云南省江城县杂鳞库蚊复合组蚊虫分子鉴定及COI基因序列分析
2015-06-11 02:21:40邓成杰王静林何于雯李华春
动物医学进展 2015年11期
邓成杰,王静林,周 洋,何于雯,胡 奇,李华春
(1.云南省畜牧兽医科学院,云南昆明650224;2.云南省镇雄县畜牧兽医局,云南镇雄657200)
杂鳞库蚊复合组包括三带喙库蚊(C.tritaeniorhynchus)、杂鳞库蚊(C.vishnui)、环带库蚊(C.annulus)、伪杂鳞库蚊(C.pseudovishnui)等数种亲缘关系较为密切的蚊种[1]。该复合组中三带喙库蚊、斑带库蚊等是乙型脑炎病毒等多种虫媒病毒的主要传播媒介,在我国分布较为广泛,也是我国南方地区的主要优势蚊种,主要吸血对象是人和家畜动物,对人类和家畜动物危害极大[2-3]。但是该复合组蚊虫形态学上非常相似,用传统形态分类的方法很难对这些蚊虫进行准确鉴定[4]。因此,本文对采集的杂鳞库蚊进行分子鉴定及COI基因序列分析,从分子水平了解三带喙库蚊等杂鳞库蚊遗传进化关系,为研究我国蚊虫分子分类提供资料,这对预防和控制以三带喙库蚊等杂鳞库蚊为传播媒介的疾病发生和传播具有重要意义。
1 材料与方法
1.1 材料
1.1.1 标本采集 2013年5月在云南省江城县农户牛圈采用诱蚊灯(功夫小帅,12V,300 mA,湖北武汉)采集标本,采集时间约为18∶00至次日早晨8:00 共14h。次日早晨收集标本,将标本置于-20℃冰箱中冷冻20min~30 min,待存活的蚊虫刚好冷冻死亡后,取出标本根据蚊虫形态学特征采用解剖镜进行蚊虫形态学鉴定,将形态学鉴定为杂鳞库蚊复合组蚊虫标本放入700 mL/L 乙醇中保存[5-6]。
1.2 方法
1.2.1 蚊虫DNA 提取及PCR 扩增 从乙醇中取出杂鳞库蚊复合组蚊虫标本,用DNA 提取试剂盒(天根公司)按说明书提取蚊虫DNA,操作步骤简要如下:每只杂鳞库蚊复合组蚊虫标本用缓冲液GA 200μL,充分研磨,加20μL ProteinaseK,56℃温浴过夜,加200μL缓冲液GB混匀,70℃放置10min。加200μL无水乙醇充分混匀15min,过柱,分别用500μL缓冲液GD 和600μL 漂洗液PW 洗涤,加60μL洗脱缓冲液TE 洗脱DNA。取2μL cDNA做反应模板,分别用COI基因特异引物[5]进行PCR扩 增,依 次 加 入10×buffer 5 μL,2.5 mmol/L dNTP 5μL,上、下游引物各1.25μL,r Taq酶0.75 μL,去离子31.75μL,充分混匀。PCR 程序为94℃5min;94℃30s,52℃30s,72℃60s,35 个循环;72℃延伸10min,10mL/L 琼脂糖电泳检查扩增条带,用TaKaRa DNA fragment Purification Kit纯化PCR 产物,PCR 产物通过华大基因公司进行测序[7-9]。
1.2.2 序列分析 使用Clustal X Version 2.1 软件包进行病毒基因核苷酸序列比对,DNA Star 软件中 Meg Align 进行核苷酸同源性分析。用MEGA 4.1 软件完成基于Neighbour-joining(NJ)方法的进化树绘制,Bootstrap值为1 000[10]。
2 结果
2.1 蚊虫采集
2013年5月在江城县共采集蚊虫标本148只,经形态学鉴定致倦库蚊112只(75.67%)、三带喙库蚊12只(8.11%)、骚扰阿蚊17(11.49%)、其他蚊虫7只(4.73%),表明了致倦库蚊是当地5 月的优势蚊虫种类。
2.2 核苷酸同源性分析
对云南省江城县采集到的12只形态学鉴定为三带喙库蚊标本(均为雌性,8只饱血,4只未吸血)进行DNA 提取和COI基因PCR 扩增,12只蚊虫标本扩增均为阳性,经序列测定获得580个核苷酸序列。12只库蚊COI基因核苷酸同源性在94.7%~100%,其中M2、M6、M7三只蚊虫核苷酸同源性在99.5%~99.8%,M1、M3、M4、M5、M8、M9、M10、M11、M12共9 只蚊虫核苷酸同源性在99.2%~99.8%,而两者之间核苷酸同源性在94.7%~95.7%。12只库蚊COI基因核苷酸与GenBank中杂鳞库蚊复合组的三带喙库蚊、杂鳞库蚊以及尖音库蚊复合组的淡色库蚊和致倦库蚊相应序列一起进行分析(表1),M1、M3、M4、M5、M8、M9、M10、M11、M12共9只蚊虫与三带喙库蚊核苷酸同源性最高为98%~100%,与杂鳞库蚊和伪杂鳞库蚊同源性分别在94.7%~95.5%和94.2%~94.5%,与尖音库蚊复合组的淡色库蚊和致倦库蚊同源性均低于93.2%;M2、M6、M7三只蚊虫与三带喙库蚊核苷酸同源性在94.8%~95.9%,与杂鳞库蚊和伪杂鳞库蚊同源性分别在94.4%~94.5%和93.9%~94.3%,与尖音库蚊复合组的淡色库蚊和致倦库蚊同源性均低于92.2%。

表1 云南省江城县杂鳞库蚊复合组COI基因核苷酸序列同源性分析Table 1 The homologous analysis of COI genes of Culex vishnui complex in Jiangcheng County,Yunnan Province
2.3 遗传进化分析
云南省江城县采集12只蚊虫与GenBank中12条库蚊COI基因序列一起构建系统进化树(图1)。结果显示所有24条库蚊COI基因序列形成2个较大进化分支,即杂鳞库蚊复合组库蚊进化分支和尖音库蚊复合组库蚊进化分支;云南省江城县采集的12只蚊虫序列位于杂鳞库蚊复合组库蚊进化分支内,提示云南省江城县采集的12只蚊虫标本均为杂鳞库蚊复合组库蚊。进一步对杂鳞库蚊复合组库蚊分析发现M1、M3、M4、M5、M8、M9、M10、M11、M12共9只蚊虫与三带喙库蚊位于同一进化分支内,提示这9只库蚊为三带喙库蚊;而M2、M6、M7蚊虫明显区别于杂鳞库蚊复合组库蚊的三带喙库蚊、杂鳞库蚊和伪杂鳞库蚊,单独形成一个进化簇,提示这3只蚊虫为杂鳞库蚊复合组的库蚊,分类地位的确定还有待于进一步的研究。
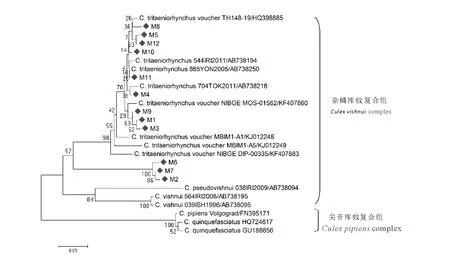
图1 云南省江城县采集杂鳞库蚊复合组蚊虫COI基因(600nt)遗传进化分析(◆代表本次采集蚊虫标本)Fig.1 Phylogenetic analysis of COI genes(600nt)of Culex vishnui complexs in Jiangcheng county,Yunnan province(Diamonds represent insects collected this time)
3 讨论
DNA 序列分析是鉴定生物体不同种、亚种和地理株,以及研究其系统进化关系的方法之一[11-12]。近年来,一些国内外学者采用线粒体基因COI、COII、COIII和ITS基因序列以及其他一些基因作为分子靶标进行蚊虫种类鉴定,其中线粒体基因COI进化速率适中,在能够保证足够变异的同时又很容易被通用引物扩增。目前,国内外众多学者采用COI基因作为分子靶标对等翅目、跳虫类和鞘翅目等昆虫进行深入研究,结果显示无论在物种鉴定还是系统发育方面COI基因都是不错的选择。因此,本次研究采用COI基因作为分子靶标对云南省江城县采集到的蚊虫标本进行分子鉴定,结果显示12只形态方法均鉴定为三带喙库蚊的标本,在系统进化树上形成2个进化簇,这2个进化簇的蚊虫标本之间的核苷酸同源性均低于95.7%,这个值明显低于蚊虫COI基因种内判定值在98%以上[13-14],表明这12只形态学鉴定为三带喙库蚊的标本分属于2种杂鳞库蚊复合组不同库蚊种。其中M1、M3、M4、M5、M8、M9、M10、M11、M12等9只蚊虫与三带喙库蚊形成通一进化簇,核苷酸同源性均在98%以上,表明这9只蚊虫标本为三带喙库蚊,而M2、M6、M7蚊虫在杂鳞库蚊复合组库蚊中单独形成一个进化簇,与三带喙库蚊和杂鳞库蚊核苷酸同源性均明显低于98%,提示这三只蚊虫并不是三带喙库蚊,而是杂鳞库蚊复合组的一种库蚊,分类地位的确定还有待于进一步的研究。
蚊虫是一类医学上非常重要的昆虫,其种类多,分布广。不同蚊虫种类的生活以及吸血等行为存在明显差异,这些差异直接关系到蚊传疾病的传播、流行及分布,因此必须对蚊科昆虫进行系统发育的研究,从蚊虫种群的演化历史和进化关系分析环境变迁因子对蚊类生物学特性的制约和影响,是解释现存蚊类的生态特性和传病关系的重要基础。本次研究开展形态学上相似的杂鳞库蚊复合组蚊虫种类的分子鉴定和遗传进化关系的研究,避免了形态学对相近蚊虫种类的错误分类,从分子水平较好明确这些蚊虫分类,为人和动物蚊传疾病的传播控制提供科学依据。
[1] 杨 明,陈汉彬.杂鳞库蚊复组28SrRNA 基因5′端序列研究[J].贵州科学,2003,21(2):185-187.
[2] 邓绪芳,史子学,邱亚峰,等.流行性乙型脑炎病原生态学的研究概况[J].动物医学进展,2011,32(2):111-114.
[3] Wang J,Zhang H,Sun X,et al.Distribution of mosquitoes and mosquito-borne arboviruses in Yunnan Province near the China-Myanmar-Laos border[J].Am J Trop Med Hyg,2011,84(5):738-746.
[4] 赵 民,陈创夫,盛今良,等.新疆地区蚊类携带西尼罗病毒情况调查研究[J].动物医学进展,2008,29(3):40-43.
[5] Flomer O,Black M.DNA primers for amplification of mitochondrial cytochrome C oxidase subunitⅠfrom diverse metazoan invertebrates[J].Mol Marine Biol Biotechnol,1994,3(5):294-299.
[6] 钟 敏,王 英,付蒙颖,等.中缅边境地区部分传疟按蚊蚊种的形态学及分子鉴定[J].第三军医大学学报,2014,36(24):2432-2436.
[7] 王 刚.基于DNA 条形码基础上的我国主要蚊虫分子分类系统的建立[D].北京:中国人民解放军军事医学科学院,2011.
[8] 郑学礼.我国蚊媒研究概况[J].中国病原生物学杂志,2014,9(2):183-187.
[9] 李国庆,陈 敏,刘本来,等.乌鲁木齐市尖音库蚊形态学及分子生物学鉴定分析[J].医学动物防制,2014,30(10):1066-1068.
[10] 刘春晓,顾大勇,赵纯中,等.成蚊形态学鉴定能力验证研究[J].中国国境卫生检疫杂志,2011,34(5):355-358.
[11] 杨飞龙,李旭东,闫振天,等.中华按蚊的分子鉴定标准[J].重庆师范大学学报:自然科学版,2014,31(4):40-45.
[12] 刘小波,次仁顿珠,郭玉红,等.拉萨市2009-2013年蚊虫种类构成及密度动态研究[J].中国媒介生物学及控制杂志,2014,25(3):200-204.
[13] 赵广宇,李 虎,杨海林,等.DNA 条形码技术在昆虫学中的应用[J].植物保护学报,2014,41(2):129-141.
[14] 张 莉,苏 生,吕召云,等.动物DNA 条形码鉴定技术及其在害虫种类鉴定中的应用[J].贵州农业科学,2014,42(4):152-156.
猜你喜欢
小学生学习指导(爆笑校园)(2022年9期)2022-09-30 06:49:16
保健医苑(2022年6期)2022-07-08 01:26:16
国画家(2022年3期)2022-06-16 05:30:18
今日农业(2021年12期)2021-11-28 15:49:26
大自然探索(2021年9期)2021-11-07 21:12:58
云南畜牧兽医(2021年1期)2021-02-24 04:37:28
当代音乐(2020年10期)2020-10-26 18:34:52
源流(2020年3期)2020-07-14 05:38:15
武汉广播影视(2017年3期)2017-06-01 09:21:42
环境昆虫学报(2016年4期)2016-08-23 03:13:46