含量比‘紫妍氏’显著降低,且以叶绿素b降低幅度更大,而其类胡萝卜素含量则无显著变化。
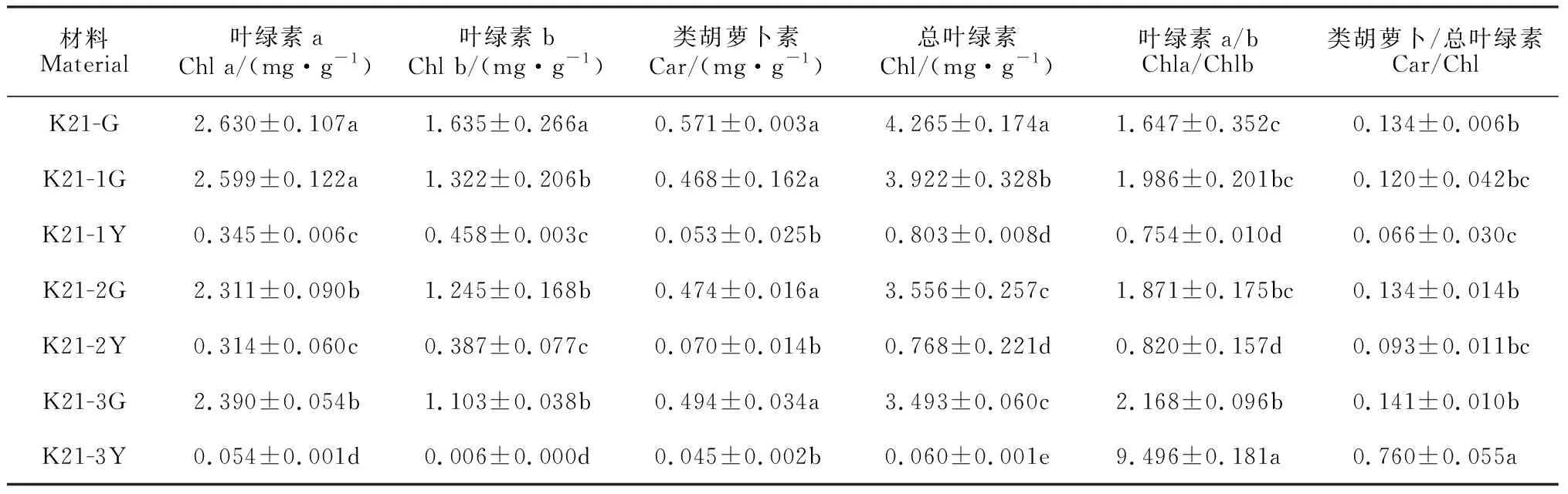
表1 ‘紫妍氏’不同品系叶片光合色素含量及比值
同时,3个叶艺品系叶片叶艺区域(K21-1Y、K21-2Y和K21-3Y)的光合色素含量均显著低于其相应的绿色区域,三者的Chl含量降幅分别达到79.5%、78.4%和98.3%,Car含量降幅分别达到88.7%、85.2%和90.9%(P<0.05)。与叶艺品系相应绿色区域相比较,Car/Chl值在K21-3Y中显著增加了4.39倍(P<0.05),而在K21-1Y、K21-2Y中分别减少了45.0%和30.6%(P>0.05),说明叶艺品系叶艺区域光合色素含量和比例的变化导致植株叶色黄化。进一步分析发现,Chla/b值在K21-1Y、K21-2Y中也显著低于其相应的绿色区域,说明‘福韵西施’和‘爪艺紫妍氏’叶艺区域叶绿素a含量降低程度明显高于叶绿素b含量,推测叶绿素a缺乏是其叶片变异的主因; Chla/b值在K21-3Y中显著高于其相应的绿色区域,说明‘中透紫妍氏’叶艺区域叶绿素b含量降低程度明显高于叶绿素a,推测叶绿素b缺乏是其叶片黄化的主因。
2.2 杂交兰‘紫妍氏’及其叶艺品系叶片叶绿素合成前体物质相对含量比较
叶绿素生物合成代谢过程中的某一步发生障碍,就会导致该反应的前体物质积累,其后的物质积累降低。因此,通过比较突变品系和亲本叶绿素生物合成过程中的一些中间产物量,即主要前体物质的累积量,可以来初步判断发生障碍部位所在。本研究测定了3个叶艺新品系和亲本叶片叶艺区域与绿色区域叶绿素生物合成前体物质δ-氨基乙酰丙酸(ALA)、胆色素原(PBG)、尿卟啉原 Ⅲ(Urogen Ⅲ)、粪卟啉原 Ⅲ(Coprogen Ⅲ)、原卟啉Ⅸ(ProtoⅨ)、镁原卟啉Ⅸ(Mg-Proto Ⅸ)和原叶绿素酸酯(Pchlide)的含量。将‘紫妍氏’绿叶7种叶绿素合成前体物质含量均设为100%,叶艺品系材料相应叶绿素合成前体物质含量以相对于‘紫妍氏’绿叶的百分比表示。
图1显示,与‘紫妍氏’绿叶(K21-G)相比,‘福韵西施’和‘爪艺紫妍氏’叶片绿色区域(K21-1G和K21-2G)的ALA、PBG、Urogen Ⅲ相对含量大多显著提高,其Coprogen Ⅲ、ProtoⅨ及Mg-Proto Ⅸ相对含量均变化不显著(P>0.05),而它们的Pchlide相对含量则显著降低(P<0.05);‘中透紫妍氏’叶片绿色区域(K21-3G)除了Pchlide相对含量比K21-G显著降低(P<0.05)外,其余6个前体物质相对含量均与K21-G差异不显著(P>0.05)。
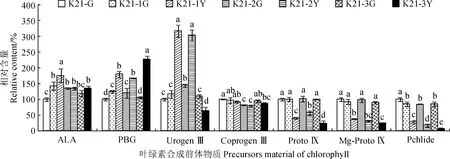
ALA. δ-氨基乙酰丙酸;PBG. 胆色素原; Urogen Ⅲ. 尿卟啉原 Ⅲ;Coprogen Ⅲ. 粪卟啉原 Ⅲ;ProtoⅨ. 原卟啉Ⅸ;Mg-Proto Ⅸ. 镁原卟啉Ⅸ;Pchlide. 原叶绿素酸酯图1 ‘紫妍氏’不同叶艺品系叶片叶绿素合成前体物质相对含量ALA. δ-aminolevulinic acid; PBG. Porphobilinogen; Urogen Ⅲ. Uroporphyrinogen Ⅲ; Coprogen Ⅲ. Coproporphyrinogen Ⅲ;ProtoⅨ. Protoporphyrin Ⅸ; Mg-Proto Ⅸ. Mg-protoporphyrin Ⅸ; Pchlide. ProtochlorophyllideFig.1 Relative contents of precursor materials of chlorophyll in leaves of different ‘Purple Element’ strains
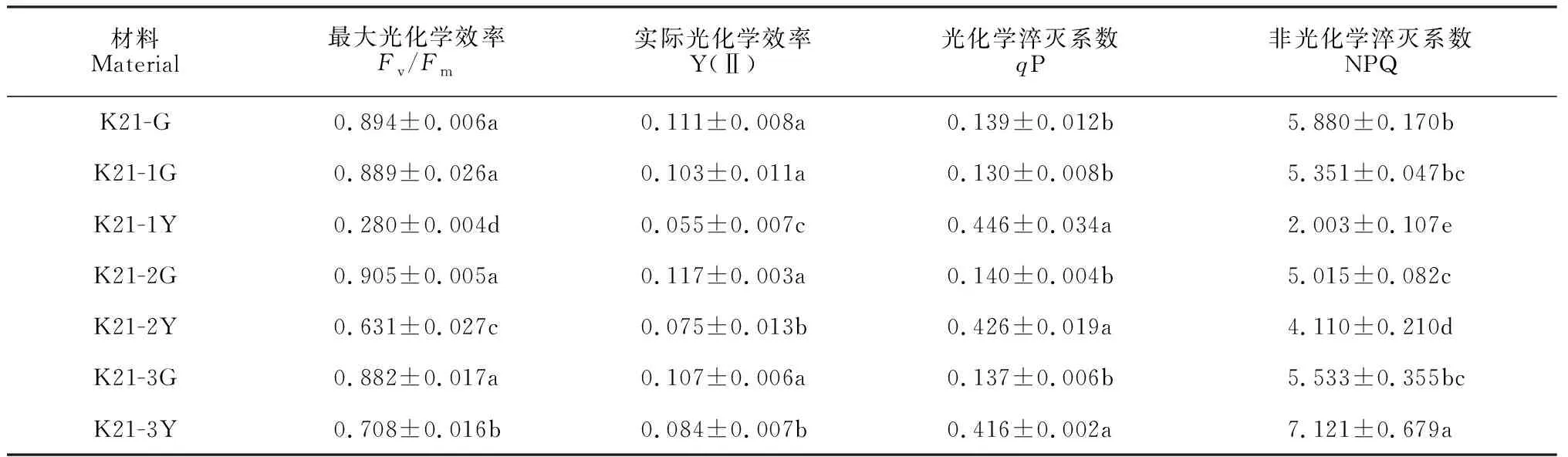
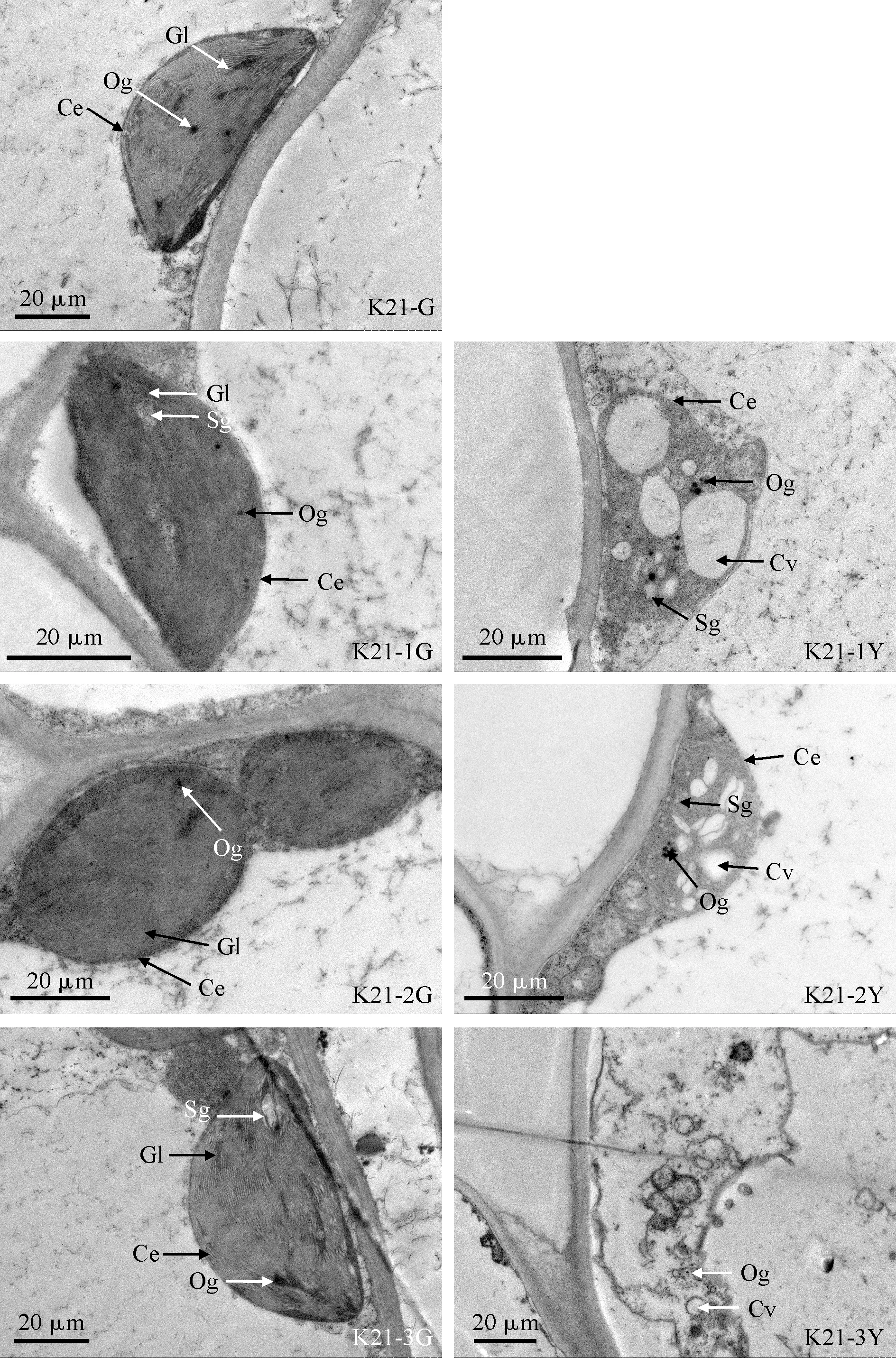
同时,在ALA →PBG→Urogen Ⅲ阶段,‘福韵西施’和‘爪艺紫妍氏’各前体物质相对含量在K21-1Y和K21-2Y中均不同程度地高于其相应的绿色区域,且随着进程呈逐渐迅速增加的趋势(ALA 在叶绿素生物合成代谢过程中,催化从ALA到ProtoⅨ反应的酶有氨基乙酰脱氢酶(ALAD)、胆色素原脱氨酶(PBGD)、尿卟啉原Ⅲ合成酶(UROS)、尿卟啉原Ⅲ脱羧酶(UROD)、粪卟啉原Ⅲ氧化酶(CPOX)和原卟啉原氧化酶(PPOX)。本研究测定了亲本‘紫妍氏’叶片及其3个叶艺新品系叶艺区域与绿色区域中这6种叶绿素合成相关酶的活性。将亲本‘紫妍氏’绿叶6种叶绿素合成相关酶活性均设为100%,3个叶艺品系叶绿素合成相关酶活性以相对于亲本绿叶的百分比表示。 从图2可知,与‘紫妍氏’绿叶(K21-G)相比,3个叶艺品系叶片绿色区域的ALAD相对活性有不同程度的提升,其中‘中透紫妍氏’叶片绿色区域(K21-3G)显著提高;三者的PBGD、CPOX和PPOX相对活性的变化不显著(P>0.05);而UROS和UROD相对活性则显著下降(P<0.05)。3个叶艺品系叶片叶艺区域的ALAD相对活性均高于其相应的绿色区域,其中K21-3Y和K21-2Y显著高于K21-3G和K21-2G;K21-1Y和K21-2Y的PBGD、UROS相对活性显著高于其相应的绿色区域,其UROD、PPOX相对活性则与其相应的绿色区域差异不显著,其COPX相对活性却显著低于其相应的绿色区域。同时,K21-3Y的PBGD、UROD、COPX相对活性与K21-3G差异不显著,其UROS和PPOX相对活性显著低于K21-3G。因此,叶艺区域叶绿素合成相关酶活性表现与上述叶绿素生物合成前体物质含量测定的结果相似,都表明K21-1和K21-2叶艺区域叶绿素合成受阻于尿卟啉原 Ⅲ(Urogen Ⅲ)与粪卟啉原 Ⅲ(Coprogen Ⅲ)之间,K21-3 叶艺区域叶绿素合成受阻于胆色素原(PBG)与尿卟啉原Ⅲ(Urogen Ⅲ)之间。 ALAD. 氨基乙酰脱氢酶;PBGD. 胆色素原脱氨酶;UROS. 尿卟啉原Ⅲ合成酶;UROD. 尿卟啉原Ⅲ脱羧酶;CPOX. 粪卟啉原Ⅲ氧化酶;PPOX. 原卟啉原氧化酶图2 ‘紫妍氏’不同品系叶片叶绿素合成代谢途径相关酶活性ALAD. Aminoacetyl dehydrogenase; PBGD. Porphobilinogen deaminase; UROS. Uroporphyrinogen Ⅲ synthase; UROD. Uroporphyrinogen Ⅲ decarboxylase; CPOX. Coproporphyrinogen Ⅲ oxidase; PPOX. Proporphyrinogen oxidaseFig.2 Relative activities of chlorophyll synthesis pathways related enzymes in leaves of different ‘Purple Element’ strains 叶绿素荧光动力学参数能够有效地表征光能的利用、传递和耗散等作用。光系统Ⅱ最大光化学效率(Fv/Fm)反映PS Ⅱ反应中心处于开放时的量子产量,是PS Ⅱ最大的光化学量子产量,能够反映PS Ⅱ天线色素捕光效率;实际光化学效率Y(Ⅱ)反映实际光化学量子产量,较高的Y(Ⅱ)值表示其光能转化效率高,利于合成更多的ATP和NADPH。同时,光化学淬灭系数(qP)反映的是PSⅡ天线色素吸收并用于光化学电子传递的那部分光能,其在一定程度上反映了PSⅡ反应中心开放部分的比例;非化学淬灭系数(NPQ)反映的是PSⅡ天线色素吸收并以热能方式耗散掉的那部分光能,其值越高,说明对光合机构的保护作用越强。由表2可见,各叶艺品系绿色区域的Fv/Fm、Y(Ⅱ)、qP值,以及K21-1G和K21-3G的NPQ值均与K21-G差异不显著,但K21-2G的NPQ值却显著低于K21-G,即叶艺品系绿色区域PS Ⅱ天线色素捕光效率、光能转化效率和光化学电子传递均没有受到显著影响。与相应的绿色区域相比,各叶艺品系叶艺区域的Fv/Fm、Y(Ⅱ)值均显著降低,其qP值均显著提高,K21-1Y、K21-2Y的NPQ值显著降低,K21-3Y的NPQ值却显著提高。其中,K21-1Y、K21-2Y和K21-3Y的Fv/Fm分别为其相应绿色区域的31.45%、69.69%、80.23%,Y(Ⅱ)值分别为其相应绿色区域的53.22%、63.82%、78.42%,qP值分别为其相应绿色区域的3.43、3.05、3.03倍,NPQ值分别为其相应绿色区域的37.43%、81.95%、128.70%。可见,叶艺品系叶艺区域光化学能力下降,光能转化效率变低,且K21-1叶片叶艺区域受影响程度最高,K21-2次之,K21-3最低;K21-1叶片叶艺区域的PSⅡ反应中心开放程度最高,电子传达能力最强,K21-2次之,K21-3最弱; K21-3叶片叶艺区域对光合结构的保护能力最强,K21-2次之,K21-1最弱。 表2 杂交兰不同品系叶片叶绿素荧光参数 叶片结构与光合作用及生长发育机理等密切相关。通过对‘紫妍氏’及其3种叶艺品系的叶片显微结构进行观测(图3)发现,不同品系叶片均主要由上表皮、栅栏组织、海绵组织和下表皮组成,但其中栅栏组织不明显,主要为海绵组织;亲本‘紫妍氏’绿叶(K21-G-A)和3个叶艺品系绿色区域(K21-1G-A、K21-2G-A、K21-3G-A)的海绵组织细胞中含有较多的叶绿体淀粉粒,K21-3叶片叶艺区域(K21-3Y-A)的海绵组织细胞中含有少量叶绿体淀粉粒,而K21-1、K21-2叶片叶艺区域(K21-1Y-A、K21-2Y-A)的海绵组织细胞中未见明显的叶绿体淀粉粒。同时,一个视野中叶片气孔数的统计结果表明,K21-3叶片绿色区域(K21-3G-B)中的气孔数量少于亲本绿叶(K21-G-B)和其他2个叶艺品系绿色区域(K21-1G-B、K21-2G-B);K21-2和K21-3叶片叶艺区域(K21-2Y-B、K21-3Y-B)的气孔数量少于其相应的绿色区域(K21-2G-B、K21-3G-B)。另外,对气孔的开合程度分析发现,K21-1和K21-2叶片叶艺区域(K21-1Y-B、K21-2Y-B)气孔开度要低于其相应的绿色区域(K21-1G-B、K21-2G-B)。 A.叶片显微结构; B.叶片气孔特征图3 杂交兰不同品系叶片显微结构和气孔特征A. Leaf microstructure; B. Stomatal characteristics of leavesFig.3 Microstructure and stomatal characteristics in leaves of different strains of Cymbidium hybrid 前面的杂交兰叶艺品系叶片叶艺区域的叶绿素含量明显降低,我们推测叶艺区域的叶绿体结构可能发生了变化,或者是叶绿体的发育受到了一定的影响。为了进一步了解叶艺品系叶绿体的发育情况与叶绿素含量的关系,本研究利用透射电镜观测分析了‘紫妍氏’及其3个叶艺品系叶片叶绿体超微结构。结果(图4)表明,‘紫妍氏’绿叶(K21-G)和3个叶艺品系叶片绿色区域(K21-1G、K21-2G、K21-3G)的叶绿体被膜清晰,大而饱满;叶绿体基粒颜色较深,基粒的片层结构清晰,嗜锇颗粒分散且数量较少;其中K21-G和K21-1G、K21-3G的叶绿体中部较厚呈梭形,而K21-2G的叶绿体多为紧凑排列,呈椭圆形。3个叶艺品系叶片叶艺区域(K21-1Y、K21-2Y和K21-3Y)的超薄切片着色较绿色区域浅,且叶绿体发育成熟程度也低于绿色区域;其中K21-1Y、K21-2Y的叶绿体被膜基本可见,但未看到明显的基粒片层,嗜锇颗粒较绿色区域多,叶绿体内有空化小泡,较多而且积聚在一起;K21-3Y的叶绿体被膜模糊且破损严重,基本上不具备正常叶绿体的结构特征,无基粒片层,存在嗜锇颗粒和空化小泡,几乎呈现出一种空腔化的结构。 Ce.叶绿体被膜;Og.嗜锇颗粒;Gl.基粒片层;Sg.淀粉粒: Cv.叶绿体内空化小泡图4 杂交兰不同品系叶片叶绿体的超微结构Ce. Chloroplast envelope; Og. Osmiophilic granule; Gl. Grana lamella; Sg. Starch granule; Cv. Cavitation vesicles in chloroplastFig.4 Ultrastructure of chloroplast in leaves of different strains in Cymbidium hybrid 植物叶色变异的发生与色素类物质合成与降解、叶绿体发育与形成、核-质信号转导及调控这些代谢途径的相关基因等因素相关[29,30]。植物叶色突变体的光合色素含量一般均低于其相应的野生型,如铁皮石斛白绿杂色突变体和绿黄杂色突变体[31]、泡桐黄化突变体[32]、西瓜叶色黄化突变体[33]等。本研究中,3个叶艺品系叶片叶艺区域中的叶绿素a、叶绿素b及类胡萝卜素含量均显著低于对照(K21绿色叶片和相应的叶片绿色区域),与前人的研究结果一致。叶色突变体光合色素的变化不仅体现在含量的减少,其Chl a/b值和Caro/Chl值也常常发生改变[13]。本研究中K21-3叶片叶艺区域中Chl a/b值和Car/Chl值均显著高于对照,说明Chl降低程度大于Car,且 Chl b 降低程度大于 Chl a,推测K21-3叶艺形成与叶绿素b缺乏有关;而K21-1和K21-2叶片叶艺区域中仅Chl a/b值显著低于对照,说明 Chl a 降低程度大于 Chl b,推测K21-1和K21-2叶艺的出现与叶绿素a缺乏有关。 植物叶绿素荧光特性与叶绿素含量密切相关,其常常随着叶绿素含量的变化而发生改变,与叶绿体的形成与发育程度也有一定的相关性[34]。本研究中,3个叶艺品系叶片叶艺区域中的Fv/Fm值和Y(Ⅱ)值均显著低于其相应的叶片绿色区域,而其qP值则显著高于其相应的叶片绿色区域,推测这3个叶艺品系叶片叶艺区域在电子传递过程或光能转化过程受到影响,光化学能力下降,光能转化效率降低,但反应中心开放程度高,电子传达能力强。 植物叶绿素的生物合成是由一系列酶催化反应组成的,该途径中任何一步发生变化都有可能阻碍叶绿素的生物合成,导致叶色改变[35]。目前,已在植物多种叶色突变体中发现其叶绿素合成的受阻位点,如水稻叶色突变体在UrogenⅢ到ProtoⅨ合成过程受阻[36];芥菜型油菜黄化突变体在CoprogenⅢ到ProtoⅨ转化过程中受到了阻遏[37]。本研究中,K21-1和K21-2叶片叶艺区域叶绿素合成的受阻位点可能在由Urogen Ⅲ脱羧产生Coprogen Ⅲ间的反应步骤,而K21-3叶片叶艺区域叶绿素合成的受阻位点可能在由PBG合成Urogen Ⅲ间的反应步骤。 叶色突变体中叶绿体的结构和数量往往存在一定的变化[20]。墨兰叶艺品系叶片绿色区叶绿体存在不同程度的发育缺陷,而黄色区的叶绿体更是没有完整的叶绿体结构[18];文心兰黄化突变体中黄叶组织内叶绿体减少,部分叶绿体异常[19];水稻黄绿叶突变体内叶绿体数量明显减少,且叶绿体内没有基粒类囊体[38]。本研究中,3个叶艺品系叶片叶艺区域超薄切片着色较浅,叶绿体发育成熟程度明显低于绿色区域,推测该区域叶绿体的结构和功能都已受到影响;同时叶片叶艺区域的基粒片层结构解体甚至消失,嗜锇颗粒大量增加,这种变化与衰老叶片中退化叶绿体的结构特点有相似之处,反映了叶绿体内膜脂代谢的不平衡[39-40]。 综上所述,杂交兰紫妍氏3个叶艺品系叶片的叶艺区域均存在光合色素含量低,光化学效率低,叶绿体发生了不同程度的退化等现象。不同的是,叶艺品系K21-1和K21-2表现为叶绿素a缺乏,且叶绿素生物合成受阻于尿卟啉原 Ⅲ(Urogen Ⅲ)脱羧产生粪卟啉原 III(Coprogen Ⅲ)的反应步骤;而K21-3表现为叶绿素b缺乏,且叶绿素生物合成代谢受阻位点可能在由胆色素原(PBG)到尿卟啉原Ⅲ(Urogen Ⅲ)的反应步骤。本研究结果表明叶绿素前体物质合成受阻和叶绿体结构发育不良,影响了杂交兰紫妍氏叶片中叶绿素的正常合成,造成叶绿素含量降低,导致叶艺品系叶色改变,植株光能利用率下降。2.3 杂交兰‘紫妍氏’及其叶艺品系叶片叶绿素合成相关酶活性比较

2.4 ‘紫妍氏’与叶艺品系叶片叶绿素荧光参数比较
2.5 ‘紫妍氏’及其叶艺品系叶片显微结构

2.6 ‘紫妍氏’及其叶艺品系叶片叶绿体的超微结构
3 讨 论
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
园林科技(2020年2期)2020-01-18 03:28:18
中国果业信息(2019年1期)2019-01-05 17:41:42
生物学教学(2017年9期)2017-08-20 13:22:32
广西林业科学(2016年1期)2016-03-20 05:33:01
食品工业科技(2014年6期)2014-05-10 06:04:50
作物研究(2014年6期)2014-03-01 03:39:03
食品科学(2013年6期)2013-03-11 18:20:13
