
可溶性糖和有机酸对稻田土壤活性有机碳含量与甲烷排放的影响*
2021-09-01 10:14刘成前冯书华何永美张彦雪祖艳群谢春梅
云南农业大学学报(自然科学) 2021年4期
刘成前,冯书华,李 想,何永美,张彦雪,祖艳群,李 元,谢春梅
(云南农业大学 资源与环境学院,云南 昆明 650201)
温室气体大量排放导致全球气候变暖已成为当今重要的环境问题之一[1]。甲烷(CH4)作为温室气体,对温室效应贡献仅次于CO2,贡献率可达15%,增温潜势是CO2的23~25 倍[2]。稻田是重要的CH4排放源之一,中国作为水稻(Oryza sativa)种植大国,水稻种植面积占全球水稻面积的18.5%,稻田CH4排放量占全球排放量的19%[3]。因此,研究稻田CH4排放对控制全球气候变暖具有重要意义。
水稻根系分泌物是影响稻田CH4排放的重要因素,稻田土壤中的产甲烷菌以此为碳源释放CH4[4]。水稻根系分泌物包括氨基酸、低分子量有机酸和可溶性糖等小分子有机物,其中有机酸易被根际细菌分解为CO2、H2和醋酸盐等产CH4基质,为产甲烷菌提供原料,促进稻田土壤CH4的产生[5]。根系分泌物的释放可改变土壤中活性有机碳的组分,为CH4产生提供了重要的底物,稻田CH4排放量中60%来源于土壤活性有机碳[6]。根系分泌物类似物的增加促进微生物活性和酶促反应,加快土壤中物质转化速率[7];土壤微生物是土壤生化反应的驱动力,土壤酶大多数来源于此[8]。有研究表明:向土壤中添加根系分泌物类似物如乙酸盐、甲酸盐、苹果酸盐、草酸盐和糖等,可显著增加土壤中CO2和CH4的排放量[9]。现有研究集中于田间原位添加根系分泌物类似物或覆压植物秸秆增加有机质输入,提高土壤脲酶、过氧化氢酶和蔗糖酶活性等,为了解土壤微生物、酶活性和CH4排放之间的关系提供理论依据[10];多数室内研究主要模拟根系分泌物碳输入促进生物代谢活力、酶活性和基因表达等方面[11],缺少对CH4排放影响的研究。
梯田农业广泛分布于亚洲、欧洲、美洲和非洲,是人类适应山地环境而形成的一种古老的生产模式,目前全球已有14 处古梯田被列入世界文化遗产名录[12],元阳梯田的规模和海拔跨度具有较强的典型性和代表性。元阳哈尼梯田位于云南省元阳县哀牢山南部,海拔1 400~1 800 m,传统水稻种植历史1 500 多年,水稻种植时间为每年5—10 月,冬季休闲期水稻秸秆还田,浮水固氮植物满江红(Azolla imbricate)自然生长,土壤有机碳含量丰富,利于CH4产生,在水稻生长期和冬季休闲期均有CH4排放,具有持续和高通量排放的特点,是CH4排放量最大的一类稻田[13]。土壤有机碳是CH4排放的关键底物,而根系分泌物是根际沉积过程中的一部分,也是植物根系释放有机碳的主要来源[14]。本研究以元阳梯田土壤为对象,模拟水稻根系分泌物增加,研究添加可溶性糖和有机酸对土壤酶活性、土壤活性有机碳组分和含量的影响,探究CH4排放的影响因素,以期为了解梯田系统土壤温室气体排放及其影响因素提供科学依据。
1 材料与方法
1.1 供试材料
稻田土壤样品采集于元阳县新街镇箐口村(N23°7′,E102°44′)梯田,年平均温度15 ℃,年均降水量1 397 mm,年均蒸发量1 543 mm,属于山地季风气候,海拔1 600 m,5—10 月种植水稻,梯田全年淹水,是元阳哈尼梯田核心区[15]。供试土壤背景值为:全氮含量2.54 g/kg,全磷含量0.431 g/kg,全钾含量13.85 g/kg,碱解氮含量92.17 mg/kg,速效磷含量14.03 mg/kg,速效钾含量16.50 mg/kg,土壤pH 值5.67,有机质含量29.93 g/kg,土壤容重1.45 g/cm3,含水率48%。
1.2 厌氧培养试验
使用土壤采样器“S 布点法”采集水稻收获后的稻田0~20 cm 处土壤,迅速运回试验室。直接用新鲜土样过10 目尼龙网筛,以去除水稻秸秆、植物根系和石砾等杂质[15],参照杨玉楠等[16]的方法开展厌氧培养试验。
可溶性糖和有机酸组合溶液质量分数参照SHI 等[17]的方法,设置为可溶性糖0.245 mg/g、草酸0.423 mg/g、琥珀酸0.403 mg/g 和苹果酸0.033 mg/g,相当于在低、中和高的植物光合速率下根系分泌物中的碳含量,碳添加比率为0.1、0.2 和0.3 mg/(g·d) (按土壤干质量计算)。试验共设计10 种根系分泌物类似物及其组合处理,每个处理3 组重复,各处理添加量见表1。组合溶液均用去离子水配制,用NaOH 调节pH值至5.5,定容至150 mL,保存于4 ℃冰箱内。
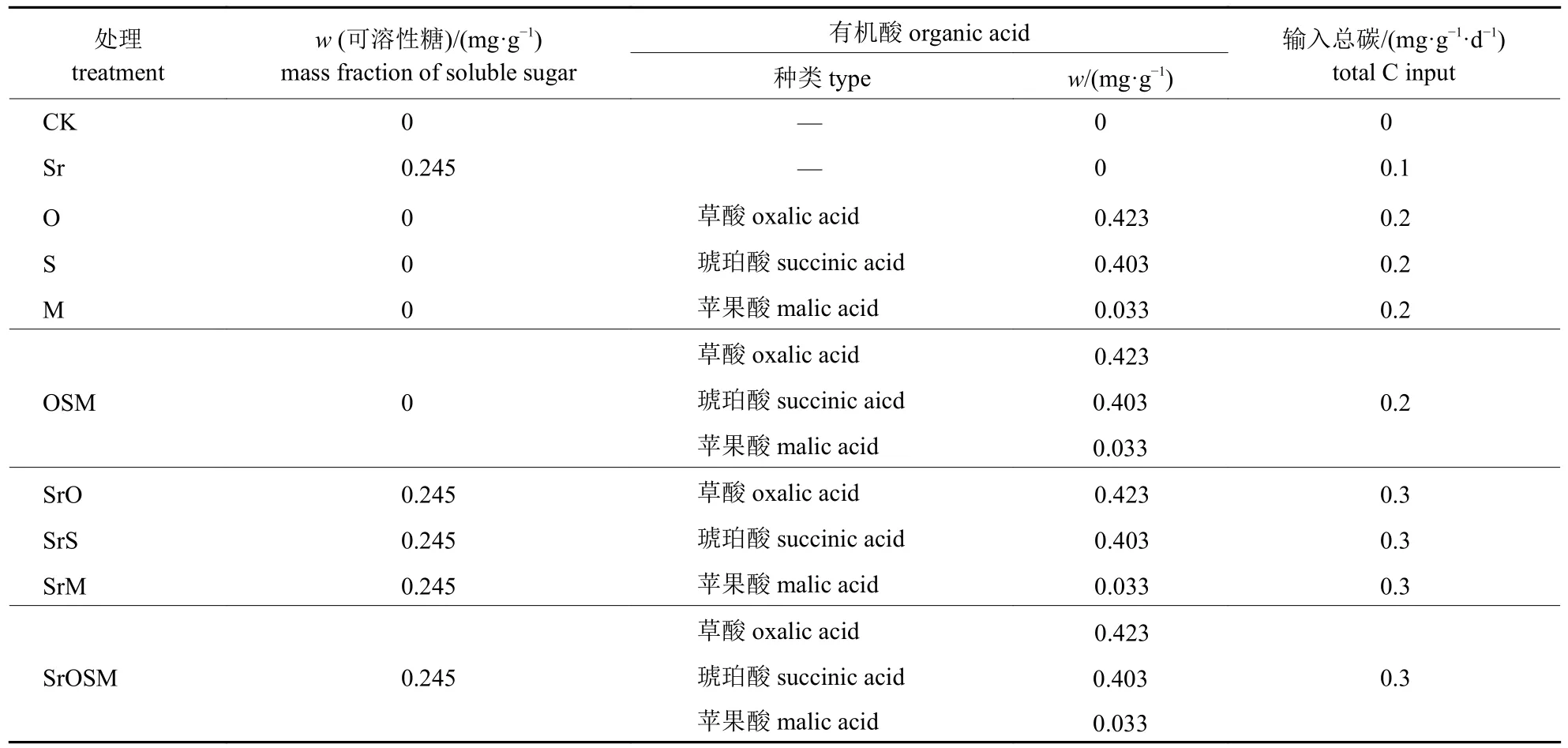
表1 可溶性糖和有机酸的添加组合Tab.1 Addition combination of soluble sugar and organic acid
每个处理取300 g 土壤样品(干质量)加入1 L 棕色广口瓶中,用去离子水淹没3 cm,使其达到厌氧状态,培养2 周,以培养微生物群落。培养结束,用氮气冲洗消散瓶中累积的气体,随后以10 mL/d 的速率加入可溶性糖和有机酸溶液,空白处理加10 mL 高压灭菌的去离子水作为对照(CK),室温下培养2 周。广口瓶用橡胶塞密封,橡胶塞留两口,一口添加可溶性糖和有机酸,另一口连接马氏发酵管;马氏发酵管内装有过量0.5 mol/L NaOH 溶液,用来吸收培养过程中产生的二氧化碳气体,利用马氏发酵管的排液法测得CH4排放量。
1.3 测定方法
直接取瓶中土壤悬液混匀后测定土壤酶活性和有机碳含量,同时测定含水率、确定土壤干质量。
土壤活性有机碳测定采用严昌荣等[18]的方法,硫酸钾浸提,重铬酸钾—硫酸外加热容量法测定。土壤微生物量碳(MBC)按照《土壤与环境微生物研究法》中的改进方法[19],氯仿熏蒸,K2SO4浸提,重铬酸钾—硫酸外加热容量法测定。土壤易氧化有机碳(LOC)采用传统方法[15],高锰酸钾氧化,离心取上清液稀释后比色,根据高锰酸钾的消耗量计算土壤易氧化有机碳含量。
土壤脱氢酶活性测定采用关松荫等[20]的TTC比色法,加TTC Assay buffer 工作液,避光孵育,于485 nm 测定吸光度,计算TTC 还原量,以该还原量表示脱氢酶活性。过氧化氢酶活性测定采用高锰酸钾滴定法[20],加H2O2和铝钾矾震荡后过滤,高锰酸钾滴定,计算H2O2消耗量,以该量表示过氧化氢酶活性。蔗糖酶活性测定采用3,5—二硝基水杨酸比色法[15],加蔗糖和二硝基水杨酸沸水浴加热,于508 nm 测定吸光值,计算蔗糖消耗量,以该消耗量表示蔗糖酶活性。纤维素酶活性测定采用甲苯培养比色法[15],加羧甲基纤维素溶液加热培养,于540 nm 测定吸光值,计算纤维素消耗量,以该消耗量表示纤维素酶活性。多酚氧化酶活性测定采用邻苯三酚—乙醚比色法[15]。加邻苯三酚溶液培养,于525 nm测定吸光值,计算邻苯三酚—乙醚消耗量,以该消耗量表示多酚氧化酶活性。每一土样需做无基质对照。
CH4气体采集;用双联球连接广口瓶导出口,另一端连接集气袋,采集气体,气体样品采用Agilent 7890B 气相色谱仪测定分析[15]。CH4排放量计算公式为:
F=dC/dtt=0×h×ρ×60×273/(273+T)
式中,F为气体排放通量;h为采样瓶高度(液面离顶部的高度);T为瓶内温度;ρ为标准状况下CH4气体的密度,0.717 g/L;dC/dtt=0为单位时间内气体浓度线性变化率。
1.4 数据统计分析
采用Excel 对数据统计整理,用Origin Pro 9.1绘图,用SPSS 20.0 软件进行单因素方差分析,处理组间差异显著性采用LSD 法分析。
2 结果与分析
2.1 添加可溶性糖和有机酸对CH4 排放的影响
由图1 可知:添加可溶性糖和有机酸可影响CH4排放量和排放通量。除Sr 和S 处理外,其他处理的CH4排放量较CK 增加趋势更明显;除S 和OSM 处理外,其他处理的CH4排放通量显著增加(P<0.05),SrM 和SrO 处理增量与CK 相比分别高达124 和197 mg/(m·h)。
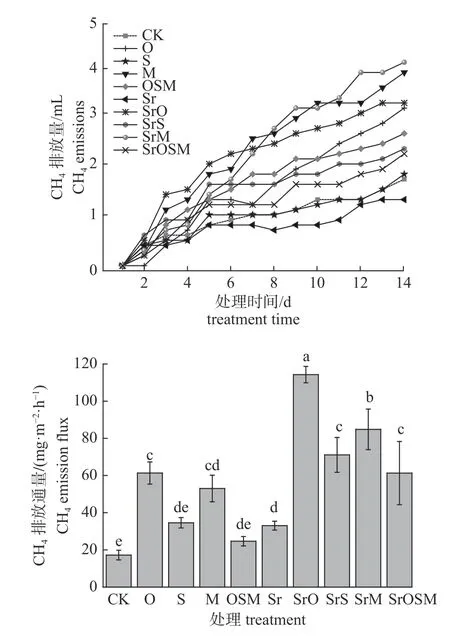
图1 添加可溶性糖和有机酸对CH4 排放的影响)Fig.1 Effect of the addition of soluble sugar and organic acid on the CH4 emission
2.2 添加可溶性糖和有机酸对土壤有机碳和CH4排放通量的影响
由图2 可知:添加可溶性糖和有机酸对土壤微生物量碳和可溶性有机碳含量影响较大,表现为促进作用,易氧化有机碳含量变化不明显。与CK 处理相比,SrO 处理易氧化有机碳含量显著增加,增幅为7.8%,O 和OSM 处理显著降低,降幅分别为10.0%和22.4%,其他6 组处理变化均不显著;除SrS 和SrOSM 处理外,其他处理组的微生物量碳含量均显著高于对照组(P<0.05),SrO 处理组的增量高达250%;8 组处理可溶性碳含量均显著增加,增幅为48.1%~153%。
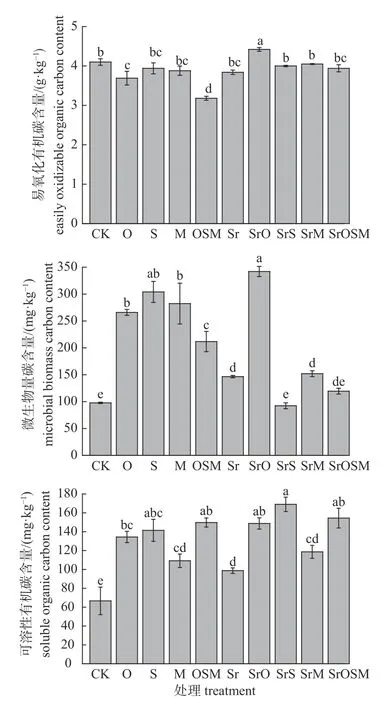
图2 添加可溶性糖和有机酸组分对土壤活性碳含量的影响)Fig.2 Effects of the addition of soluble sugar and organic acid on the activated organic carbon content in soil
由图3 可知:易氧化有机碳含量与CH4排放通量呈极显著正相关(P<0.01),微生物量碳与CH4排放通量显著正相关(P<0.05),表明土壤中微生物量碳含量和易氧化有机碳含量与CH4排放通量密切相关。
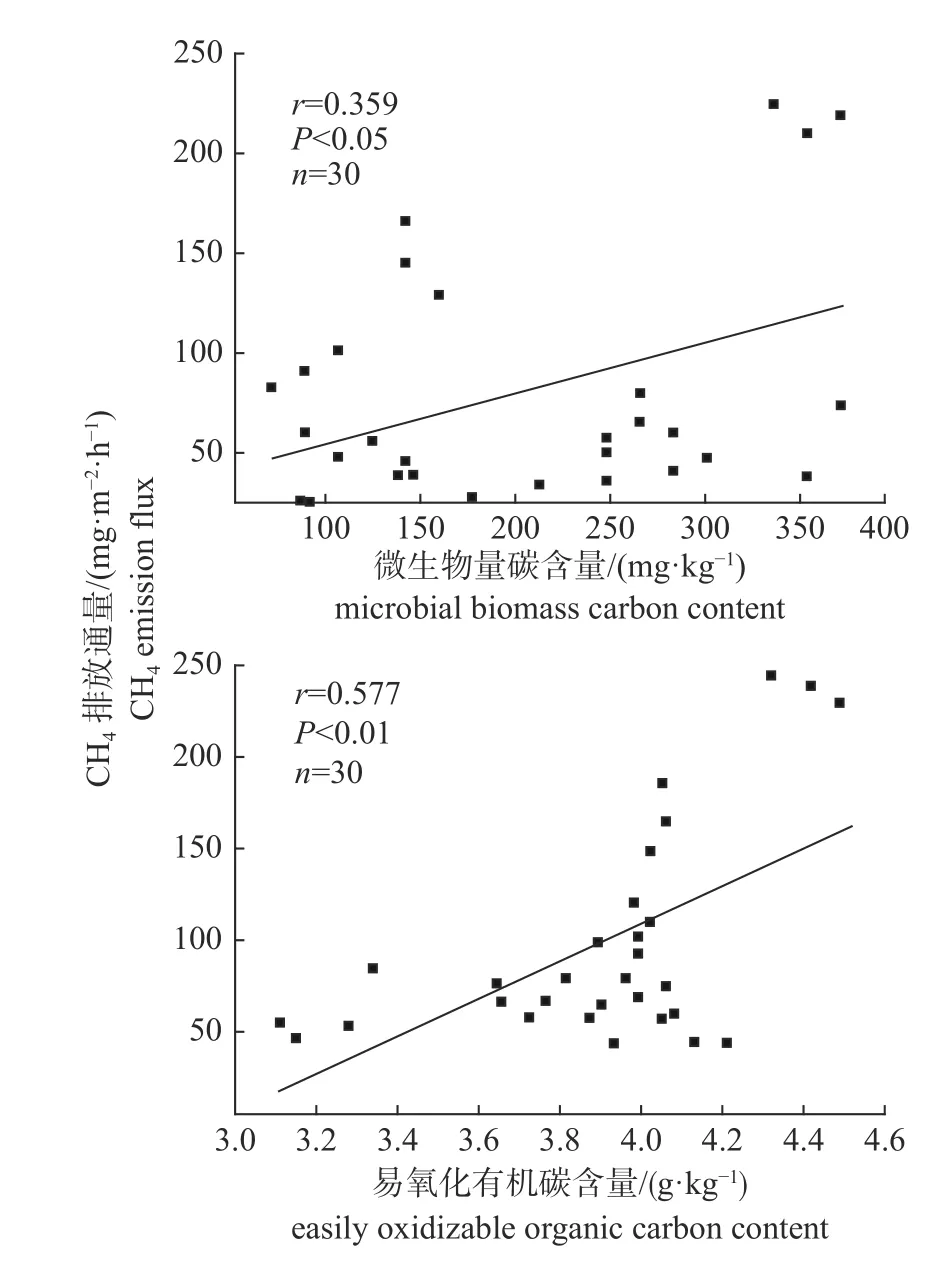
图3 微生物量碳含量和易氧化有机碳含量与CH4 排放通量的相关性)Fig.3 Correlation between microbial biomass carbon content and easily oxidizable organic carbon content and CH4 emission flux
2.3 添加可溶性糖和有机酸对土壤酶活性的影响
由图4 可知:添加不同可溶性糖和有机酸组合对土壤酶活性的影响呈现不同的变化趋势。与CK 处理相比,S、OSM、Sr、SrO、SrS 和SrOSM处理的脱氢酶活性显著增加,OSM 处理增幅高达374%;O、M 和SrM 处理的脱氢酶活性则显著降低;各处理过氧化氢酶活性均显著降低,降幅为14.7%~45.3%,其中,O、M、Sr 和SrM 处理的降幅较小;S、M、Sr、SrM 和SrOSM 处理的蔗糖酶活性显著降低,降幅为38.7%~50.5%,而SrS 处理的蔗糖酶活性则显著增加;S、M、OSM、Sr、SrO 和SrOSM 处理的纤维素酶活性显著降低,降幅为31.2%~40.5%;O、S、OSM、SrO、SrS 和SrOSM 处理的多酚氧化酶活性显著增强,O 和OSM 处理增量高达6.53 和6.66 U/g。
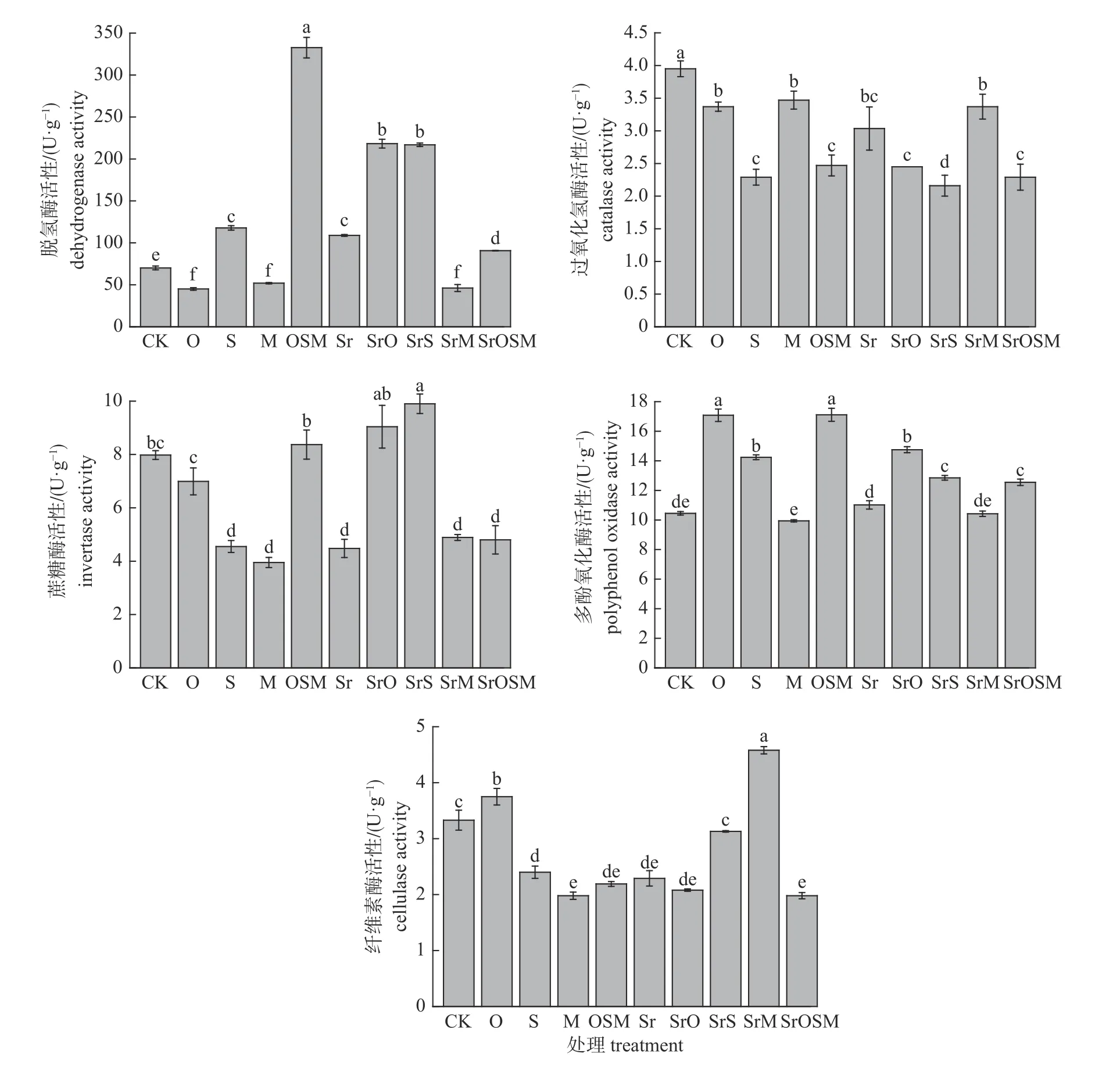
图4 添加可溶性糖和有机酸对土壤酶活性的影响)Fig.4 Effect of the addition of soluble sugar and organic acid on the enzyme activity in soil
通过对土壤碳转化酶活性与活性有机碳含量作相关性分析可知(表2):可溶性碳含量与脱氢酶和多酚氧化酶活性呈极显著正相关,与过氧化氢酶活性呈极显著负相关;易氧化有机碳含量与多酚氧化酶活性呈显著负相关;微生物量碳含量与所测5 种酶活性相关性均不明显。
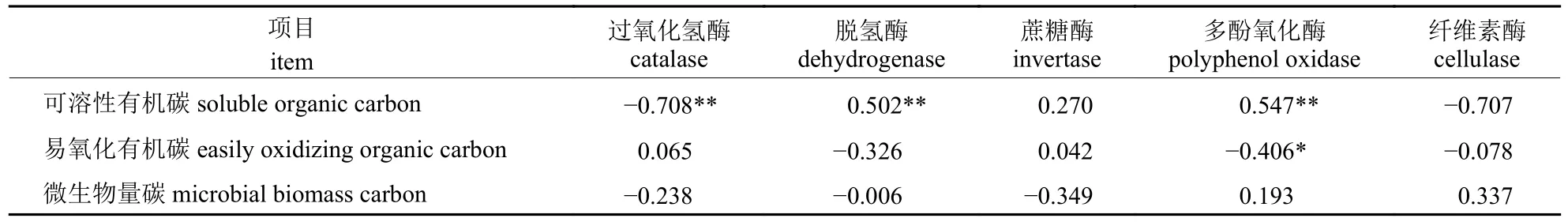
表2 土壤酶活性与活性有机碳含量的相关性Tab.2 Correlation between soil enzyme activity and active organic carbon contents
3 讨论
3.1 添加可溶性糖和有机酸对土壤酶活性的影响
可溶性糖和有机酸是水稻根系重要的根系分泌物,是土壤微生物的主要碳源和氮源,在促进土壤微生物生长的过程中也影响着土壤酶的活性[21]。本研究发现:添加可溶性糖和有机酸后脱氢酶活性显著增加。脱氢酶活性随草酸、苹果酸和琥珀酸质量分数增加呈先降低后增加的趋势,琥珀酸可使酶活性较快地从下降趋势转为上升趋势[22],琥珀酸处理土壤脱氢酶表现为显著增加,本研究设置草酸和苹果酸质量分数未达到拐点仍处于降低阶段,而草酸和苹果酸处理的酶活性显著降低。
添加可溶性糖和有机酸后水稻土壤过氧化氢酶和蔗糖酶活性显著降低,其原因为土壤酶含有大量羧基(-COOH)和氨基(-NH2),低分子量有机酸含有羧基(-COOH)和羟基(-OH),相同的基团会促进有机酸对酶活性的作用,相异基团会减弱该作用;草酸、琥珀酸和苹果酸都含有2 个羧基,苹果酸含有1 个羟基,抑制有机酸作用于酶活性的强度,导致土壤过氧化氢酶和蔗糖酶活性显著降低;可溶性糖含有较多羟基,与不同酸组合后的促进作用是导致SrO 和SrM 处理蔗糖酶活性增加的原因[23]。
水稻土壤多酚氧化酶活性显著提高,有机酸的消耗使体系pH 上升,多酚氧化酶活性在pH 6~9 间呈迅速上升趋势[24];随添加物增加,土壤含水率上升,导致有机质分解过程中间产物多酚的积累,多酚作为多酚氧化酶的催化底物,其含量上升会刺激土壤微生物分泌多酚氧化酶[25];Sr、M 和SrM 处理均含有较多的羟基,抑制添加物作用于酶活性的效果显著,其他处理抑制效果弱,酶活性较强,这与以上结果吻合。纤维素酶主要降解纤维素和木质素等难降解的物质[26],而本试验中添加有机酸和糖类物质并不含有纤维素和木质素等物质,因此导致纤维素酶活性下降。
3.2 添加可溶性糖和有机酸对酶活性与活性有机碳的交互影响
土壤酶是土壤生物与环境间的主要调节者,参与土壤活性有机碳的分解和转化过程,与土壤活性有机碳含量密切相关[27]。本研究中,过氧化氢酶活性与可溶性碳含量呈极显著负相关,脱氢酶和多酚氧化酶活性与可溶性碳含量呈极显著正相关,这是由于可溶性有机碳和易氧化有机碳含量在土壤中周转速度较快、较敏感,能及时反映土壤碳库的变化[28-29]。肖烨等[30]报道土壤易氧化有机碳含量与蔗糖酶活性、纤维素酶活性和过氧化氢酶活性呈显著正相关,与本研究结果存在差异,可能是由于添加可溶性糖和有机酸的组合利用率的不同以及微生物群落结构的改变引起的[11]。多酚氧化酶活性与可溶性有机碳呈极显著正相关,与易氧化有机碳含量呈显著负相关,原因是添加可溶性糖和有机酸改变了根际环境,增加了多酚氧化酶活性,促进土壤有机质的分解以及活性有机碳的转化[15],而本试验中SrO、SrM、O 和M 处理可溶性有机碳含量显著提高,有机酸通过促进微生物活动影响土壤有机质的矿化作用、促进净损失,与YUAN 等[31]的研究一致。试验中直接添加可溶性糖和有机酸,快于水稻根系分泌到环境之中的速度,可溶性有机碳含量显著增加,其为微生物提供充足的底物,微生物量碳含量增加显著。易氧化有机碳含量变化趋势不显著,部分呈降低趋势,相关性分析表明易氧化有机碳与多酚氧化酶呈负相关,多酚氧化酶活性对有机质含量依赖不大[32],其变化趋势可能与其他酶活性共同作用有关,其机理有待深入研究。可溶性糖和有机酸含有的粘胶类物质等,改变土壤的颗粒状态,其有机碳组分比例较高,影响根际土壤的碳组分和含量;此外,可溶性糖和有机酸促进土壤微生物活动和代谢[33],也间接地对土壤碳组分和含量产生复杂影响。
3.3 添加可溶性糖和有机酸对CH4 排放量的影响
可溶性糖和有机酸易被微生物作为碳源利用,分解成CO2、H2和CH4等气体[34]。土壤碳排放能力的重要指标是土壤活性有机碳,尤其是可溶性有机碳、微生物量碳和易氧化有机碳,与CH4的排放密切相关;土壤厌氧微生物将植物根系提供的有机质分解,产物在产甲烷菌的作用下生成CH4[35]。本研究发现:可溶性糖有机酸的输入能够使水稻土壤CH4排放量显著增加,并且这种效应的强弱依赖于可溶性糖和有机酸种类的组成。试验处理中,输入总碳量为0.2 mg/(g·d)时,仅有机酸加入,低分子量有机酸可直接被微生物利用,微生物量碳含量增加显著,土壤酶活性改变,促进CH4排放;输入碳总量为0.3 mg/(g·d)时,可溶性糖的加入使底物进一步增加,导致CH4排放通量和排放总量继续增加[36-37]。由相关性分析可知:CH4排放通量与草酸和苹果酸含量呈正相关,添加草酸和苹果酸处理的微生物量碳含量和可溶性有机碳含量增加,CH4排放量和排放通量显著增加,这是由于草酸和苹果酸结构简单,更容易被微生物利用[38],因此含苹果酸和草酸处理的CH4排放量增加显著;可溶性糖结构复杂,需经分解后才可被利用,与蔗糖酶活性在有可溶性糖组合中增加吻合;添加可溶性糖增加了微生物活动的可利用底物,最终导致SrM 和SrO处理的CH4排放总量显著增加。霍莲杰等[39]研究表明:外源添加可溶性糖和有机酸体系中,易氧化有机碳含量在处理10 d 后趋于稳定,本研究中易氧化有机碳含量表现为差异不显著,有少量处理表现为显著下降,由于添加的可溶性糖和有机酸均是易氧化有机碳,其在土壤中周转快,极易被利用[40],因此,易氧化有机碳被迅速消耗,这与关振寰等[41]的研究结果一致。研究表明:CH4产生途径主要包括有机酸腐解途径和H2-CO2还原途径,其中,有机酸腐解途径产生的CH4占总产量60%~76%[42-43]
相关性分析可知:微生物量碳含量与CH4排放通量呈显著正相关,易氧化有机碳含量与CH4排放通量呈极显著正相关,易氧化有机碳含量受草酸影响显著,微生物量碳含量受苹果酸影响显著,因此CH4排放通量受苹果酸影响显著,草酸次之。易氧化有机碳含量与CH4排放通量呈极显著正相关,原因是在淹水还原环境下,有机物的数量和对厌氧微生物的有效性在CH4形成过程中起着决定性作用[31],虽然微生物量碳含量与可溶性有机碳含量同样对CH4排放产生影响,但较易氧化有机碳含量影响不显著,其原因为CH4产生过程较CO2更为敏感,最终的产生潜力受底物等较多环境因素影响[44]。由此可知,可溶性糖和有机酸的添加影响稻田土壤中有机碳的组分含量,提高稻田土壤中易氧化有机碳和微生物量碳等与CH4排放密切相关的活性有机碳的含量[45],进而影响到CH4的产生,增加CH4的排放量。
水稻根系分泌物释放改变土壤活性有机碳的组分,为土壤CH4产生提供重要底物;根际土壤活性有机碳组分、含量和根际微生物直接影响CH4排放[46]。根系分泌物排放增加,土壤中活性物质增加,进而影响微生物活性,促进CH4排放[47]。土壤有机碳具有的巨大库容,其微小变化就会影响到碳向大气的排放速率,进而影响大气中温室气体浓度[48]。因此,土壤碳库动态变化的研究在全球的气候变化中具有重要地位。
4 结论
添加可溶性糖和有机酸显著增加微生物量碳和可溶性有机碳含量,显著增加多酚氧化酶和脱氢酶活性,降低蔗糖酶、纤维素酶和过氧化氢酶活性。苹果酸处理对CH4排放的促进效果显著,草酸次之。添加可溶性糖和有机酸通过改变土壤碳含量和酶活性,进而影响CH4排放量和排放通量。
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26
中成药(2018年5期)2018-06-06
分析仪器(2016年2期)2016-09-07
广东石油化工学院学报(2016年6期)2016-05-17
中国粮油学报(2016年5期)2016-01-23
中国茶叶加工(2015年3期)2015-02-27
茶叶通讯(2014年2期)2014-02-27
河南城建学院学报(2014年4期)2014-02-27
食品科学(2013年15期)2013-03-11
中国果业信息(2013年12期)2013-01-22
