
半干旱草地不同植物枯落物分解对放牧和封育的响应
2021-09-01 10:55刘任涛张安宁蒋嘉瑜唐希明
生态学报 2021年14期
陈 蔚,刘任涛,*, 张安宁, 蒋嘉瑜, 唐希明
1 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,银川 750021 2 中卫市自然资源局,中卫 755000
在半干旱区,放牧作为草地生态系统主要的人类干扰方式,易导致草地生态系统发生极度退化和沙漠化,而封育管理是退化草地生态系统重要的生态恢复措施,能够有效促进退化草地生态系统结构恢复[1-2]。枯落物分解是调控地上-地下生态系统物质循环与能量流动的重要环节[3],是深刻理解生态系统结构与功能、物质循环规律的重要途径[4]。在封育管理条件下,通过改变植物枯落物组成[5]、土壤生物活动和生物活性[6]及非生物环境[7]来直接或间接影响枯落物分解,进而影响到退化生态系统的恢复进程。所以,研究封育管理对枯落物分解过程的影响规律,对于退化生态系统恢复、物质循环过程解析及采取有效的草地管理措施均具有重要意义。
目前,关于草地植物枯落物分解的研究,主要集中在环境和生物因素对枯落物分解的影响及养分释放动态等方面。Porazinska等[8]研究表明,水分不足会严重影响枯落物的分解,而干旱条件往往会降低分解率。Bradgett等[9]研究表明,节肢动物与微生物的共同作用有助于枯落物的分解。并且,欧阳林梅等[10]研究发现,枯落物的基质质量与其分解速率均具有显著相关性。但Song等[11]研究表明,不同功能群的植物共存会加速根叶枯落物的分解,从而促进草地营养循环。综合分析表明,草地植物枯落物分解过程与枯落物基质质量、生物活性和非生物环境条件密切相关。但是,目前关于草地生态系统中放牧和封育管理条件下不同植物组成的枯落物分解过程研究,报道较少。
国内外研究表明,放牧主要通过食草动物行为直接和间接影响枯落物分解。放牧活动可以减少高质量的枯落物量[12]、加速枯落物破碎[13]和影响土壤动物分布[14],亦会对枯落物分解的无机环境因子包括太阳辐射[15]、水热条件[16]等产生深刻影响。但在封育条件下,植被覆盖度和多样性提高,土壤食物网复杂,土壤动物数量增加[17],枯落物的分解过程可能发生改变。所以,通过比较荒漠草原优势植物枯落物对放牧和封育管理的响应规律,有助于揭示放牧管理对于半干旱草地生物地球化学循环的内在影响机制。并且,利用网孔分解袋法来完全或部分限制土壤动物进入,可以深入阐明放牧和封育条件下枯落物分解过程中土壤动物对枯落物分解的生态作用。
鉴于此,本研究以宁夏半干旱不同植物枯落物作为研究对象,采用网孔分解袋法研究放牧和封育条件下不同植物枯落物分解率变化特征及土壤动物对枯落物分解的贡献情况,旨在揭示半干旱区植物枯落物分解对放牧与封育管理的响应规律,为半干旱区脆弱草地生态系统生物地球化学循环过程解析、放牧管理与植被恢复提供科学依据。
1 研究区概况
本研究样地位于宁夏盐池县东北部(37°49′ N,107°30′ E,海拔1348 m)。该区域属中温带大陆性季风气候。年平均气温为8.5℃,最低平均气温-8.7℃(1月),最高平均气温22.4℃(7月)。多年平均降水量281.7 mm,7—9月降水量占总降水量的60%。年实际蒸散2136 mm,年潜在蒸散1120 mm。主要的地带性土壤类型有灰钙土和黄绵土;非地带性土壤类型为风沙土,土壤肥力低,结构疏松,易受风蚀侵蚀。在过度放牧条件下,草地几无植被覆盖,易发生退化、沙化。
本研究样地包括相邻的放牧和封育2种类型草地。放牧草地:通常在每年的6—8月进行季节性羊群放牧,放牧强度为重度放牧(11只/hm2)。放牧草地植被主要有赖草(L.secalinus)、猪毛菜(Salsolacollina)和白草(Pennisetumcentrasiaticum)。生长季前的植被覆盖率<5%,生长季后约20%。
封育草地:于1993年,采取刺栅栏的围栏管理措施,目前已封育了23年,主要植被主要包括猪毛蒿(Artemisiascoparia)、山苦荬(Ixerischinensis)、牛枝子(L.potaninii)、草木樨状黄芪(Astragalusmelilotoides)。生长季前植被覆盖率为20%,生长季后约为60%。
2 研究方法
2.1 枯落物样品采集与布设
于2017年10月,在放牧和封育草地中,采用样方法(50 cm×50 cm),用剪刀齐地面采集赖草和牛枝子2种优势植物,带回实验室置于65℃下烘干至质量恒定,以调查自然条件下赖草和牛枝子地上枯落物生物量分布特征及二者混合物比例。然后,依据分解袋规格(10 cm×10 cm)、样方面积以及枯落物地上生物量分布特征,得出分解袋中枯落物质量为10 g。
将上述烘干后的枯落物剪短至5 cm左右,将赖草、牛枝子(牛枝子为枝和叶均匀的混合物,保证分解袋中比例一致)及二者混合物(4:3)分别称量10 g,装入网孔大小为30目和250目2种型号的网孔分解袋中[18](30目网孔允许全部土壤动物进入参与枯落物分解,250目网孔仅允许微生物进入参与枯落物分解),为了防止枯落物损失,贴近地表的枯落物网孔为250目。每种网孔分解袋有3个重复。
首先,在各个分解袋内放入刻有编码的标签,做好记录。然后,将分解袋按9行×15列方式平铺至研究样地中,枯落物网袋之间的间隔为10 cm,以保证分解袋所在环境条件基本一致。2个样地总共铺设分解袋180个。自放分解袋之日(2017年9月24日起),分别于非生长季(2018年5月21日和2019年5月18日)、生长季(2018年9月28日和2019年9月28日)进行取样,取样5次。共取样180袋(3种处理×3个重复×2种网孔×2种生境×5次取样)。
2.2 枯落物样品收集与分析
每次取分解袋时,按照同样的顺序各从放牧、封育样地中取回样品36袋(3种处理×3个重复×2种网孔×2种生境)。具体取样措施为:首先用毛笔小心刷除粘附在分解袋表面的沙土,将枯落物分解袋装入信封,保存在恒温箱内,带回实验室;然后取出分解袋中枯落物样品,去除沙粒等杂质后置于65℃下烘干至恒重,记录干重;最后,将样品粉碎用于测定灰分含量。
本实验中,枯落物全碳(%)采用重铬酸钾-硫酸氧化法测定[19],枯落物营养元素中的全氮(%)采用凯氏定氮法测定[19],全磷(%)采用钼锑抗比色法测定[19],木质素(%)采用由Van Soest改进的范式法进行测定[20]。
2.3 土壤样品采集与分析
在试验结束(2019年9月下旬)于样地采集土壤样品进行环境特征的测定。首先,在分解袋贴地表处采用地温计测定土壤温度(℃)。然后,用铝盒取1/3的土壤样品(0—10 cm),以测定土壤含水量;最后,用小铲将其余2/3的土壤样品(0—10 cm)取回放在自封袋中带回实验室。自然风干后的土壤过2 mm土壤筛,去除树根等杂质,以测定土壤pH、电导率和粒径;将土壤研磨后过0.01 mm筛,以测定土壤全碳(TC)和土壤全氮(TN)。
土壤含水量(%)采用烘干称重法进行测定。土壤pH(2.5∶1水土比悬液)和电导率(5∶1水土比浸提液;μs/cm)分别采用酸度计法和电极法进行测定。土壤全碳(%)采用重铬酸钾-硫酸氧化法测定[19],全氮(%)采用凯氏定氮法测定(表1)[19]。土壤粒径采用Mastersizer 3000激光衍射粒度分析仪测定。

表1 放牧和封育草地土壤特征
2.4 数据处理与统计分析
枯落物的质量损失率(Lm)及土壤动物对枯落物质量损失的贡献率(Pfau)计算公式[21-22]如下:
式中,M0为分解袋埋置前的烘干枯落物质量;Mt为干枯落物在t时刻的残留率;(ML30-ML250)为最近一次采样30孔径与250孔径分解袋质量损失率的差值;(MLtotal)为在最终采样时从30孔径中获得的枯落物质量损失率。文中枯落物最终质量残留率Rm即为(1-MLm)。
在分析之前,对所有重复的测量值和采样时间进行平均。然后通过拟合一个单独的混合指数衰减函数来估计衰减常数[23](k值)。其中,k值越大,枯落物分解速率越快[24]:
Mt=M0e-kt
式中,Mt为枯落物在t时刻的质量(g),M0为初始质量(g),t为分解时间,k为枯落物分解常数。
为了评价封育、放牧对枯落物类型分解的广义效应,采用双向检验方法分别检验了枯落物的衰减常数(k)和最终残留率(%)方差分析。采用单因素方差检验分解时间下网孔处理与物种差异对枯落物分解的影响,从基质质量指标中挑选最优子集进行多元线性混合模型拟合。采用Pearson相关性系数检验枯落物分解与基质质量之间的关系。固定效应包括放牧和生境处理,网孔和枯落物类型作为随机效应嵌套在区间内。其交互作用和主要效应的显著性被设置为α=0.05。在进行参数分析之前,所有数据都进行了正态性检验。在Origin 2018和R 3.6.0软件中进行作图。
3 结果与分析
3.1 植物枯落物基质质量
从表2可以看出,植物C含量表现为3种枯落物间无显著差异(P>0.05)。植物N含量表现为赖草>混合物>牛枝子(P<0.05)。植物P含量则表现为赖草和混合物显著高于牛枝子(P<0.05),而前者无显著差异(P>0.05)。植物木质素含量表现为牛枝子>混合物>赖草(P<0.05)。
由表2可知,植物C/N和木质素/N均表现为牛枝子>混合物>赖草(P<0.05)。植物C/P和N/P则表现为牛枝子显著高于赖草,且均与混合物无显著差异(P>0.05)。

表2 植物枯落物基质初始含量及初始含量比
3.2 基于网孔分解袋的放牧和封育生境中植物枯落物分解特征
通过多因素方差分析(表3),在相同的分解时间内,30目和250目网孔仅对牛枝子枯落物的最终质量残留率(Rm)产生显著影响(F=6.89,P=0.030),而放牧和封育生境对牛枝子枯落物最终质量残留率以及分解衰减常数(k)均产生显著影响(FRm=6.15,P=0.038;Fk=20.35,P=0.002)。
不同网孔处理下放牧和封育生境中枯落物Rm和k均存在显著差异(图1)。放牧样地中,赖草和混合枯落物Rm与k均表现为不同网孔间无显著差异,而牛枝子枯落物Rm表现为30目显著低于250目网孔(P<0.05),k表现为30目显著高于250目网孔(P<0.05)。封育样地中,3种枯落物中Rm均表现为不同网孔间无显著差异,而牛枝子枯落物中k表现为30目显著高于250目网孔(P<0.05),赖草和混合物中均无显著差异。
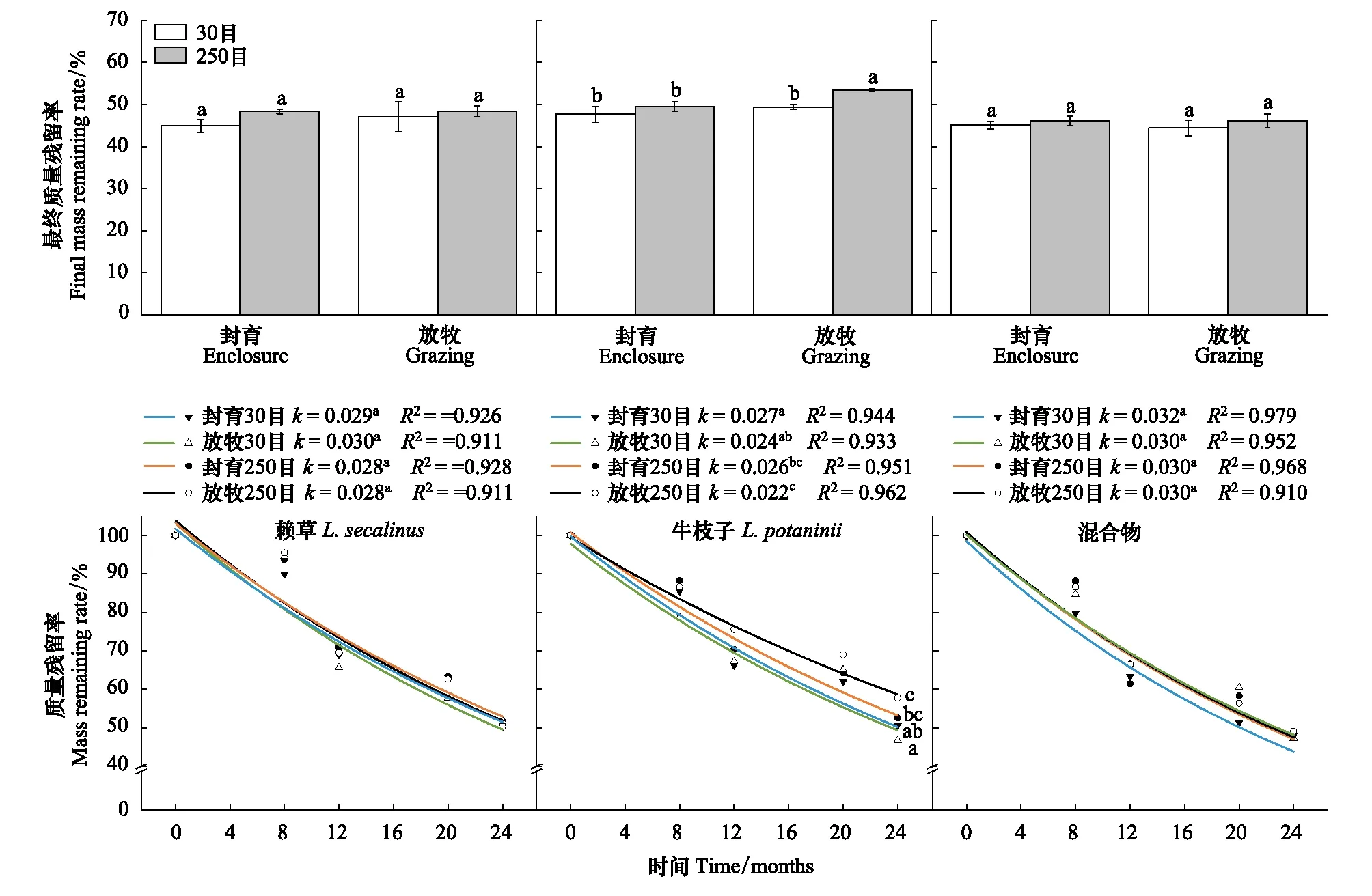
图1 放牧和封育生境下不同网孔枯落物袋Rm和kFig.1 Rm and k of plant litter from different mesh-size litterbags in grasslands under grazing and enclosure management不同小写字母表示不同生境不同网孔枯落物之间差异显著(P<0.05)。Rm 最终质量残留率 Final mass remaining rate;k 分解衰减常数 Decomposition decay constant

表3 生境、网孔和物种对枯落物k值和Rm影响的多因素方差分析
在放牧样地,Pfau表现为牛枝子枯落物显著高于混合物,赖草居中(P<0.05)。而在封育样地,土壤动物贡献率(Pfau)表现为赖草显著低于牛枝子和混合枯落物,而后两者间无显著差异(表4);3种类型的枯落物中,赖草和牛枝子枯落物中的Pfau表现为放牧和封育样地间均无显著差异,而混合枯落物的Pfau则表现为放牧显著低于封育(P<0.05)。
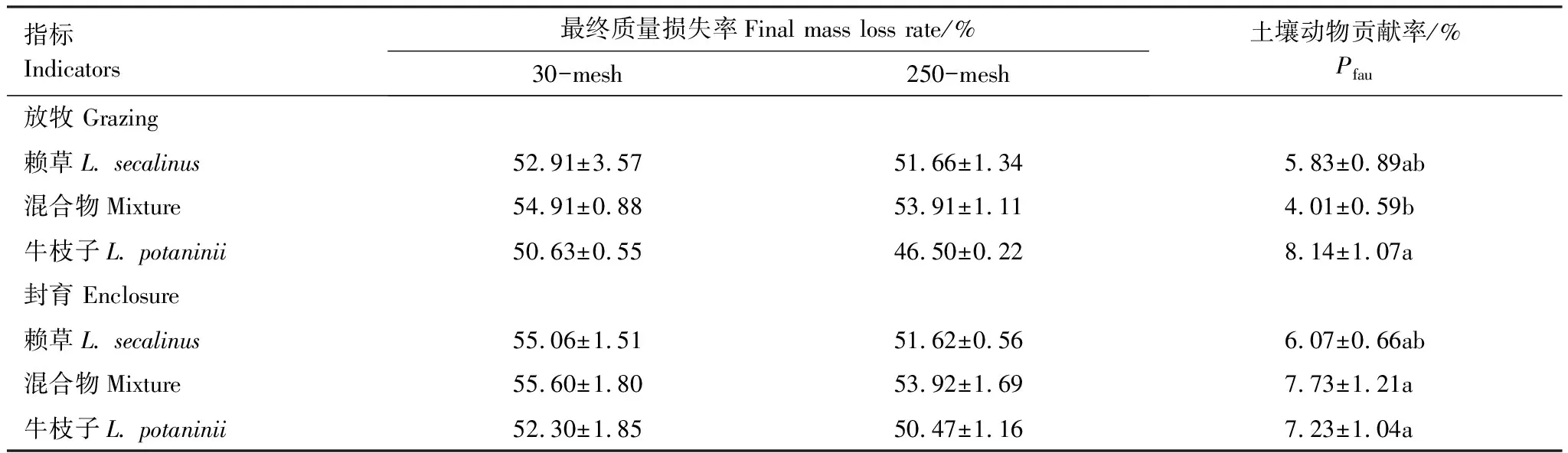
表4 放牧和封育生境下3种枯落物中土壤动物贡献率
3.3 基于物种组成的放牧和封育生境中植物枯落物分解特征
通过多因素方差分析(表3),在相同的分解时间内,枯落物种类对不同网孔下植物枯落物k的影响均显著(F30目=5.14,P=0.024;F250目=15.75,P<0.001),且对250目网孔中枯落物Rm影响显著(F=11.73,P=0.002)。但放牧和封育生境对不同网孔下枯落物的k以及Rm的影响均不显著。放牧和封育生境与枯落物种类的交互作用仅对250目网孔中枯落物的k的影响显著。
不同物种处理下放牧和封育生境中的Rm和k均存在显著差异。250目网孔中,放牧样地的枯落物Rm表现为牛枝子显著高于赖草和混合物,而后两者间无显著差异(图2;P<0.05);而封育和放牧样地中k均表现为混合物显著高于牛枝子,赖草居中,且牛枝子枯落物k表现为封育样地显著高于放牧样地(图2;P<0.05)。而在30目网孔中,封育和放牧生境下不同枯落物Rm均无显著差异(图2;P>0.05),k表现为封育和放牧样地的赖草和混合物显著高于牛枝子枯落物,而前两者间无显著差异,且牛枝子枯落物k表现为封育样地显著高于放牧样地(图2;P<0.05)。
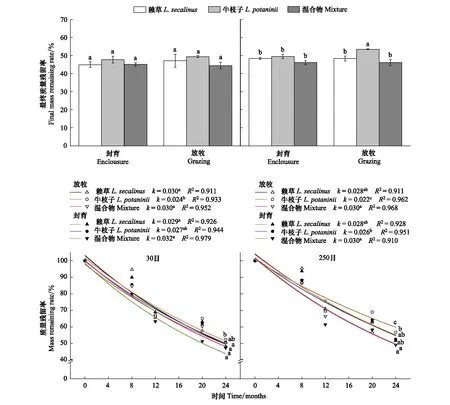
图2 放牧和封育生境下不同物种枯落物Rm和kFig.2 Rm and k of three plant litter under grazing and enclosure management不同小写字母表示不同物种枯落物之间差异显著(P<0.05)
3.4 植物枯落物分解特征与枯落物初始质量的关系
由图3可以发现,枯落物Rm与枯落物N、P含量呈负相关,而与木质素/N呈正相关(P<0.05),且与枯落物C/P呈显著正相关(P<0.01)。枯落物k与枯落物N/P呈负相关(P<0.05),与木质素含量呈显著负相关(P<0.01),与木质素/N、C/N、C/P均呈极显著负相关(P<0.001),而与N、P含量呈极显著正相关(P<0.001)。
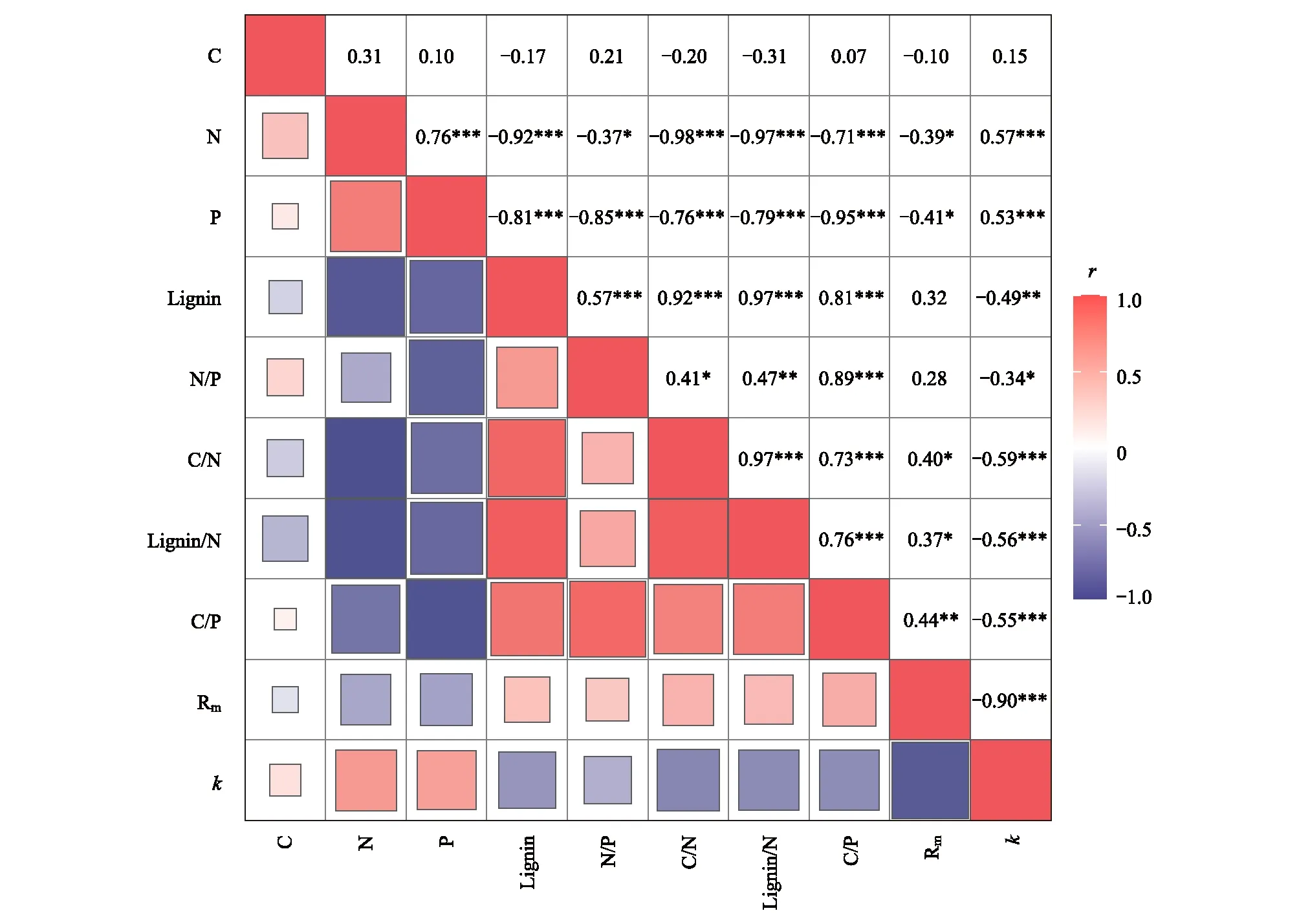
图3 植物枯落物分解与初始基质质量之间的相关性分析Fig.3 Correlation analysis between the litter decomposition and initial litter qualityC 初始全碳含量 Initial total carbon content;N 初始全氮含量Initial total nitrogen content;P 初始全磷含量Initial total phosphorus content;Lignin 初始木质素含量 Initial lignin content;N/P 初始氮磷比 Initial nitrogen phosphorus ratio;C/N 初始碳氮比 Initial carbon nitrogen ratio;Lignin/N 初始木质素氮比 Initial lignin nitrogen ratio;C/P 初始碳磷比 Initial carbon phosphorus ratio。*P<0.05;**P<0.01; ***P<0.001
通过多元线性模型拟合发现(图4),混合效应下枯落物分解最终质量残留率拟合度较低(截距=0.26),而分解衰减常数具有较高的拟合度(截距=0.0005)。同时,在枯落物残留率的拟合模型中,所有数据点的56%在1:1直线的左侧,仅44%的点在右侧。在分解衰减常数的拟合模型中,所有数据点的58%在1∶1直线的左侧,仅42%的点在右侧。

图4 放牧和封育处理下枯落物Rm和k的多元混合线性模型Fig.4 Multivariate mixed linear model of Rm and k of plant litter under grazing and enclosure management红色实线表示预测值与观测值的线性拟合方程。虚线表示1∶1下的直线
4 讨论
4.1 基于网孔分解袋的封育和放牧管理对植物枯落物分解的影响
枯落物分解袋法是目前研究不同体型土壤动物对枯落物分解影响普遍采用的方法。本研究中,网孔处理对3种枯落物分解过程无显著影响(表3),这与图1研究结果一致。表明在半干旱地区土壤动物对枯落物分解作用有限,不足以影响整个枯落物分解过程[25]。例如,在冬季土壤动物如双翅目、鳞翅目和鞘翅目等进入冬眠,对于枯落物分解作用较小[26]。另外,土壤动物的破碎作用往往优先,与之相比体型更小的微生物则通过产生相关分解酶(蛋白质酶、木质素降解酶等)直接有效的提高枯落物分解率[27]。总得来说,土壤动物的存在一定程度上影响枯落物质量损失,但并未影响分解的总体变化趋势。这与Frouz等[28]的研究一致。而网孔处理对牛枝子枯落物最终质量残留率存在显著影响,研究发现牛枝子的土壤动物贡献率较高(图1,表4),这可能是牛枝子较难分解(木质素含量高达19.30),其对土壤动物破碎的需求更大,且最终取样时间在夏季,处于土壤动物活动期[29],这使得土壤动物在这一时期对牛枝子枯落物影响较大。
研究发现,放牧和封育管理对于赖草和混合枯落物分解的影响并不显著,但显著影响牛枝子枯落物分解(表3)。本研究中,牛枝子枯落物中k表现为封育样地显著高于放牧样地(图1)。这表明相较于放牧管理,封育促进了牛枝子枯落物的分解。这与众多研究[30-31]相悖,放牧管理一方面通过践踏效应促进了枯落物的破碎,另一方面牲畜的粪尿排泄为土壤动物(特别是腐食性和粪食性)的繁殖和定居提供良好的条件,从而有利于枯落物分解。但这与Shariff等[32]的研究相一致。主要原因在于区域尺度上,温度、湿度等局部尺度的环境因子对枯落物分解具有深刻的影响[4,33]。可以发现,封育样地中植被的恢复导致地表覆盖度增加,土壤温度显著降低,土壤水分有效性显著增加(表1)。而肖玲等[34]的研究发现,土壤水分作为半干旱区土壤生物生存的限制性因子,不仅会影响枯落物的分解过程,同样深刻影响土壤生物多样性,这一定程度上有利于封育对枯落物分解的促进作用。放牧活动会造成土壤微生境极不稳定,而在封育管理下植被恢复所形成的更加稳定的立地条件,往往更适合枯落物中土壤动物生存和定殖[14]。
通过枯落物分解袋法可以有效量化土壤动物在枯落物分解中的贡献,进而深入了解放牧和封育管理下土壤动物在不同种枯落物分解中的作用。本研究中,3种枯落物的土壤动物贡献率均大于0,这表明土壤动物对半干旱区枯落物的分解具有促进作用[35]。在研究样地的前期研究结果发现,蜱螨目作为半干旱区枯落物分解主要的土壤节肢动物类群,其中一部分植食性或腐食性类群通过采食和破碎的形式促进枯落物的分解[36],一部分食真菌或是细菌类群通过选择性取食可以有效抑制微生物之间的竞争,从而具有更高的微生物多样性和更有效的分解能力[37]。研究发现,分解较慢的牛枝子枯落物中土壤动物贡献率在封育(Pfau=7.23)和放牧(Pfau=8.14)样地中较高。这与Gergocs等[37]的研究一致,相对于优质枯落物,中小型土壤动物对难降解枯落物的促进作用往往更强,这也进一步说明微生物对于枯落物(尤其是难降解的)的分解作用需要建立在土壤动物破碎作用的基础上。且放牧条件下增加了践踏破碎作用,使得这种作用更加显著,这也解释了放牧和封育样地中牛枝子枯落物在30目网孔的分解作用显著高于250目。本次研究结果显著低于武海涛等[38]在三江平原湿地的研究结果,原因可能在于半干旱区独特的环境条件如较高的紫外线辐射和较低的土壤含水量限制了中小型土壤动物的数量和多样性[39]。研究中,封育样地中混合枯落物土壤动物贡献率显著高于放牧样地,但其枯落物分解并未出现显著差异。可能的原因在于虽然放牧干扰一定程度上降低了混合物中土壤动物多样性,但捕食者的减少和食草动物的排泄行为增加了土壤中微生物多样性,从而促进了混合枯落物的分解[40]。同样在Bonanomi等[41]的研究发现,混合枯落物相比于单一枯落物,具有更广泛的分解者和更密集的食物网结构,这一定程度上有效抵御了放牧干扰所造成的影响,从而导致放牧和封育生境下枯落物分解未出现显著差异。而赖草和牛枝子枯落物在不同生境中的土壤动物贡献率均无显著差异,这表明封育和放牧并不能直接通过影响赖草和牛枝子枯落物中土壤动物来影响枯落物分解。研究表明,封育和放牧管理下,土壤动物深刻影响混合枯落物的分解,且对低质量枯落物(牛枝子)的分解效果更强。
4.2 基于物种组成差异的封育和放牧管理对植物枯落物分解的影响
众多研究表明,基质特性往往是影响小尺度的枯落物分解的决定性因素,初始基质特性是不同种枯落物化学属性的重要体现[42]。本研究中,不同种枯落物在250目和30目网孔分解下均存在显著差异(表3),表明物种差异显著影响半干旱区优势枯落物的分解,这与赵娟等[43]的研究一致。30目和250目网孔中,赖草和混合物在封育和放牧样地间Rm和k均无显著差异,而牛枝子枯落物的k均显著低于赖草和混合物(图2)。原因在于牛枝子枯落物的初始P和C/N含量显著低于赖草和牛枝子。初始P、C/P含量显著影响枯落物的分解(图3),足够的P含量能够为枯落物分解过程中的微生物提供大量的三磷酸腺甙(ATP),有利于微生物进行代谢活动,加速枯落物分解[44]。另外,Taylor等[45]研究显示,枯落物中木质素含量>20%时,C/N可以作为枯落物分解速率的理想指标。其中,牛枝子枯落物的C/N>30时,表明N将成为土壤有机质中的限制因素,此时C/N增加会限制牛枝子枯落物的分解,从而导致不同网孔下禾本科的赖草枯落物分解速率显著高于豆科的牛枝子。另外,混合枯落物与赖草的Rm无显著差异,但其k要高于赖草枯落物,主要原因在于枯落物自身理化性质可以通过在枯落物类型之间转移营养物质,或通过影响土壤生物活性来间接影响整体分解率和混合物中各组分枯落物的衰变率,由此混合枯落物在分解过程中会表现出不同组分间的协同或者拮抗作用,且研究显示大多数混合枯落物分解会产生一种“非加性”协同效应[46]。本研究中,混合枯落物中存在一种协同的非加性混合效应。
本研究中,枯落物中k与初始N含量存在显著正相关关系,而与初始N/P、木质素/N和Rm均存在显著负相关关系(图3),这与Berg等[47]的研究一致,枯落物分解过程中微生物需要利用N进行矿化作用[48]。而枯落物残留率越高,会抑制枯落物的分解过程,从而表现为Rm与k呈极显著负相关关系。研究表明,半干旱区不同枯落物分解具有显著差异,基质质量显著影响枯落物的分解,且放牧管理对不同物种枯落物分解的影响并不显著。有研究表明,在小尺度的枯落物分解过程中,枯落物基质质量往往是枯落物分解的决定性因素[42]。且土壤动物对枯落物分解的作用取决于枯落物的质量,往往枯落物质量不仅仅由一种元素或元素比影响,而是由多种元素共同作用[10]。本研究中,通过多元线性混合模型拟合发现预测值和实际测量值具有较高的拟合度,其中,k的拟合方程截距较小,拟合程度更高。表明枯落物分解过程中,k受到初始基质质量的共同作用的影响更大,这与图3中的Pearson相关性结果相一致。另外,拟合模型发现Rm和k的数据测量值均高于试验预测值。可能的原因在于还有其他因素如淋溶作用[49]、光照[50]的影响,这从Rm和k的拟合决定系数较小可以看到(R2=0.43;R2=0.52)。可以看到,枯落物分解受到多种元素的作用,且无机环境与生物的协同作用促进枯落物的分解。
5 结论
(1)放牧和封育管理对赖草和混合枯落物分解的影响并不显著,但封育促进了牛枝子枯落物分解。(2)土壤动物有效调控混合枯落物的分解,且对低质量枯落物(牛枝子)的分解效果更强。(3)无论是在放牧还是封育管理下,植物物种差异显著影响枯落物的分解,赖草比牛枝子枯落物分解速率更快,而混合枯落物通过具有协同作用的“非加性”效应可以加速分解。研究表明,放牧与封育管理通过影响网孔中土壤动物分布来调控半干旱区草地植物枯落物的分解,封育能够有效促进半干旱草地低质量枯落物(牛枝子)的分解。
猜你喜欢
铁道科学与工程学报(2022年10期)2022-11-30
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
毛纺科技(2021年8期)2021-10-14
绿色科技(2021年10期)2021-06-23
林业科技(2020年3期)2021-01-21
模具制造(2020年6期)2020-08-03
铁道标准设计(2020年4期)2020-04-26
民间故事选刊·下(2020年2期)2020-03-18
幸福·婚姻版(2017年1期)2017-01-24
