
木贼类营养器官凯氏带类型的新观察
2021-09-04 03:16:20张倩倩林晓艳顾钰峰刘保东
西北植物学报 2021年7期
张倩倩,林晓艳,顾钰峰,刘保东,*
(1 哈尔滨师范大学,哈尔滨 150025;2 黑龙江省普通高等学校植物生物学重点实验室,哈尔滨 150025)
凯氏带(Casparin strip)是由Robert Caspary于1865年在被子植物根内皮层细胞壁上发现的一种加厚结构,具有防御及选择性控制物质径向运输的功能[1-2]。随着研究的深入,陆续发现凯氏带分布于被子植物叶的内皮层[3]、茎的木栓细胞[4]、根的外皮层[5]等。近年来,在裸子植物根[6]和叶[7-8]的内皮层上也发现了凯氏带。此外,蕨类植物茎[9]和叶[10]的内皮层上也有凯氏带。目前对凯氏带的研究主要集中在生理功能[11-13]和分子调控机制[14-16]等方面,如:玉米根凯氏带对镧离子具有阻滞作用[2]、CASPs家族和两个低表达量的小肽影响拟南芥根凯氏带的形成[17-18]。关于凯氏带的化学成分即木质素、多糖类和氨基酸等物质的代谢、合成机理[6,19]等也是目前的研究热点之一。
上述都是基于1层内皮层细胞上的1条凯氏带的研究,尽管在水稻[5]、白屈菜[20]等极少数植物根的外皮层中有过关于2条凯氏带的描述,但都过于零散或仅辅以黑白简图。早在1933年Johnson[21]就关注过Equisetumscirpoide根双内皮层的问题,直到1983年的《维管植物比较形态学》中仍只用线条图简要勾勒了E.sylvaticum地下茎的内外两层内皮层结构[22]。古老的木贼科(Equisetaceae)具有复杂的地上茎、地下茎和根系,是研究植物营养器官起源演化的重要类群,遗憾的是除上述研究之外,至今未发现关于木贼科2条凯氏带的报道。本研究从不同生境的木贼亚属(Subg.Hippochaete)和问荆亚属(Subg.Equisetum)[23]中选择5个种,就地上茎、地下茎和根的内皮层,尤其是凯氏带的分布规律,首次进行了比较观察,发现地下部分都只有1条凯氏带,地上茎多为2条凯氏带。为维管植物营养器官的起源演化、凯氏带的分子调控及生理功能等研究提供了新的资料,讨论了地上茎与地下茎的结构性区别,以及凯氏带与厚壁组织的协作关系等。
1 材料和方法
1.1 材 料
5种木贼科植物的相关信息见表1。凭证标本存于哈尔滨师范大学植物标本室(HANU)。
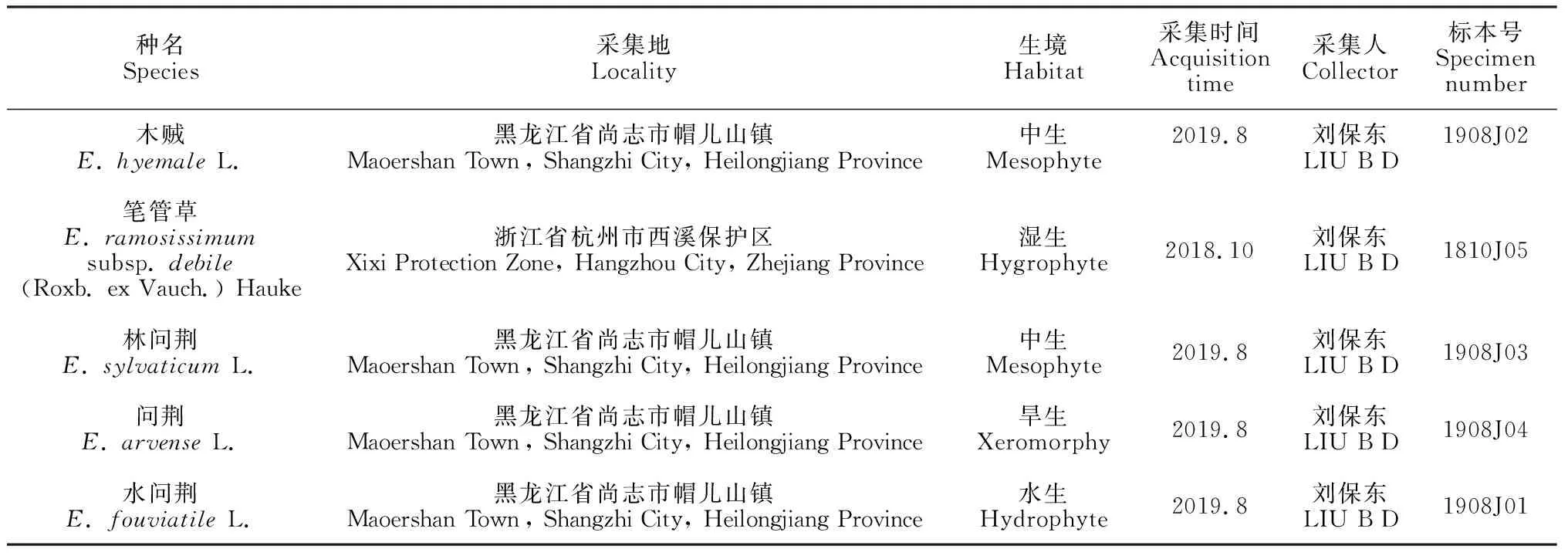
表1 材料来源一览表
1.2 方 法
观察水问荆地下形态并拍照;选取5种木贼科植物的成熟营养器官,经FAA固定、梯度酒精脱水、石蜡包埋等步骤进行常规石蜡切片,切片厚度为8~10 μm,番红-固绿对染,中性树胶封片,用Olympus BX53显微镜观察拍照,并选择典型照片排版。
2 结果与分析
2.1 木贼亚属营养器官凯氏带分布
2.1.1 地上茎木贼(E.hyemaleL.)地上茎横切面近圆形(图版Ⅰ,1),具有凹陷的气孔(St),厚壁组织(S)由4~15层细胞构成,约10层嵌入薄壁组织(Pa),薄壁组织中分布14~16个槽沟(V),髓腔(P)约占总面积的二分之一。薄壁组织内侧有脊沟(图版Ⅰ,2),脊沟外侧有维管束(图版Ⅰ,3),二者周围有2层内皮层围绕,即外侧内皮层(紫虚线示)波折程度较大,内侧内皮层(黄虚线示)波折程度较小,个别内皮层细胞上有凯氏带(CS,白箭头示)。笔管草[E.ramosissimumsubsp.debile(Roxb. ex Vauch.) Hauke]地上茎(图版Ⅰ,4)髓腔较大,有2层内皮层(图版Ⅰ,5),外层(紫虚线示)波折程度较大,而内层(黄虚线示)波折程度较小,细胞间有明显的凯氏点(白箭头示)。
2.1.2 地下茎木贼地下茎横切面近圆形(图版Ⅰ,6),表皮下有6~7层细胞构成的厚壁组织,薄壁组织中均匀分布着9~10个槽沟,中央的髓腔约占总面积的五分之一。脊沟位于维管束内侧(图版Ⅰ,7),每个维管束和脊沟都由1层内皮层(黑虚线示)包围,相邻内皮层细胞间有明显的凯氏点(白箭头示),但靠近髓腔的内皮层细胞和凯氏点不明显。笔管草地下茎的结构与木贼基本相同。
2.1.3 根笔管草根(图版Ⅰ,8)由外及内依次是表皮(E)、厚壁组织(S)、薄壁组织(Pa)及内皮层(图版Ⅰ,9,黑虚线示)。1层内皮层的个别细胞上有凯氏带(CS),相邻细胞间有凯氏点(白箭头示)。维管束中央为后生木质部管胞(MT),周围有4个原生木质部管胞(PT)。木贼根的结构与笔管草基本相同。
2.2 问荆亚属营养器官凯氏带分布
2.2.1 地上茎林问荆(E.sylvaticumL.)地上茎(图版Ⅰ,10)的表皮下有厚壁组织和薄壁组织,薄壁组织中分布9个槽沟,中央的髓腔约占总面积的五分之一。只有1层内皮层(黑虚线示),内皮层的相邻细胞间有凯氏点(图版Ⅰ,11,白箭头示)。问荆(E.arvenseL.)地上茎(图版Ⅰ,12;图版Ⅱ,1)有2层内皮层,2层内皮层上都有凯氏带(图版Ⅰ,12,CS)和较明显的凯氏点(白箭头示),外侧内皮层(紫虚线示)由一层细胞构成,偶有相临的多个内皮层细胞愈合成复合内皮细胞(complex endodermis cell)(图版Ⅱ,1,CE),外层(紫虚线示)波折程度比内层(黄虚线示)大。水问荆(E.fouviatileL.)地上茎的结构与问荆基本相同。
2.2.2 地下茎林问荆地下茎(图版Ⅱ,2)只有1层内皮层(黑虚线示),细胞上有明显的凯氏带(CS),相邻细胞间有凯氏点(白箭头示)。问荆和水问荆地下茎的结构与林问荆基本相同。

图版Ⅱ 3种木贼科(Equisetaceae)植物营养器官横切及水问荆(Equisetum fluviatile L.)地下营养器官形态C.脊沟;CE.复合内皮细胞;V.槽沟;CS.凯氏带;VB.维管束;Pa.薄壁组织;E.表皮;FR.细根;Rhi.地下茎;Sh.芽;CR.粗根;Par.通气道;S.厚壁组织;PX.初生木质部;紫虚线示外侧内皮层;黄虚线示内侧内皮层;黑虚线示单层内皮层;白箭头示凯氏点1.问荆地上茎复合内皮细胞;2.林问荆地下茎内皮层放大;3.水问荆细根横切;4.水问荆地下营养器官形态;5.水问荆粗根横切;6.水问荆粗根内皮层放大。Plate Ⅱ Transverse section of vegetative organs of three species of Equisetaceae and the morphology of underground vegetative organs of Equisetum fluviatile L.C. Carinal canal; CE. Complex endodermis cell; V. Vallecular canal; CS. Casparian strip; VB. Vascular bundle; Pa. Parenchyma; E. Epidermis; FR. Fine root; Rhi. Rhizome; Sh. Shoot; CR. Coarse root; Par. Parichnos; S. Sclerenchyma; PX. Primary xylem; Purple dotted line shows that outer endodermis; Yellow dotted line indicates inner endodermis; Black dotted line illustrates single layer endodermis; White arrow shows Casparian dotsFig.1. Complex endodermis cell of aerial stem of E. arvense L.; Fig.2. Endodermis enlargement of rhizome of E. sylvalicum L.; Fig.3. Transverse section of fine root of E. fluviatile L.; Fig.4. Morphology of underground vegetative organs of E. fluviatile L.; Fig.5. Transverse section of coarse root of E. fluviatile L.; Fig.6. Endodermis enlargement of coarse root of E. fluviatile L..
2.2.3 根水问荆细根(图版Ⅱ,3)由外及内依次为表皮(E)、薄壁组织(Pa)、内皮层(黑虚线示)及维管束(VB),内皮层细胞间的凯氏点(白箭头示)较明显。水问荆粗根(图版Ⅱ,4)数量很少,呈光滑的圆柱状,既不分节也不分枝,且无气孔和根毛。表皮下有厚壁组织(图版Ⅱ,5),由2层细胞构成,其内侧为薄壁组织,薄壁组织间有多数不规则的通气道(Par)。只有1层内皮层(图版Ⅱ,6,黑虚线示)包围着维管束,内皮层细胞间都具有凯氏点(白箭头示),初生木质部(PX)较均匀地分布于维管束中。林问荆和问荆根的结构与水问荆细根基本相同。
综上所述,本研究所观察木贼科5个种地上茎、地下茎及根的凯氏带特征归纳为表2。
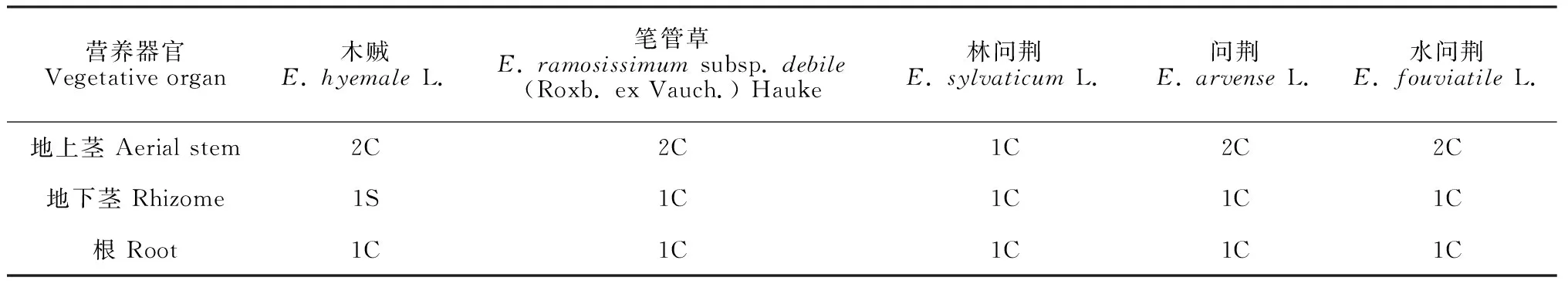
表2 5种木贼科植物营养器官凯氏带特征
3 讨 论
部分学者根据木贼科地上茎分枝的多寡、常绿与否、孢子囊穗顶端是否为钝形等特征,主张将木贼科分为两个亚属,即木贼亚属和问荆亚属[23-24]。而本研究所观察的木贼科2个亚属5个种的地下茎和根都只有1条凯氏带。仅就地上部分而言,尽管木贼亚属的木贼和笔管草都有2条凯氏带,但问荆亚属既有1条也有2条,如问荆亚属的问荆和水问荆都有2条,而林问荆只有1条,所以笔者认为,木贼类植物凯氏带数量不能作为分类的依据。
就木贼类地上茎和地下茎的本质而言,一般观点认为二者是相同的,因为地上茎一旦被埋入地下,其节上休眠的根原基就都能发育出正常的根[22]。而本研究对5个种的分析发现,仅林问荆地上茎和地下茎的凯氏带数量相同,都是1条,而其余4个种的地上茎都是2条,地下茎都是1条,因此地上茎与地下茎是有显著的结构性区别的。木贼科地上茎多有2条凯氏带,而地下茎和根都只有1条,就功能而言,地下部分的1条凯氏带应该不及地上部分的2条凯氏带,但地下部分都有发达的厚壁组织或(和)与其紧密相连的表皮,这似乎表明厚壁组织或(和)表皮具有与凯氏带相同的功能,这应该与植物根系外皮层细胞壁加厚可以控制离子运输的观点[25-27]一致。
凯氏带的主要功能是控制水分和离子进出维管柱[2,27],进而对胁迫中的植物体起到保护作用[28-29]。本实验发现,不同类型的凯氏带防御能力也不同:2层公共内皮层上的凯氏带,既能保护维管束,也能抵御来自外界环境和内部髓腔的伤害;1层散生内皮层上的凯氏带,只能保护其内部的维管束,并不能阻止外界和髓腔的物质交流;而1层公共内皮层上的凯氏带,只能抵御来自外界环境对维管束的伤害,却不能抵御来自髓腔的伤害。因此,我们认为3种类型凯氏带的防御能力由强到弱依次是:2层公共内皮层上的凯氏带 > 1层散生内皮层上的凯氏带 > 1层公共内皮层上的凯氏带。此外,内皮层的分化能够阻断相邻细胞的质外体运输途径,不仅能控制物质进入中柱[30],也能阻止营养物质回流[12,31],而问荆地上茎外侧内皮层中偶有多个内皮层细胞愈合成较大的复合细胞,即复合内皮细胞,该细胞似乎降低了物质运输效率,但从原生质体的主动运输能力来看,该类细胞应该能增强内皮层的双向调控能力。复合内皮细胞在真蕨类根、茎及叶中都较常见(论文将另文发表),似乎表明木贼类与真蕨类[32]有着类似的演化渊源,而木贼类的复合内皮细胞较少,从形态结构与数量来看,似乎支持将木贼科归为最原始真蕨类的分类方案[33]。
猜你喜欢
幼儿画刊(2020年4期)2020-05-16 02:53:14
浙江农业学报(2020年1期)2020-03-07 05:07:12
幼儿画刊(2019年2期)2019-04-08 01:23:46
浙江农业科学(2017年8期)2017-08-30 12:48:08
浙江农业科学(2016年5期)2016-12-07 07:03:27
——系统分类与检索*
寄生虫与医学昆虫学报(2015年3期)2015-11-12 00:32:38
首都食品与医药(2015年13期)2015-11-03 08:54:34
中老年健康(2015年9期)2015-05-30 03:12:00
家庭医学(2015年2期)2015-05-06 01:02:15
红蜻蜓·低年级(2009年9期)2009-05-31 09:25:37
