
蚊传呼肠孤病毒研究进展
2021-08-31 08:24郭晓芳
中国人兽共患病学报 2021年8期
田 杰,郭晓芳
虫媒病毒(Arbovirus)是指一些通过吸血节肢动物叮咬传播引起人或动物发病的病毒,其中有130余种可引起人兽共患疾病,主要分布于披膜病毒科、黄病毒科、布尼亚病毒目和呼肠孤病毒科,感染后表现为发热、皮疹、关节痛、出血热和脑炎等症状[1-2]。呼肠孤病毒科(Reoviridae)由9~12个基因组片段的双链RNA病毒组成,其病毒呈等轴二十面体球形结构,有1~3层衣壳包裹,直径为60~80 nm,目前仅发现Aedes pseudoscutellaris reovirus(APRV)为9节段并单衣壳病毒[3-4]。根据核结构不同可将呼肠孤病毒科分为无刺突呼肠孤病毒亚科(Sedoreovirinae)和刺突呼肠孤病毒亚科(Spinareovirinae,又称为“炮塔型”呼肠孤病毒[4])。根据国际病毒分类委员会(International Committee for Taxonomy of Virus, ICTV)2019年分类(https://talk.ictvonline.org/taxonomy/),现有9属58种刺突呼肠孤病毒,无刺突呼肠孤病毒亚科包括6属39种(见表1),包括未列入分类的共15属[5-6]150余种[1]。

表1 呼肠孤病毒分类
呼肠孤病毒科病毒分布广,其宿主小到真菌大到哺乳动物[4],蚊虫作为其重要的传播媒介,分布范围最广,繁殖和传播病毒能力最强。自新中国成立以来,我国从库蚊属、按蚊属、阿蚊属、伊蚊属等蚊虫中相继分离到10多种以往未分离到的病毒,其中呼肠孤病毒科包括版纳病毒(Bannavirus, BAV)、辽宁病毒(Liaoning virus, LNV)、Kadipirovirus(KDV)、云南环状病毒(Yunnan orbivrus, YUOV)、西藏环状病毒(Tibet orivirus, TIBOV)和芒市病毒(Mangshi virus)[7]。BAV作为我国于世界首次发现的呼肠孤病毒科新属代表株,在国际上引起了极大的关注,但自首次分离后鲜有报道。目前已知蚊传呼肠孤病毒中确认对人致病的有YUOV[8]和BAV[9],其余则对哺乳动物、禽和鱼类致病,人感染后可表现为中枢神经系统疾病、呼吸道及肺部感染以及胃肠疾病[10]。本文对蚊传呼肠孤病毒科的研究现况进行综述,旨在了解其分布情况,及对人和动物致病情况,为我国蚊传呼肠孤病毒病防治措施的制定提供依据。本文中列出的蚊传呼肠孤病毒及分布地区和媒介蚊类见表2。

表2 蚊传呼肠孤病毒种类及媒介蚊类
1 无刺突呼肠孤病毒亚科(Sedoreovirinae)
1.1 Orbivirus(环状病毒属) 环状病毒属是1971年由Borden等命名的以吸血节肢动物作为传播媒介,在脊椎动物与节肢动物体内繁殖,引起人兽共患病的虫媒病毒属[11-12]。环状病毒属的病毒均为10节段dsRNA病毒,双层衣壳呈20面体对称结构,核心含32个环状粒子,编码7种结构多肽(VP1-7)和4种非结构多肽(NS1-4)[5,11],NS4在VP6段中编码[6]。VP1(Pol)、T2和VP7(T13)是环状病毒最保守的序列,常用于环状病毒属病毒的鉴定[13-14],OC1(VP2/3)和OC2(VP5)高度可变,常用于环状病毒属内部血清型鉴定[15]。通常VP2编码T2蛋白的病毒由蚊和蜱传播,VP3编码T2蛋白由蠓传播[15-16],但是新发现的一些VP3编码T2蛋白的病毒多数从蚊中分离。
据ICTV记录现有22种环状病毒,另有15种未记录[17]。近年来全球通过双链RNA病毒的全长cDNA扩增技术与高通量测序鉴定出了多种新型蚊传环状病毒:尤马蒂拉病毒(Umatilla virus,UMAV)、科里帕塔病毒(Corriparta virus, CORV)及其新亚型California mosquito pool virus(CMPV)、Sathuvachari virus(SVIV)、Kammavanpet virus(KVPTV)[6, 16]、秘鲁马病病毒(Peruvian horse sickness virus,PHSV)、Mobuck virus、云南环状病毒(Yunnan orbivirus,YUOV)及其新亚型西藏环状病毒(Xizang orbivirus,TIBOV)和广西环状病毒(Guangxi orbivirus,GXOV)[18]。各蚊传环状病毒的基因组特征见表3。
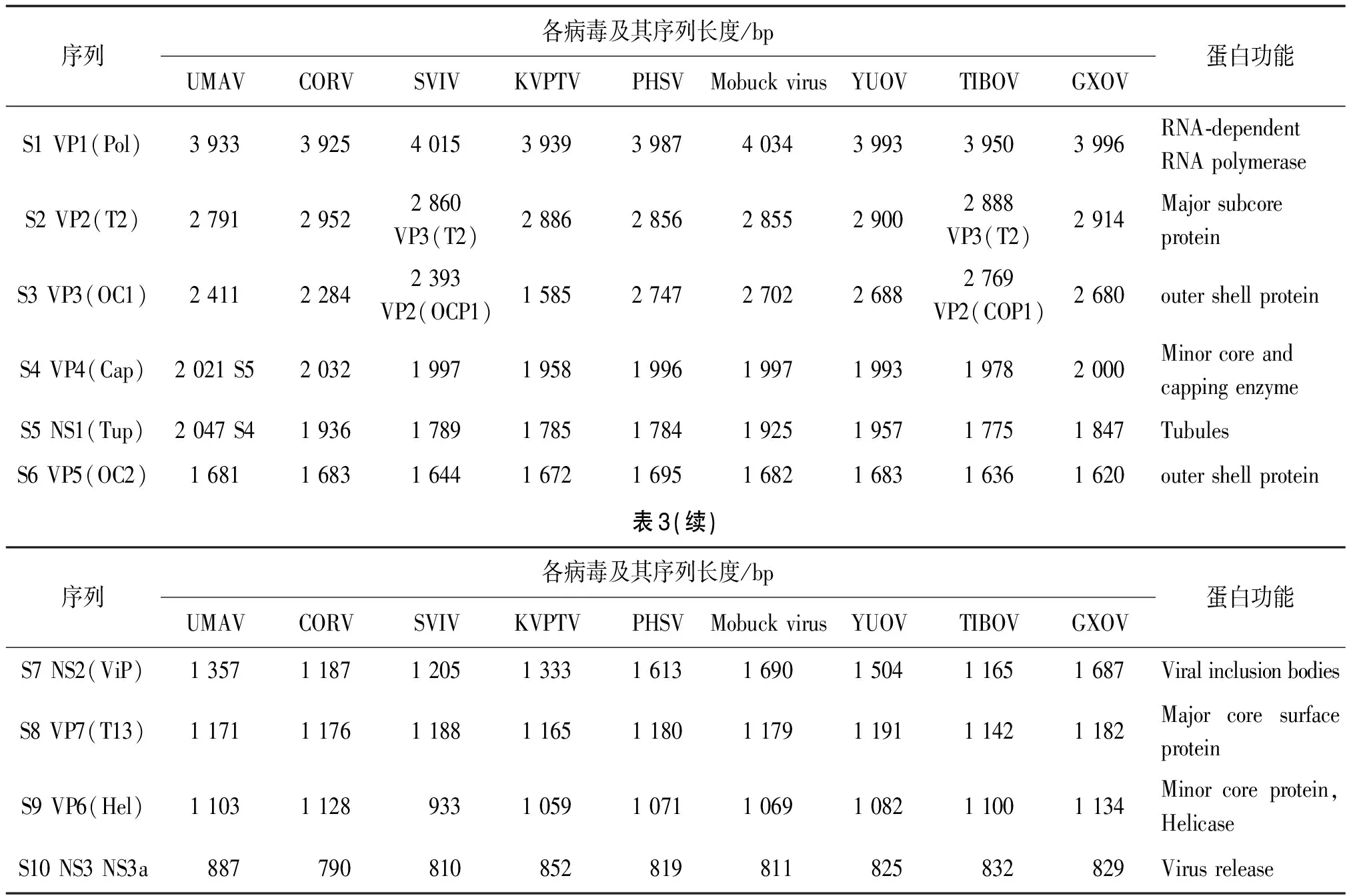
表3 环状病毒属各病毒分子结构及编码蛋白
环状病毒属的病毒主要流行于欧洲、亚洲和非洲[17]的30多个国家,以热带分布最广,亚热带高于温带。1988年徐普庭首次在云南省中华按蚊和棕头库蚊中分离到环状病毒[19],国内分布以云南、西藏、福建等地为主。1995年对西藏亚东和错那地区人群抗体调查发现,藏族人群感染率高于其他地区人群[12]。易感动物以 牛、羊和野生反刍动物为主,传播媒介可分为蚊、库蠓、蜱类[18,20],以库蠓传播为主,有记载秘鲁马病病毒可由蚊传播[21],Lebombo virus和Orungo virus、Changuinola virus最初从蚊中分离[22]。环状病毒以侵害神经系统为主,表现为3F(Fever、Face和Foot)症状:高热、面部充血、口鼻粘膜水肿、内脏充血、出血、跛行等。以弱毒疫苗和灭活疫苗预防为主,以支持疗法和对症疗法为辅,预后良好[11]。
1.1.1 尤马蒂拉病毒(Umatilla virus, UMAV) UMAV是1969年7月在美国俄勒冈州尤马蒂拉市的尖音库蚊(Culex pipiens)中分离到并第一个被完全测序的禽源环状病毒,基因组呈1-1-1-2-1-1-2-1的独特条带型,其节段4编码NS1(Tup),节段5编码VP4(Cap)。目前已发现4个亚种:Umatilla(UMAV),Minnal(MINV),Netivot(NETV)和Llano Seco virus(LLSV),MINV和NETV的基因组序列尚未报道[13]。2002年西澳大利亚金伯利地区环纹库蚊(Culex annulirostris)中分离到Stretch Lagoon orbivirus(SLOV),经VP2(T2)序列测定为UMAV新亚型,可感染马、驴和山羊,接种于BHK-21细胞后,48~72 h后出现细胞病变(cytopathic effect, CPE)[5]。2014年在日本东京端叉库蚊(Culex sasai)中分离到的Koyama Hill virus(KHV)病毒,是全球首次在东亚地区分离到的UMAV病毒,该病毒只能在C6/36细胞系增殖,48~72 h出现严重CPE,72~96 h可收集病毒[13]。
1.1.2 科里帕塔病毒(Corriparta virus, CORV) CORV是1960年从澳大利亚环纹库蚊中分离的一种环状病毒,电泳条带呈2-4-4(2-4-3-1)模式迁移与蠓传环状病毒3-3-4(3-3-3-1)条带型不同。目前已发现6种不同的CORV,其原型为CORV-MRM1,随后CS109、V654和V730病毒也在澳大利亚分离到,Acado virus和Jacareacanga virus分别于1963年埃塞俄比亚和1975年巴西的库蚊分离到[23]。该病毒主要流行于澳大利亚、非洲和南美洲,并在野生及家养禽类、牛、有袋动物、马和人体中发现中和抗体。1974年在加州环跗库蚊(Culex tarsalis)中分离到的California mosquito pool virus(CMPV),由于缺乏所有环状病毒的代表序列进行比对,当时认为是环状病毒的一个新种[23]。2013年Manjunatha.N等报道CORV-MRM1的全基因组测序后,经对比分析,认为CMPV属于CORV。CORV接种于BHK-21细胞后,48~72 h后出现严重CPE[23]。2016年Harrison等报道了澳大利亚北部环纹库蚊、Cx.Pullus、常型曼蚊(Mansonia uniformis)和诺曼伊蚊(Ae.Normanensis)中发现的Parry’s Lagoon virus(PLV)已在多种哺乳动物中发现中和抗体,但病毒分离只能在C6/36细胞中进行,5 d出现空泡、聚集等细胞病变[24]。
1.1.3 Sathuvachari virus(SVIV)/Kammavanpet virus(KVPTV) SVIV是1963年自印度南部黑冠椋鸟(Brahminy myna)中分离到一种新环状病毒,其片段2编码VP3(T2),片段3编码VP2(OCP1),但经VP1(Pol)、VP3(T2)、VP7(T13)序列分析鉴定为一种新型蚊传环状病毒的原型[25]。1980年自印度尼西亚杂鳞库蚊(Culex vishnui)中分离到Tagtag virus(TGV),经鉴定与SVIV同属一种病毒,2008年Yadav等对与SVIV同时分离的KVPTV的VP1、VP5(OC2)和VP7序列进行分析,确定与TGV归于一类,与UMAV的亲缘性最近[6,16]。近年来,1株自50年前越南蚊中分离的CGLT392被鉴定为SVIV,2005年在日本西南地区健康牛血清中分离到5株SVIV,研究表明SVIV目前已扩散至南亚热带以外的地区,对人和哺乳动物的致病性至今未知[26]。SVIV只能接种于C6/36[25];TGV接种于C6/36细胞后4 d出现CPE,接种于BHK和Vero细胞后5 d出现CPE[25];KVPTV不能在任何已知的细胞系中复制,可感染小鼠,对人和其他动物致病性未知[6, 16]。
1.1.4 秘鲁马病病毒(Peruvian horse sickness virus, PHSV) PHSV于1997年从死于神经系统综合症的秘鲁马血液、大脑和脾脏样本中分离出,经VP3(OC1)和VP7(T13)序列分析认为与YOUV最相近,是一种蚊传环状病毒。主要流行于南美洲热带地区,在马、牛、羊特别是家养马中传播,对马致病,表现为高烧(>39 ℃)、运动协调障碍、下颌下垂或口疮,通常8~11 d后死亡,最初主要在马等哺乳动物中发现,后来在圣马丁地区的蚊虫中也分离到该病毒。1997年圣马丁马匹病死率高达78%,至今未发现对其他动物或人类致病,1999年4—5月之间,在北澳大利亚出现神经系统疾病症的两匹马中,分离出了埃尔西病毒(Elsey virus, ELSV),经VP2(T2)和VP3(OC1)序列鉴定为与PHSV同一血清型,2002年又于健康马匹血液中分离出了4株PHSV。将PHSV接种于C6/36细胞后7 d可收集该病毒[27]。
1.1.5 Mobuck virus Mobuck virus是2012年8月在对美国密苏里州一只患肠出血热的雄性白尾鹿脾脏组织中进行流行性出血热病毒(epizootic haemorrhagic disease virus,EHDV)分离时发现的一种新环状病毒原型[28-29]。对Mobuck病毒VP2(T2)和VP7(T13)进行序列分析发现,与PHSV、YUOV和Middle Point orbivirus(MPOV)的亲缘性最近,属于蚊传环状病毒,该病毒接种于C6/36细胞中,通过剔除EHDV而得到[28],目前已报道全基因组序列,但对人兽致病性未知。
1.1.6 云南环状病毒(Yunan obivirus, YUOV) YUOV是1999年在中国云南省采集的三带喙库蚊(Cx.Tritaeniorhynchus)中首次分离的一种新型环状病毒[30-31]。随后在云南省中华按蚊(An.Sinensis)、棕头库蚊(Cx.Fuscocephalus)、菲律宾按蚊(An.Philippinensis)中也分离到YUOV。2005-2006年于西双版纳勐海县三带喙库蚊中分离到6株YUOV,并对中-老边境地区135例发热病人急性期血清标本进行YUOV抗体检测,发现1例IgM抗体阳性,8例IgG抗体阳性[8]。2007-2009年在西双版纳勐腊县三带喙库蚊中分离到5株YUOV[7]。YUOV基因组呈3-2-5条带型,建立于VP2(T2)和VP3序列分析,1997年分离自圣马丁PHSV感染马匹中的Rioja virus(RIOV)于2005年确定为YOUV血清1型,1998年分离自澳大利亚健康牛中的MPOV于2007年确认为YOUV血清2型,RIOV可引起羊和狗的脑炎症,MPOV不引起临床症状[27]。YOUV在C6/36细胞中CPE表现为细胞圆缩、脱落,也可在AP61和AA23细胞中复制[8,31],盲传三代可见CPE[7]。
1.1.7 西藏环状病毒(Tibet orbivirus, TIBOV) TIBOV是继YOUV后在我国首次发现的另一种新型环状病毒,主要由库蚊和蠓传播。2009年西藏林芝墨脱县圆斑按蚊(An.maculatus)中分离到的首株TIBOV(XZ0906),其基因组呈3-3-3-1条带型,VP3编码T2蛋白,与蠓传蓝舌病毒等更亲近,但其传播媒介以库蚊属居多[21, 32-33]。2008年在广东省的致倦库蚊(Cx.Fatigan)中发现的Fengkai virus也属于西藏环状病毒[17]。近年来,还在云南省采集的蠓虫和致倦库蚊标本中分离到TIBOV,2017年又确定了捕自2007年西双版纳的三带喙库蚊中的1株病毒为版纳环状病毒(Banna orbivirus, BAOV),经过系统发育分析表明BAOV为XZ0906和Fengkai的重组病毒,是中国首次报道的基因重组TIBOV[17]。2011年在湖北省首次从三带喙库蚊中分离到1株(HN11121),三带喙库蚊广泛分布于我国南部,中部及东部沿海地区[32],为TIBOV的传播提供了条件。2013年8月在云南省芒市库蠓中分离到DH13C120,经序列测定属于TIBOV,而与XZ0906代表不同的血清型,并首次证明了西藏环状病毒是家畜的潜在病原[21]。蚊中分离的TIBOV接种于BHK-21细胞48 h可见圆缩、少量破碎,72 h可见圆缩、脱落、破碎严重,不能引起C6/36的细胞病变;而库蠓中分离的DH13C120可引起Vero、C6/36细胞病变[21, 32-33]。
1.1.8 广西环状病毒(Guangxi Oribivirus, GXOV) 2015年在广西壮族自治区牛血样中分离到V172/GX/2015,基因组呈3-4-3条带型,经测序,其VP1(Pol)和VP7(T13)序列与YUOV、PHSV和Mobuck virus相似,VP2(T2)序列与蚊传环状病毒接近及其基因组核酸电泳条带与PHSV相似,都说明该病毒是一种新的蚊传环状病毒。目前只在牛身上分离出该病毒,但对其他哺乳动物及人类致病性未知。接种于C6/36细胞盲传4代,第5 d出现细胞皱缩、脱落、溶解与漂浮[18]。
1.2 东南亚十二节段dsRNA病毒属(Seadornavirus) Seadornavirus是呼肠孤病毒科的一个新属,由12节段双链RNA病毒构成,包含:BAV、KDV、LNV,主要在东南亚、印度尼西亚和中国流行。BAV是一种新型病原体,到目前为止,已从多种蚊子、蜱、脑炎患者、猪和牛中分离出,另外2种只从蚊子中分离出来[34]。这3种病毒都可从伊蚊、按蚊、库蚊中分离,仅BAV确认对人类致病[35]。ICTV第六次分类报告曾将KDV、BAV、LNY和科罗拉多蜱热病毒(Colorado tick fever virus, CTFV)归于Colti病毒属[36-38],随全基因组测序的完成,在2005年ICTV第8次分类报告中被确定为呼肠孤病毒科新属——Seadornavirus,该属病毒 VP1(RdRp)和VP3最保守,用于病毒种类的鉴定[9, 35, 38],各编码序列见表4。该病毒属最初在中国发现,并从我国云南省西双版纳地区发热病人的血液标本中分离到,随后在北京、甘肃、海南、新疆等地的人、家畜和蚊中分离到,我国目前已分离到170多株以BAV为主的东南亚十二节段dsRNA病毒[39]。近年来,又发现了两种新病毒:Balaton virus和芒市病毒,Balaton virus是从匈牙利淡水鲤鱼肠道中分离到的十二节段病毒,芒市病毒与Balaton virus和BAV最接近[35]。
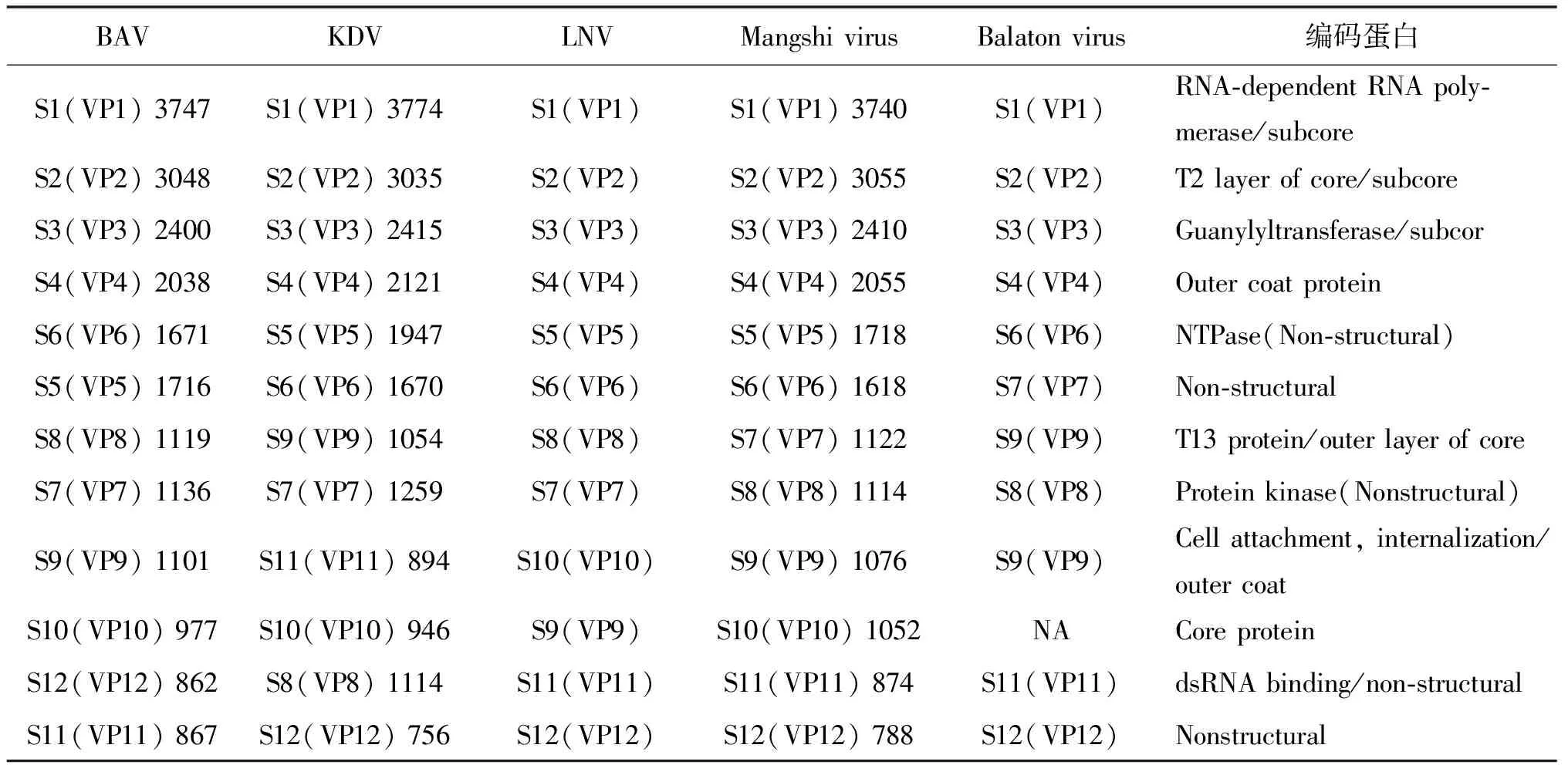
表4 东南亚十二节段病毒属各病毒分子结构、序列长度及编码蛋白
1.2.1 版纳病毒(Banna virus,BAV) BAV是引起脑炎和无名热的重要病原之一,最早于1987年由徐普庭等在云南省西双版纳的脑炎患者中分离得到,故命名为版纳病毒[9]。其基因组呈6-6条带型,VP12序列只有690 bp常用作基因分型鉴定,基因型分为A(中国、越南)和B(印度尼西亚)两型,A型又分为A1(北方)和A2(南方)两亚型,北方分离株多于南方[40]。分布于赤道至北纬42°之间的东南亚热带至中国北部的温带地区[9, 40]。媒介包括3属10种蚊虫。库蚊:三带喙库蚊、淡色库蚊(Cx.Pipienspallens)(淡色库蚊是在我国新发现的一种蚊虫媒介,并在中国境内广泛分布[41])、环带库蚊(Cx.Modestus)、伪杂鳞库蚊(Culex.Pseudovishnui)、凶小库蚊(Cx.Annulus);按蚊:迷走按蚊(An.Vagus)、中华按蚊;伊蚊:白纹伊蚊(Ae.Albopictus)、骚扰伊蚊(Ae.Vexans)和背点伊蚊(Ae.Dorsalis),近几年来发现BAV亦可由库蠓传播[9]。该病毒具有丰富的遗传多样性和巨大的分化潜能,且未出现物种屏障[42],不仅从患者体内分离,还从中国、印度尼西亚、南韩、匈牙利和越南的蚊子、猪、牛和蜱中分离103株[8],并在人群中发现BAV IgM/IgG,感染BAV者呈流感症,严重会出现昏迷、脑炎,猪、牛等则表现为消瘦甚至失明[35, 40-41]。BAV接种于C6/36细胞中48 h可见细胞聚集,72 h后聚集和脱落[9, 35]。
1.2.2 Kadipiro virus KDV于1981年在印度尼西亚爪哇岛褐头库蚊(Culexfuscocephalus)中分离出来,于1998年由法国学者Attoui H完成全基因组测序后归于Seadornavirus[39, 42]。该病毒基因组呈6-5-1条带型,是区别于BAV的重要特征。流行于印度尼西亚和中国,传播媒介为库蚊、按蚊、阿蚊和伊蚊,而在中国只从库蚊、按蚊和阿蚊中分离到该病毒[42],到目前为止,GenBank也只记录了两个完整的KDV基因组序列。2005-2006年首次在中国于云南省西北部收集到的三带喙库蚊、中华按蚊和骚扰阿蚊(Armigeressubalbatus)中分离到5株KDV[39]。2016年9月从山东省垦利市收集的中华按蚊中新分离出的SDKL1625,是北温带首例KDV,2018年我国首次报道该株病毒全基因组序列,目前KDV在我国南方和北方均有发现[42]。最近,在肯尼亚沿海地区进行的一项关于急性人类免疫缺陷病毒1型(HIV-1)感染的研究中,证实了KDV可能对人类致病[42],但目前尚无对人致病的报告[39]。KDV接种于C6/36细胞后4 d可见圆缩、细胞间隙加大,5~7 d逐渐脱落,而接种于BHK-21不出现CPE[39, 42]。
1.2.3 辽宁病毒(Liaoning virus,LNV) LNV是1997年在中国东北吉林白城采集的背点伊蚊中首次分离到的东南亚十二节段dsRNA病毒[43],电泳条带与KDV相同,其中 VP12最为保守,对VP10和VP12进行序列分析,可将LNV分为两个基因型,1997年辽宁株(NE91-31)为LNV-2。LNV严格分布于我国北纬36°-47°的西北-东北地区(新疆、山西、青海、甘肃及辽宁等地区[43]),表明辽宁病毒可能受气候限制[44],有学者认为LNV可能是近期引入北方的BAV新亚型[43, 45]。主要分布于新疆地区,2005年分离自新疆多地库蚊和背点伊蚊的27株均为LNV-1,此后新疆分离株均为LNV-1[43, 45]。其传播媒介主要是背点伊蚊,2005之后发现以凶小库蚊为主的库蚊携带该病毒,2012年8月在辽宁省从中华按蚊和背点伊蚊中分离到2株,这是第2次在东北地区分离到LNV-2,其余分离株均为LNV-1[43, 46]。该病毒能对小鼠致病,表现为病毒血症甚至死亡,目前尚无对人致病的报道,但可能对人和其他动物致病[44-45]。LNV接种于C6/36上表现为圆缩融合、脱落,于BHK-21上只表现为圆缩、脱落,接种后3 d出现CPE。LNJZ08024、LNJZ08045、LNJZ08033株只引起C6/36细胞病变,4 d可出现CPE[46]。
1.2.4 芒市病毒(Mangshi virus) 芒市病毒(DH13M041)是2013年在我国云南省德宏市的三带喙库蚊中首次发现的一种新东南亚十二节段dsRNA病毒,Balaton virus只在匈牙利鲤鱼中被鉴定但没有分离出来,而且只发现11个完整的基因序列(VP1-11),芒市病毒基因组呈6-4-2条带型,其VP8序列与BAV(VP7)和Balaton virus(VP8)编码的蛋白激酶序列最接近[35]。2005年在中国、老挝和缅甸库蚊中分离的MX6,通过基因组测序确定为我国西南边境首次报道的十二节段病毒,其VP9与芒市病毒的同源性仅81%,可能是同种病毒的不同基因型。芒市病毒接种于C6/36细胞中72 h后出现圆缩、脱落、融合,接种于BHK-21和Vero细胞后盲传三代仍未见CPE[35]。
2 刺突呼肠孤病毒亚科(Spinareovirinae)
根据ICTV分类,Spinareovirinae包含9个病毒属(见表1):对淡水鱼和甲壳动物类致病的水生呼肠孤病毒属(Aquareovirus,AQRV)[47],对人致病但经蜱传播的科罗拉多蜱传病毒属(Colitiviruses)[48],对昆虫致病的质型多角体病毒属(Cytoplasmicpolyhedrosisvirus,CPV)[49]和昆虫非包涵体呼肠孤病毒属(Idnoreovirus),对植物致病的斐济病毒属(Fijivirus)[50]和水稻病毒属(Oryzavirus)[51],对真菌致病的真菌病毒属(Mycoviruses)[52],对禽类致病的禽正呼肠孤病毒属(orthoreovirus)[53],仅一个从蚊中检测到的迪诺维纳病毒属(Dinovernavirus)[4, 54]。
Dinovernavirus只包含一种病毒——伪盾伊蚊呼肠孤病毒(Aedes pseudoscutellaris reovirus, APRV),是已知唯一具有9节段dsRNA基因组的病毒,其序列结构见图1,也是除了质型多角体病毒属(cypoviruses,CPVs)以外唯一的单衣壳呼肠孤病毒[4],1974年分离自斐济伪盾伊蚊的幼虫AP61细胞系。该病毒第7节段是唯一可以检测同源序列的片段,经Clamp序列分析表明,APRV代表了家族中一个独特的属,与质型多角体病毒属、斐济病毒属和禽正呼肠孤病毒属关系最密切,其主要传播媒介为白纹伊蚊、伪盾伊蚊、浆足蚊属(Eretmapodites dracaenae)[54]。2010年在喀麦隆的Fako地区进行的一项监测研究中第一次于自然界中发现迪诺维纳病毒,并初步确定为Fako virus(FAKV)[4]。相关研究证明,FAKV在白纹伊蚊中垂直传播和水平传播,水平传播以食源性传播为主,是否对哺乳动物和人致病还未可知。APRV不能在vero等哺乳动物细胞系中复制,接种于C6/36或C7/10细胞系后4 d便可见明显的细胞融合和聚集,而FAKV不引起任何细胞系病变,也不能在连续传代后产生RT-PCR产物[4]。
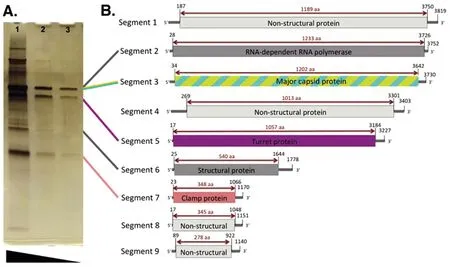
9个片段依次编码:非结构蛋白、RNA依赖RNA聚合酶、主要衣壳蛋白、非结构蛋白、炮塔蛋白、结构蛋白、夹道蛋白、非结构蛋白、非结构蛋白
3 结 语
目前全球已发现586种可传播虫媒病毒的媒介,其中蚊虫媒介有300多种[2],以伊蚊和库蚊为主要传播媒介[55],经蚊虫传播的呼肠孤病毒有3属13种,而在我国发现的全球首例就有2属6种(YUOV、TIBOV、GXOV、BAV、LNV、Mangshi病毒)。我国幅员辽阔,地理气候环境复杂,全球已证实的数千种蚊虫在我国就发现近500种,仅库蚊就达57种[56],主要分布于热带、亚热带地区,且同种蚊媒又能传播不同病毒,病毒输入后受气候等因素影响又发生变异,如新发现的BAOV是XZ0906和Fengkai virus的基因重组型,为我国首次报道基因重组型环状病毒。云南省作为我国西南边陲,地处热带及亚热带,国内多数虫媒病毒病首先在云南发现,据2012年查冰的调查报告显示,自1975年以来于云南省共采集到11属74种50余万只蚊虫标本,共分离361株病毒,其中321株经鉴定为5科5属11种病毒,呼肠孤病毒科包含YUOV、Colti病毒、KDV、BAV[57],还有近年来发现的Mangshi病毒等,这些病毒有的对动物致病,有的对人致病,有的在人群中发现抗体阳性,如YUOV和BAV自首次发现后,便很少再有发现及相关的研究报道。蚊传呼肠孤病毒的相关研究比较少,多数对人的致病情况还未知,而我国自然环境复杂,蚊虫种类多,分布广,有可能还存在其他蚊传病毒,有必要加强对其的病例监测和调查研究工作。因此全面掌握蚊传呼肠孤病毒的病原生物学特性、流行趋势,对其媒介蚊虫种类、分布及习性全面监控和研究,才能有效应对可能由该类病毒引发的疾病流行。
利益冲突:无
猜你喜欢
广西中医药大学学报(2021年3期)2021-12-01
大自然探索(2021年9期)2021-11-07
化工管理(2021年7期)2021-05-13
中国人兽共患病学报(2021年7期)2021-03-28
寄生虫与医学昆虫学报(2021年3期)2021-02-10
养殖与饲料(2019年11期)2019-02-26
中国畜禽种业(2018年9期)2018-01-20
环境昆虫学报(2016年4期)2016-08-23
广东海洋大学学报(2015年4期)2016-01-13
动物医学进展(2015年11期)2015-06-11
