
辣椒CaNAC55基因克隆与表达分析
2021-09-04 03:16:16刁卫平王述彬刘金兵潘宝贵隋益虎
西北植物学报 2021年7期
吴 荡,刁卫平,王述彬,刘金兵,潘宝贵,隋益虎
(1 安徽科技学院 农学院,安徽凤阳233100;2 江苏省农业科学院 蔬菜研究所,江苏省高效园艺作物遗传改良重点实验室,南京 210014)
辣椒(CapsicumannuumL.)作为一种重要的蔬菜作物,具有重要的经济价值[1]。其生长发育过程中经常受到病害及非生物胁迫因素的影响,从而影响品质和产量[2-5]。为了降低这些影响,植物进化出一套抵御外界胁迫的反应机制,即通过转录因子整合外界信号和基因调控的方式产生相应的蛋白和代谢产物来适应胁迫,从而提高植物的抗逆性[6]。
NAC是近年来发现的植物独有的一类转录因子家族[7-8]。它的命名来源于矮牵牛NAM(No Apical Meristem)基因和拟南芥ATAF1/2基因以及CUC2(cup-shaped-cotyledon)基因的首字母。1997年,Aida等[9]首先报道并命名了NAC结构域,所有的NAC转录因子N端都含有一个高度保守NAC结构域,它由150~160个氨基酸组成,分为A、B、C、D、E 5个亚结构域。NAC转录因子还有一个可充当转录激活区域的高度多样化的C端序列,因此NAC蛋白可能表现不同的功能[10]。最早在水稻中把NAC转录因子家族分为3个亚族(NAM、OsNAC3和ATAF)为Ⅰ族[11],Ooka等[12]根据NAC结构域的相似性又将NAC转录因子家族分为第Ⅰ和Ⅱ两大族,其后进一步根据自展支持率对其分别细分为14个和4个亚族。随着测序技术的迅速发展和大量研究结果的发表,发现NAC基因家族中有一些基因同时参与生物和非生物胁迫反应。其中水稻OsNAC5、OsNAC9和OsNAC10基因的超表达,不仅可以提高水稻的产量,还可以提高水稻的抗旱性[13-14]。番茄StNAC基因在致病疫霉菌的侵染下诱导表达[15]。然而,当前关于辣椒NAC转录因子的研究并不太多。CaNAC1的表达不仅受到病原菌的快速特异诱导,还受到外源水杨酸和乙烯的强烈诱导[16]。CaNAC2和CaNAC23的表达均受到干旱和盐害等非生物胁迫的诱导[17-18]。CaNAC036和CaNAC72均是通过JA/ET信号传导途径参与青枯病菌侵染的防御反应[19-20]。CaNAC45和CaNAC61在辣椒逆境胁迫方面起到了重要作用[21-22]。
项目组前期在辣椒全基因组水平上共鉴定到104个NAC转录因子,依次命名为CaNAC1~CaNAC104[23],本试验以CaNAC55为研究对象,选用高抗疫病辣椒材料CM334为试材,分析转录因子的序列结构以及在不同生物和非生物胁迫下的表达模式,为后续深入研究NAC转录因子的功能提供参考。
1 材料和方法
1.1 试验材料准备
挑取籽粒饱满种子用纱布包好,55 ℃热水浸种20 min,然后在室温条件下让种子充分吸水6~8 h,最后播种于72孔穴盘中,每穴1~2粒种子。子叶出土后,将幼苗转移到光照培养箱中长日照条件下生长(16 h光照,8 h黑暗),温度为26 ℃(昼)/18 ℃(夜),当辣椒幼苗有6片真叶时,用于胁迫处理。
1.2 方 法
1.2.1 胁迫处理利用本课题组已建立的辣椒疫霉菌接种体系[24]对幼苗进行接种,向在V8培养基培养的疫霉菌中加入15 mL灭菌超纯水,将培养皿于4 ℃放置30~40 min,再取出室温放置30 min左右,在显微镜下观察游动孢子,用血球计数板调整其浓度为1×105spores/mL备用,分别将幼苗放入光照强度为160 μmol·m-2·s-1的光照培养箱内进行胁迫处理。利用事先备好的1×105spores/mL疫霉菌对幼苗进行接种;将幼苗根洗净后分别放置在300 mmol/L NaCl和400 mmol/L甘露醇溶液中进行盐胁迫和干旱胁迫处理;将幼苗放入42 ℃条件下的光照培养箱内进行热激胁迫处理;分别利用100 mmol/L ABA、SA、MeJA溶液喷施辣椒叶面,用无菌水喷洒作对照[25],分别于处理0、3、6、12和24 h取样,液氮速冻,-80 ℃超低温保存备用,生物学重复3次。
1.2.2 基因克隆在茄科基因组数据库(https://solgenomics.net/)中下载已有CaNAC55基因序列,并根据序列特点设计引物(表1),委托擎科生物技术有限公司(南京)合成。以CM334材料的cDNA和gDNA作为扩增模板,PCR反应程序为:98 ℃预变性3 min;98 ℃变性10 s,56 ℃退火20 s,72 ℃延伸20 s,35个循环;72 ℃延伸5 min,12 ℃保存。1%琼脂糖凝胶电泳(110 V,30 min)检测PCR产物,质量合格后回收相应的gDNA片段,回收片段克隆至pClone007载体(TsingKe,南京),热激法转化大肠杆菌感受态DH5α,挑选阳性单克隆送擎科生物技术有限公司(南京)测序,得到目的片段序列。
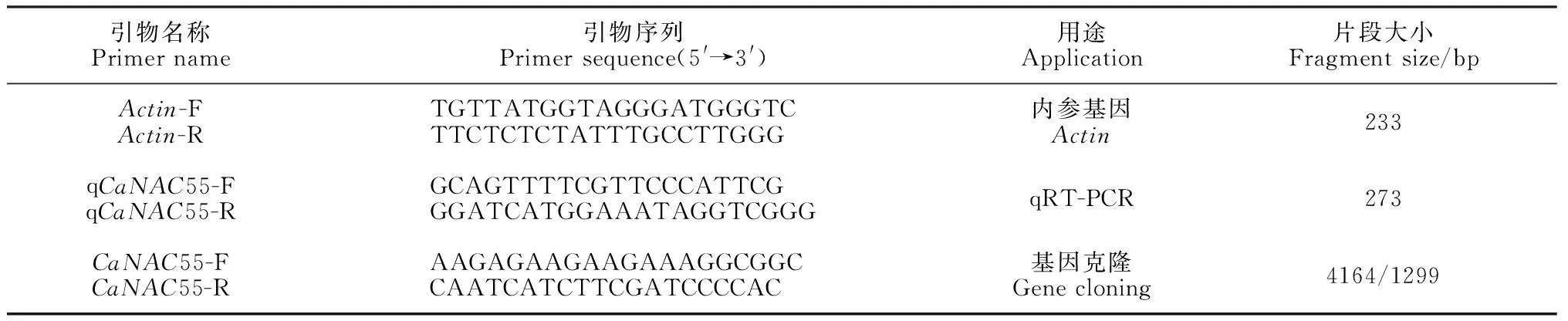
表1 所用引物信息
1.2.3 生物信息学分析利用ORFfinder程序(http//www.ncbi.nlm.nih.gov/gorf/gorf.ht-ml)对CaNAC55基因进行开放阅读框(ORF)预测;根据克隆所得的cDNA序列,利用NCBI的Blastp在线分析工具对氨基酸序列进行同源性比对,并结合IQTREE构建分子进化树。同时,选取拟南芥、水稻和辣椒不同亚家族NAC转录因子(ANAC011,ANAC022,ANAC029,ANAC055,ANAC078,ANAC083,ANAC084,ANAC090,ANAC091,ANAC098,ONAC022,ONAC074,CaNAC045,CaNAC046,CaNAC100)进行进化树构建。
1.2.4CaNAC55在胁迫下的表达分析提取辣椒叶片总RNA,并按照试剂盒的方法反转录得到cDNA(TaKaRa,大连)。根据辣椒CaNAC55序列设计RT-PCR引物(表1),以辣椒Actin(GenBank登录号为GQ339766)作为内参基因[26],引物由擎科生物技术有限公司(南京)合成。采用德国罗氏诊断有限公司的LightCycler 480 Ⅱ实时荧光定量PCR仪,进行实时荧光定量RT-PCR,反应体系(25 μL)包括2×SYBR Premix Ex Taq 10 μL、10 mmol/L上下游引物各0.8 μL,cDNA 1 μL(100 ng/μL),ddH2O 7.4 μL。反应程序:94 ℃预变性4min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,40个循环;72 ℃ 1 min。处理设3个生物学重复,3个技术重复,所得数据采用2-ΔΔct法计算CaNAC55的相对表达量[25],采用Excel进行平均数统计,以SPASS软件进行方差分析,绘制RT-PCR分析图。
2 结果与分析
2.1 辣椒CaNAC55基因的克隆
利用引物CaNAC55F/CaNAC55R(表1)分别对辣椒高抗疫病材料CM334的gDNA和cDNA进行扩增,扩增获得4 100 bp和1 200 bp左右的片段(图1),将两种目的片段切胶回收后连接转化,挑取阳性克隆用特异性引物检测后送于南京擎科生物科技有限公司测序,并用DNAMAN软件进行序列验证。结果显示,以gDNA和cDNA为模板的目的片段与辣椒基因组数据库中序列完全一致,大小分别为4 164和1 299 bp,该基因从起始密码子ATG到终止密码子TAG之间具有一个1 299 bp的完整开放阅读框(ORF)(图2),其基因编码的蛋白由432个氨基酸残基组成。

图1 辣椒CaNAC55基因PCR扩增 Fig.1 PCR amplification of CaNAC55 gene in pepper

图2 CaNAC55 cDNA序列及预测氨基酸序列Fig.2 CaNAC55 nucleotide sequence and predicted amino acid sequence
2.2 辣椒CaNAC55序列比对
对辣椒CaNAC55基因的氨基酸序列使用NCBI中的Blastp进行同源性检索,应用DNAMAN软件对番茄、烟草、马铃薯等11种NAC家族代表性植物进行NAC多序列比对(图3)。同时,利用IQTree软件将CaNAC55与拟南芥、水稻和辣椒中不同亚族进行进化树构建,根据NAC家族分类特点可知,CaNAC55属于NAC家族第Ⅰ族中的TIP亚族(图4)。

图3 NAC55多序列比对Fig.3 Multiple sequence alignment of NAC55
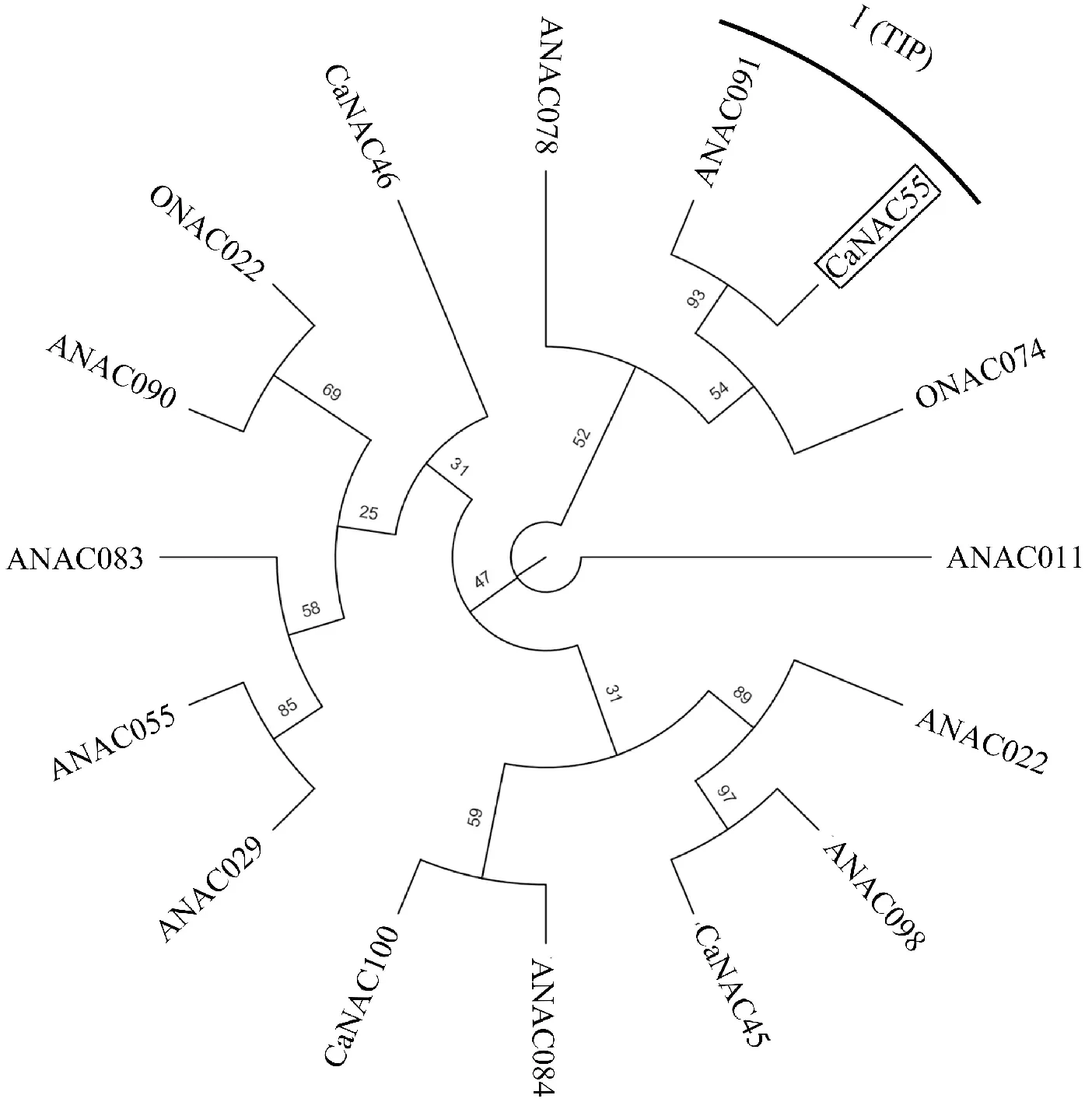
图4 辣椒CaNAC55氨基酸与拟南芥(ANAC)、水稻(ONAC)和辣椒(CaNAC)不同亚族构建进化树Fig.4 Phylogenetic tree of CaNAC55 amino acid sequences from pepper and other species from different subfamilies of Arabidopsis thaliana (ANAC), rice (ONAC) and rice (CaNAC)
2.3 辣椒CaNAC55进化树分析
使用NCBI中Blastp工具对辣椒CaNAC55基因序列进行同源检索,利用IQTree软件对检索到的数据构建系统进化树(图5)。结果表明,辣椒CaNAC55基因与辣椒Capsicumannuum(XM-016722474)、番茄Solanumlycopersicum(XM-004241285)、马铃薯Solanumtuberosum(XM-006361027)亲缘关系最近,相似性依次达到99.87%、93.37%和92.62%。推测CaNAC55基因的功能可能与马铃薯、番茄中的同源基因功能相似。
2.4 辣椒CaNAC55对胁迫的应答
盐害、干旱、热激等非生物胁迫均能诱导CaNAC55基因的表达(图6)。研究结果显示,CaNAC55基因在高盐处理下的相对表达量呈显著上升趋势,在处理后24 h达到峰值,表达量为对照的20.92倍。在干旱处理下的相对表达量呈波浪型变化趋势,在3 h表达量达到峰值,为对照的2.23倍,随后迅速降低至最低后上升,在24 h表达量达到最大峰值,为对照的3.01倍(图6)。CaNAC55基因在42 ℃热激胁迫处理3 h后表达量迅速降低,随后迅速升高,在12 h表达量达到峰值,为对照组的8.84倍(图6)。结果显示,辣椒CaNAC55基因在高盐、干旱、热激等非生物胁迫下表达量均表现一定程度上调,高盐胁迫处理下基因表达上调最为显著,可能在辣椒相应盐胁迫过程中起着重要作用。此外,研究发现辣椒CaNAC55基因同样受到激素胁迫的影响,在ABA处理下,CaNAC55基因的相对表达量均低于对照组,说明CaNAC55基因的表达受到ABA的抑制。在SA、MeJA、phyto等胁迫处理下CaNAC55基因相对表达量与对照组相比无明显变化。

图5 辣椒CaNAC55基因与GenBank中相似序列的进化树分析Fig.5 Conserved domain in phylogenetic tree analysis between pepper CaNAC55 gene and similar sequences in GenBank
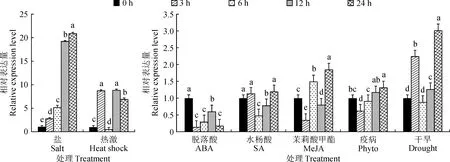
不同小写字母表示在同一胁迫处理下差异显著图6 辣椒CaNAC55基因在不同非生物胁迫中的表达Differentl letters showed significant difference under the same stress treatmentFig.6 Relative expression of CaNAC55 gene of pepper with different abiotic stress treatments
3 讨 论
植物生长发育过程中会面临各种逆境胁迫,前人研究表明NAC转录因子家族不仅在抵抗逆境胁迫时发挥着重要作用,在调节植物生长发育中也起着相关作用。在盐胁迫方面,棉花中的GmNAC4通过维持细胞内的渗透平衡来提高耐盐性[27];利用酵母单杂系统在拟南芥中分离出3个不同的NAC转录因子ANAC055、ANACO19和ANAC072,高盐、干旱、ABA依次诱导这些转录因子表达[28-30];有研究表明在盐和干旱胁迫条件下,辣椒CaNAC23基因的表达量会上升[17]。本研究发现,高盐处理下辣椒CaNAC55基因的表达量呈显著上升趋势,其表达模式与前人研究结果基本一致,推测CaNAC55基因可能在辣椒响应盐胁迫反应过程中起着重要的调节作用。干旱胁迫下,苦芥FtNAC17基因表达量呈波浪型变化,可能与多个基因协同响应干旱胁迫有关[31]。本研究发现,辣椒CaNAC55基因的表达量随着干旱处理时间的增加也呈波浪型变化趋势,推测辣椒CaNAC55基因可能与苦芥FtNAC17基因响应干旱胁迫机制相同。在热激胁迫方面,纪康[32]对高羊茅种质进行了耐热激能力评价后,从中鉴定出一个受热激显著诱导的转录因子FaNAC72。CaNAC55基因在热激处理后表达量与对照组相比增幅较大,处理12 h表达量达到峰值,为对照组的8.84倍。研究结果表明,辣椒CaNAC55基因的表达受到高盐、干旱、热激等非生物胁迫强烈诱导。
前人研究表明,苦荞FtNAC11基因通过调节SA、MeJA、ABA等信号,使其在苦荞响应非生物胁迫中起到负调控作用[33]。SaNAC36基因也在互花米草响应非生物胁迫过程中主要起到负调控的作用[34]。辣椒幼苗在ABA处理后,CaNAC55基因的表达量均低于对照组,该研究结果与前人一致。推测辣椒CaNAC55基因的表达受到激素ABA的抑制,使其在辣椒响应非生物胁迫中起到负调控的作用。在SA、MeJA、phyto等胁迫处理下CaNAC55基因表达量与对照组相比无明显变化。因此,SA、MeJA、phyto等因素不直接参与该基因的代谢调控过程。
CaNAC1的表达受到病原菌、水杨酸和乙烯等胁迫的诱导[16];CaNAC2和CaNAC23的表达受到干旱、盐害等非生物胁迫的强烈诱导[17-18];CaNAC036和CaNAC72在调控青枯病菌侵染的防御反应过程中起到了重要作用[19-20];CaNAC45和CaNAC61的研究为深入探讨该基因在生物代谢过程中的功能奠定了理论基础[21-22]。本项目组前期在全基因组水平上共鉴定104个辣椒NAC转录因子[23],本研究发现CaNAC55转录因子对不同逆境胁迫响应不同,推测辣椒CaNAC55基因可能作为重要的调节因子参与辣椒不同逆境胁迫响应。相信随着现代测序技术和生物信息学的不断进步,越来越多植物的NAC转录因子家族成员将被鉴定和发现,进而通过对NAC转录因子的深入研究来提高作物的抗逆性,促进农业发展。
猜你喜欢
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
少儿科学周刊·少年版(2022年18期)2022-05-30 10:48:04
今日农业(2021年10期)2021-11-27 09:45:24
天津市教科院学报(2021年5期)2021-11-10 07:32:40
小哥白尼(神奇星球)(2021年6期)2021-07-28 06:31:36
生物学通报(2021年9期)2021-07-01 03:24:44
中外文摘(2020年9期)2020-06-01 13:47:56
江苏农业科学(2016年8期)2017-02-15 19:54:11
中国火炬(2015年12期)2015-07-31 17:38:35
淮阴工学院学报(2014年5期)2014-09-10 09:15:56
