
LncRNA H19对大鼠胰腺腺泡细胞AR42J细胞凋亡的影响
2021-08-31 07:38王小蝶余维丽王福贵
安徽医科大学学报 2021年8期
王小蝶,余维丽,王福贵,孙 昀
急性胰腺炎(acute pancreatitis, AP)是胰腺的炎症性疾病,严重时可引起全身炎症反应综合征,可导致单个或多个器官功能障碍,甚至死亡。AP的特征是蛋白酶异常激活、实质损伤、胰腺自身消化、胰腺腺泡细胞凋亡和坏死以及强烈的炎症反应。目前AP的治疗方法有液体复苏、营养支持等,尚无针对性药物治疗。而长链非编码 RNA( long non-coding RNA, LncRNA)是由200多个核苷酸组成,参与调节了许多细胞过程,如细胞凋亡、增殖、自噬等。许多研究已发现LncRNAs参与了多种生物学和病理过程。LncRNA H19是位于染色体11P15.5上的母系表达印记基因转录而来,并在某些疾病中发现了H19的上调,例如肝癌、食管癌、动脉粥样硬化、骨关节炎等。然而,LncRNA H19是否可以通过调节胰腺腺泡细胞凋亡从而参与AP的发生发展尚不清楚。该研究拟探讨LncRNA H19在大鼠胰腺腺泡细胞AR42J细胞凋亡中的作用机制,以期为AP的发病机制以及临床诊疗提供理论依据和思路。
1 材料与方法
1.1 主要材料
大鼠胰腺腺泡细胞AR42J购于上海富衡细胞库。LncRNA H19的siRNA(si-H19),siRNA对照(si-NC),空载质粒对照(pEX-3)以及过表达H19(pEX-3-H19)序列均购于上海吉玛制药技术有限公司。Lipofectamine 2000购于美国Life Technology公司;Opti-MEM(x1)培养基购于美国Gibco公司。雨蛙素粉末购于北京索莱宝科技有限公司;qRT-PCR试剂盒购于日本Takara公司。Caspase 3、Caspase 9以及β-Actin抗体均购于武汉三鹰生物技术有限公司。Annexin-FITC/PI细胞凋亡试剂盒购于上海七海复泰生物科技有限公司。1.2 细胞培养与转染
AR42J细胞使用含10%胎牛血清的高糖DMEM培养基,置于37℃、5%CO恒温培养箱中,每2~3 d传代1次。取对数生长期细胞接种至6孔板,待细胞长至60%~80%汇合度后,根据Lip2000说明书进行转染,转染成功后用雨蛙素(100 nmol/L)刺激24 h,处理好的细胞进行后续实验。实验分为6组记为:空白对照组(不做任何处理的AR42J细胞)、雨蛙素组、si-NC +雨蛙素组、si-H19+雨蛙素组、pEX-3+雨蛙素组、pEX-3-H19+雨蛙素组。1.3 qRT-PCR检测各组细胞中H19及凋亡相关基因Caspase 3及Caspase 9的蛋白的mRNA水平
取处理好的各组细胞,按照TRIzol试剂操作说明书提取细胞中总RNA。后逆转录成cDNA备用,流程参照逆转录试剂盒说明书。按照表1所示引物序列采用荧光定量PCR扩增仪按照以下条件进行荧光定量PCR反应,95℃、30s,95℃、5s,60℃、20s,共40个循环;95℃、1s,60℃、15s,95℃、5s,反应结束后以GAPDH基因作为内参照,计算H19、Caspase3及Caspase9的mRNA相对表达量。见表1。
表1 H19、Caspase3 、Caspase9以及GAPDH的qRT-PCR的引物序列
1.4 Western blot 法检测各组细胞中凋亡相关基因Caspase 3及Caspase 9的蛋白的表达水平
取处理好的各组细胞,加入RIPA裂解液,提取总蛋白,用BCA 试剂盒进行定量。然后进行SDS-PAGE凝胶电泳、转膜。用5%脱脂牛奶在摇床上室温封闭2 h后TBST洗3×10 min。用相应的一抗在4℃条件下孵育过夜。再用TBST洗3×10 min,用相对应二抗内室温摇床下孵育膜1 h,用TBST洗涤10 min×3次后,用ECL化学发光底物显影检测蛋白。最后用Image-J图像处理系统分析并以β-Actin作为内参照计算 Western blot灰度值。1.5 流式细胞仪检测各组细胞凋亡情况
取处理好的各组细胞,加入适量胰酶消化(不含EDTA),轻轻吹打细胞后500 r/min 离心5 min,弃上清液,用1 ml PBS重悬细胞,500 r/min离心5 min,收集细胞。加入400 μl Binding Buffer重悬细胞,再每组加入5 μl Annexin V-FITC,混匀后室温避光孵育15 min。最后加入10 μl PI染色液冰浴避光5 min。在30 min内进行CytoFLEX流式细胞仪检测,用CytExpert分析软件分析实验结果。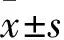
2 结果
2.1 沉默及过表达H19后各组中H19、Caspase 3及Caspase 9的mRNA表达水平
与空白对照组相比,雨蛙素组中的H19、Caspase 3及Caspase 9的mRNA表达水平增高(F
=9.097、25.58、58.10,P
均<0.05);雨蛙素组、si-NC+雨蛙素组、pEX-3+雨蛙素组中的H19、Caspase 3及Caspase 9的mRNA表达水平差异无统计学意义;而与si-NC+雨蛙素组相比,si-H19+雨蛙素组中的H19、Caspase 3及Caspase 9水平下降(F
=21.11、50.71、125.3,P
均<0.05);反之,与pEX-3+雨蛙素组相比,pEX-3-H19+雨蛙素组中的H19、Caspase 3及Caspase 9水平升高(F
=49.51、60.12、45.21,P
均<0.05),见图1。即说明H19沉默后,凋亡相关基因Caspase 3及Caspase 9的mRNA水平下降,而过表达H19后,凋亡相关基因Caspase 3及Caspase 9的mRNA水平上升。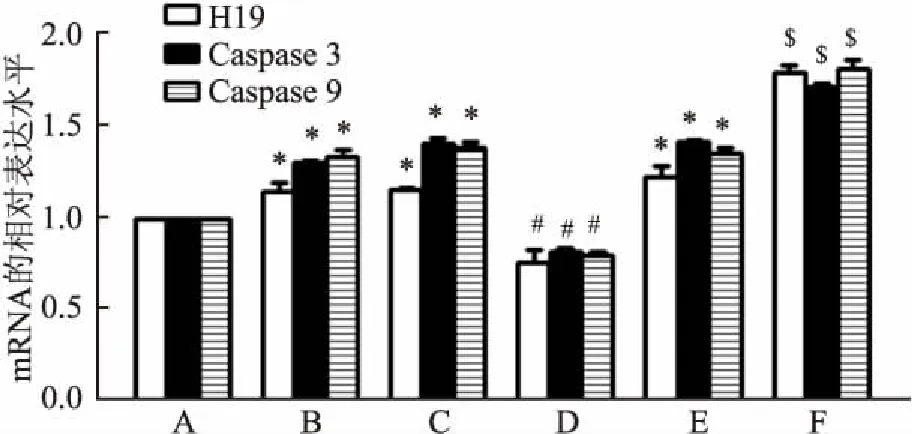
图1 qRT-PCR检测AR42J细胞中的H19、Caspase 3以及Caspase 9的mRNA表达
2.2 沉默及过表达H19后各组中Caspase 3及Caspase 9的蛋白表达水平
与空白对照组相比,雨蛙素组中的Caspase 3及Caspase 9蛋白水平增高(F
=5.182、37,P
均<0.05);雨蛙素组、si-NC组+雨蛙素组、pEX-3+雨蛙素组中的Caspase 3及Caspase 9蛋白水平差异无统计学意义;而与si-NC组+雨蛙素组相比,si-H19+雨蛙素组中的Caspase 3及Caspase 9蛋白水平下降(F
=11.17、34.13,P
均<0.05);反之,与pEX-3+雨蛙素组相比,pEX-3-H19+雨蛙素组中的Caspase 3及Caspase 9蛋白水平升高(F
=11.02、42.02,P
均<0.05),见图2。即说明H19沉默后,凋亡相关基因Caspase 3及Caspase 9的蛋白水平下降,而过表达H19后,凋亡相关基因Caspase 3及Caspase 9的蛋白水平上升。2.3 沉默及过表达H19后各组中细胞的凋亡率
与空白对照组相比,雨蛙素组的细胞凋亡率增高(F
=23.68,P
<0.05);雨蛙素组、si-NC组+雨蛙素组、pEX-3+雨蛙素组中的细胞凋亡率差异无统计学意义;而与si-NC组+雨蛙素组相比,si-H19+雨蛙素组的细胞凋亡率下降(F
=49.84,P
<0.05);反之,与pEX-3+雨蛙素组相比,pEX-3-H19+雨蛙素组的细胞凋亡率升高(F
=108,P
<0.05)。见图3。即说明H19沉默后,细胞凋亡率下降,而过表达H19后,细胞的凋亡率明显上升。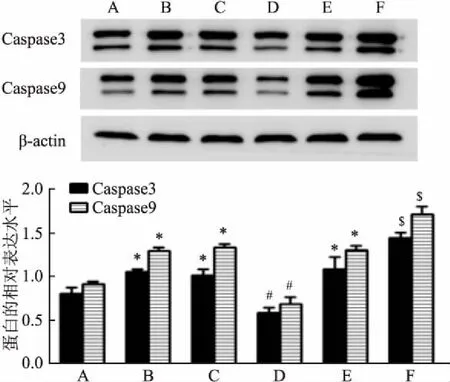
图2 Western blot 法检测各组中AR42J细胞中Caspase3、Caspase9的蛋白表达水平
3 讨论
有证据表明,LncRNAs参与了AP的发生和发展。然而,LncRNA H19在AP中的潜在机制尚不清楚。
LncRNAs已被证明是基因表达的主要调节因子,在包括癌症在内的各种生物功能和疾病过程中起着关键作用。LncRNA H19是一种印记的母系表达的转录物,是少数特征明确的LncRNA之一。H19的异常表达与多种疾病有关,且H19在炎症相关疾病中的表达水平通常较高。Wang et al实验发现,在类风湿性关节炎患者的滑膜组织中检测到H19的高表达,而磷脂酰肌醇 3 激酶(PI3K)可调控H19的水平抑制滑膜细胞的增殖及凋亡。而在乳腺癌中,H19的下调还可抑制参与细胞周期调节的p53的表达。在精原细胞的缺氧复氧模型中,H19在细胞复氧后显著增加,沉默了H19后,小鼠的精原细胞的凋亡水平明显增加。也有研究发现,H19在缺血再灌注的心肌细胞中上调,且敲除H19 可增加细胞活力,减少细胞凋亡,减少炎症细胞因子(白介素-1β、肿瘤坏死因子和白介素-6),抑制氧化应激。另一项实验表明,LncRNA H19作为miR-148b的上游,可调节WNT/β-连环蛋白信号通路,以促进氧化低密度脂蛋白刺激的血管平滑肌细胞的增殖并抑制其凋亡。半胱氨酸蛋白水解酶(Caspase)能够启动和维持细胞凋亡, 而其家族成员Caspase 3是最重要的凋亡蛋白酶之一, 通常位于哺乳动物细胞凋亡通路下游, 其表达量直接反映细胞凋亡程度。Caspase 3 可被Caspase 9直接激活,通过其自身介导的信号传导途径导致细胞发生凋亡。本研究用雨蛙素刺激在体外建立AP模型,与空白对照组相比,在雨蛙素刺激24 h后发现了H19的表达增高。本实验通过细胞转染成功获得H19沉默的AR42J细胞株,而沉默H19 后,Caspase 3及Caspase 9的表达水平下降,AR42J细胞凋亡水平下降,而过表达H19后,AR42J细胞凋亡水平上升。本研究说明H19可能参与并调控胰腺腺泡细胞的凋亡,这与上述文献一致。
图3 流式细胞术检测沉默及过表达H19后各组中细胞的凋亡率
综上所述, H19在AP的发展过程中高表达,沉默了H19后可下调Caspase 3及Caspase 9的表达,减少雨蛙素刺激引起的细胞凋亡,进而对大鼠胰腺腺泡细胞起到保护作用。本研究初步探讨了LncRNA H19在调节胰腺腺泡细胞凋亡的分子机制的认识,未来可深入研究H19的上游或下游靶基因,进一步探讨H19参与腺泡细胞凋亡的具体作用机制,以期为AP的临床诊疗提供一定的理论依据。
