
肌内脂肪与肉品质关系及其营养调控策略
2021-08-26 10:57:42吴婷,敖翔,何健
养猪 2021年4期
吴 婷,敖 翔,何 健
(1.西南科技大学生命科学与工程学院,四川 绵阳 621010;2.四川铁骑力士集团冯光德实验室,四川 绵阳 621006)
随着国民经济的快速发展,肉产品市场需求量的不断增加,促使了养殖业对猪生长速度、饲料转化率和瘦肉率的过度追求,导致猪肉品质变差,甚至丧失了原有的肉风味[1-2]。因此,解决猪肉品质、风味与养殖效率之间的矛盾,生产安全、高品质的畜产品,成为了当前养殖行业急需解决的问题。
为了有效改善肉产品质量,猪肉性状及其品质关系被广泛研究,结果发现,肌内脂肪含量及其脂肪酸组成不仅是影响猪肉感官品质、货架期、多汁性、嫩度、风味和加工价值的重要因素,还与肉的营养价值密切相关[3]。因此,了解肌内脂肪与肉品质特性的关系,是提高肌内脂肪含量和改善肉质的前提和基础,肌内脂肪的调控是提高猪肉品质的重要手段。以下将围绕肌内脂肪与肉质的关系、肌内脂肪形成的生物学基础,讨论猪肉品质研究现状和营养调控策略。
1 肌内脂肪形成的生物学基础
1.1 脂肪细胞的分化
肌内脂肪细胞的成熟晚于其他脂肪组织,它的形成开始于胚胎后期,是由小部分骨骼肌细胞分化而成,随着脂肪细胞的逐渐成熟,细胞质内出现脂肪滴,并扩大充满整个细胞,形成这些部位对于肌内脂肪的积累,从而在后期肌肉中产生大理石纹[4-5]。肌内脂肪的数量取决于胚胎时期和产后早期阶段肌内脂肪细胞的增殖;肌内脂肪组织的发育取决于肥育期脂肪细胞体积的增大和重量的增加[6]。
研究表明,脂肪细胞分化和形成的过程受一些转录因子的调控。过氧化物激活物受体γ(PPARγ)和ccAAT-增强子结合蛋白(C/EBP)是控制脂肪细胞形成的关键因子,两者的表达会引起多功能间充质干细胞向脂肪细胞的转化[7]。除了转录因子以外,一些激素因子也参与脂肪细胞的增殖和分化,例如糖皮质激素、胃动素等。糖皮质激素通过介导激活C/EBPδ转录因子的级联反应能有效诱导骨髓基质细胞分化为脂肪细胞[8]。胃动素能在增加PPAR、C/EBP、DGAT1和CD36表达的同时降低脂联素的分泌[9];并以浓度依赖方式刺激前脂肪细胞的增殖和线粒体活性,增加脂肪酸摄取、葡萄糖转运、3T3-L1细胞分化和脂肪组织中成熟脂肪细胞功能,从而实现对脂肪细胞分化和脂肪沉淀的积极作用[10]。
1.2 脂肪酸的形成
脂肪酸是脂肪组织的基础,其含量的变化受生物体对脂质消化吸收、合成和分解过程的影响(图1)。脂肪酸的从头合成是脂肪沉淀所需脂肪酸的大部分来源,脂肪酸合成酶(FAS)是该途径的关键作用酶[11],催化乙酰辅酶A和丙二酸单酰辅酶A合成甘油三酯,其表达水平的升高能显著增加脂肪在体内的沉淀[12-13];除此之外,还可以通过脂蛋白脂酶作用于低密度脂蛋白和乳糜微粒中的甘油三酯,水解产生相对分子质量较小的甘油和脂肪酸,再通过酯化合成甘油三酯[14],由此可见,提高脂蛋白脂酶表达水平能促进生物体对脂肪合成原料的消化吸收,也是增加脂肪在体内的沉淀的方式之一。脂肪的分解代谢过程与脂肪类型有关,例如磷脂在体内磷脂酶催化下水解产生不饱和脂肪酸,并以花生四烯酸为主[15-16];而甘油三酯的分解代谢主要是在脂肪细胞内激素敏感脂酶作用下,将脂肪分解为脂肪酸及甘油并释放入血供其他组织氧化[17-18](图1)。合成与分解使得脂肪的含量处于动态变化的状态,因此调节脂肪合成或脂肪分解相关酶的活性是决定生物体脂肪沉淀的关键,提供合成代谢过程中所需原料和能量是脂肪合成的支撑。
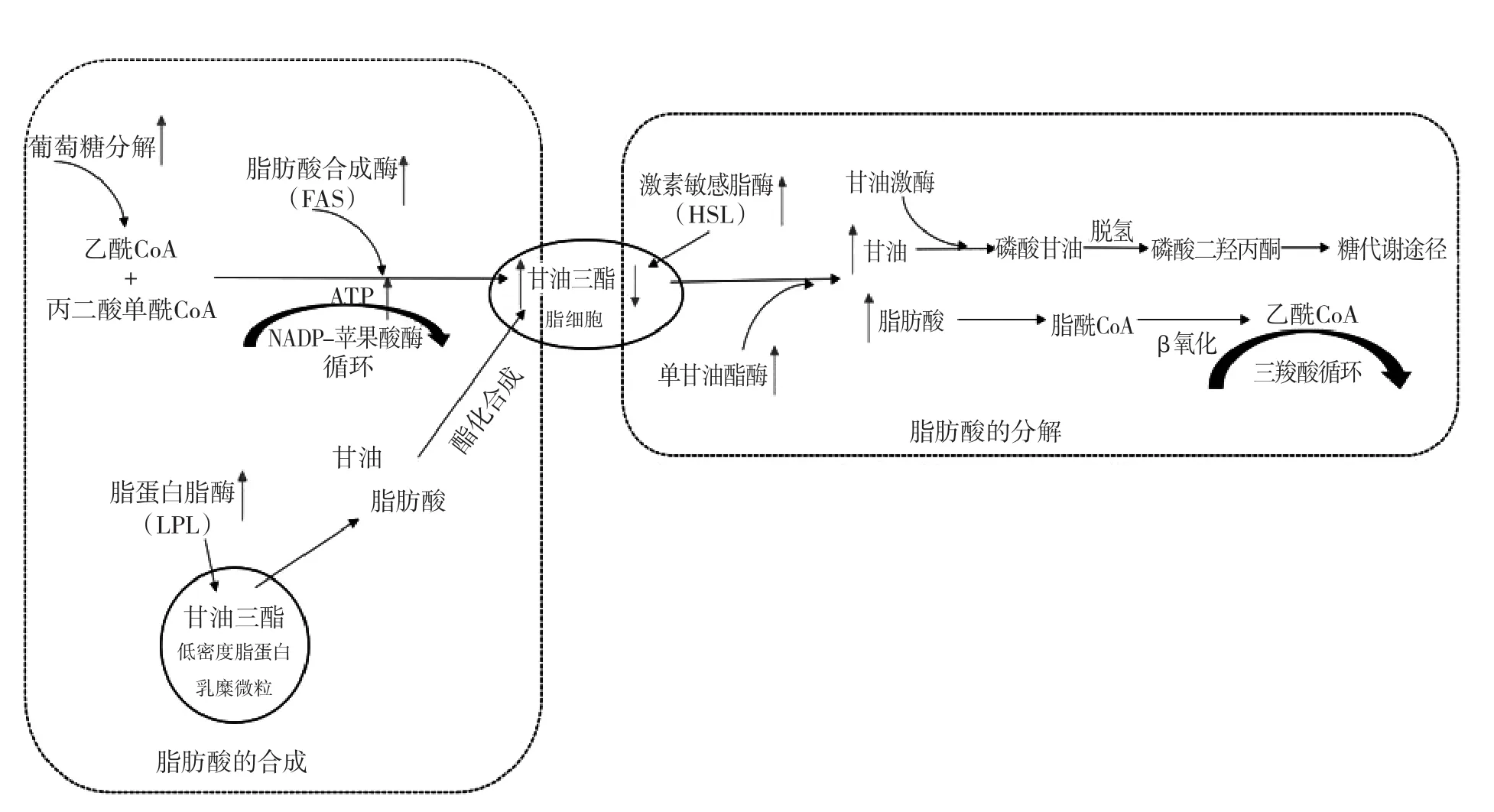
图1 甘油三酯代谢合成和分解代谢途径
2 肌内脂肪与肉品质
肉类消费过程中,消费者的感官指标包括颜色、大理石花纹、嫩度、风味等。大量研究表明,肌内脂肪含量对肉的感官品质有积极影响[3,19-22]。
2.1 肌内脂肪含量与不同品种猪肉品质
比较不同品种猪背最长肌平均肌内脂肪含量[23-39](图2),结果发现半野血藏猪、陆川猪、荷包猪、藏猪等未经选育或选育程度不高的地方猪种背最长肌中肌内脂肪含量高于杜长大三元、长大二元杂种猪等高度选育后的猪,浙江中白猪鲜肉中平均肌内脂肪含量更是低于2%的标准[40],这与消费者对目前市场上销售猪肉品质的评价基本吻合,说明肌内脂肪是影响猪肉品质的关键因素。不同品种之间肌内脂肪含量差异显著,可能与过去几十年里猪的遗传育种一直将提高胴体瘦肉率或降低背膘厚度作为主要目标,而肌内脂肪含量又与瘦肉率呈负相关有关[41]。
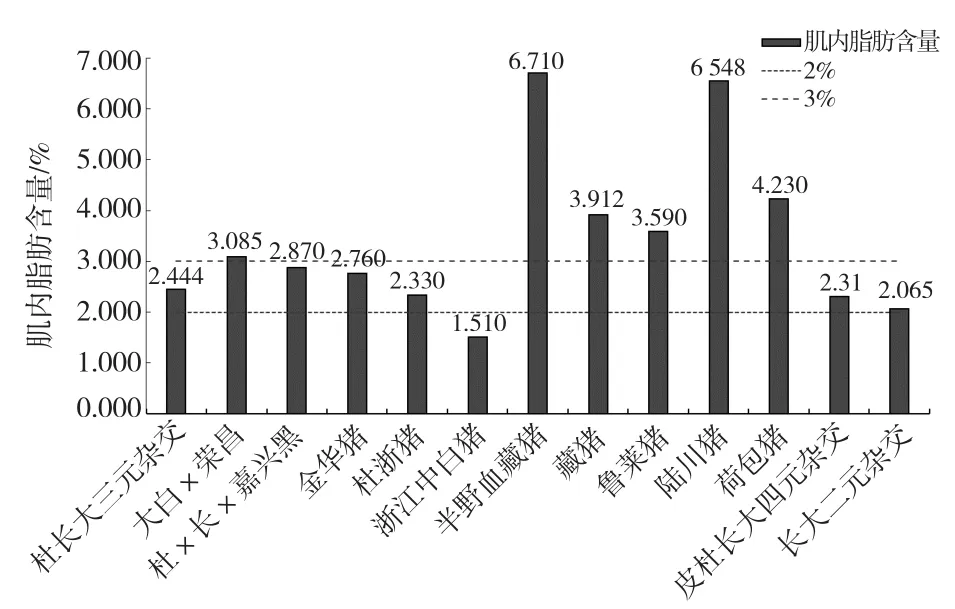
图2 肌内脂肪含量与猪品种关系
2.2 肌内脂肪与pH、肉色
统计分析肌内脂肪与pH、肉色和大理石纹之间的相关关系(表1)[23-39],结果发现,肌内脂肪含量与屠宰后45 min内新鲜猪肉的pH呈显著负相关(r=-0.359,P<0.05),与24 h的肌肉酸度呈正相关,虽然相关性不显著(r=0.352,P=0.52>0.05);屠宰后45 min内与24 h后肌肉pH之间呈显著正相关(r=0.413,P<0.05),说明肌内脂肪主要是对45 min内新鲜猪肉pH产生直接影响,对24 h后肌肉pH的直接影响不显著,后期pH主要受前期的影响,是热鲜肉在冷却排酸过程中产生糖酵解反应、大分子蛋白质分解和其他作用的结果[42]。肌内脂肪含量与肉色评分以及肌肉亮度(L*)、红度(a*)、黄度(b*)呈正相关关系,其中对亮度(L*)影响最大,主要是因为白色的脂肪细胞能提高肉的亮度,增加色差仪测定的L*值[43];另一方面,也与在pH下降过程中,肌蛋白变性,肌丝间静电斥力减小,并最终导致肌纤维收缩,肌肉系水力下降,肉表面对光的反射增加,产生浅色的外观[44]。肉的大理石纹即是肌肉纤维之间均匀分布的白色脂肪组织斑点或条纹。肌内脂肪含量与大理石纹评分有显著正相关关系(r=0.521,P<0.05),是影响大理石纹的直接因素[45]。研究表明,肌内脂肪增加46%,可使大理石纹斑点总数增加22%,平均大理石纹斑点尺寸增加14%[46]。Moon等[47]研究表明,大理石花纹与肉的品质等级、粗脂肪含量、华纳-布拉茨勒剪切力、蒸煮和滴水损失显著相关(P<0.01)。
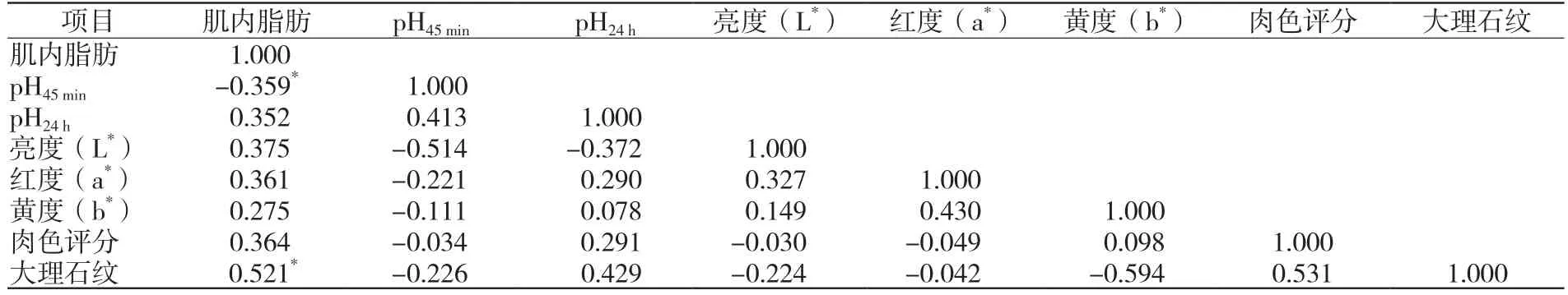
表1 肌内脂肪与肉色、pH的相关性
2.3 肌内脂肪与肌肉pH、水分的相关性
肌内脂肪能直接影响肌肉pH和水分,并通过影响肌肉pH间接影响滴水损失。统计分析肌内脂肪与肌肉pH、水分的相关性[23-39](表2),肌内脂肪含量与屠宰后45 min和24 h猪肉的pH分别呈显著负相关和正相关关系。pH45min与滴水损失呈正相关(r=0.132),初始pH越高,所造成的滴水损失越大[42]。pH24h与滴水损失之间呈显著正相关(r=0.601,P<0.05),是影响肌肉24 h滴水损失的重要因素。肌内脂肪含量与滴水损失和蒸煮损失之间相关系数不高,但两两之间有负相关关系(r=-0.194和r=-0.007),与Krzecio等[48]对猪肉加工价值的研究发现相符合。肌内脂肪含量与水分之间呈显著负相关关系(r=-0.451,P<0.05),这个结果与Watanabe等(2018)[49]的研究结果一致。
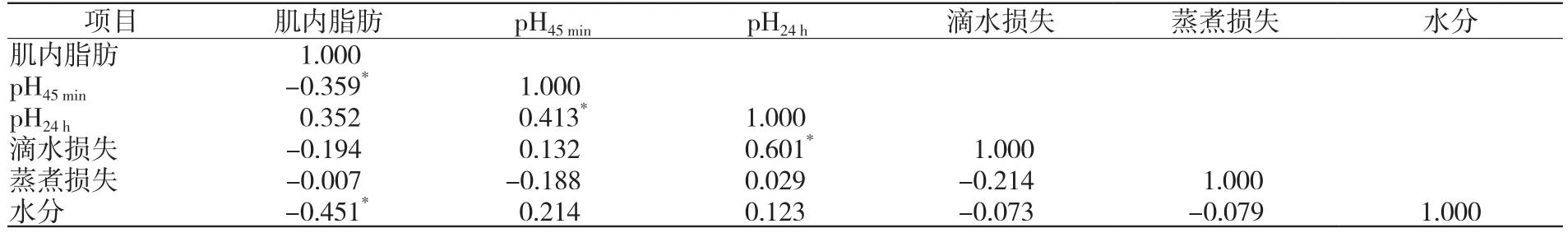
表2 肌内脂肪与肌肉pH、水分的相关性
3 肌内脂肪的营养调控
猪肌内脂肪细胞的形成和沉积所需要的营养物质都是从饮食中获得。肌内脂肪是脂肪组织中最后沉积的部分,相对于其他脂肪组织,它的生成和沉积受饲料营养水平的影响程度更大[50]。研究发现,通过调整肌内脂肪沉积过程中的营养水平可以调节组织代谢和基因表达,因此营养调控是改善肌肉组织肌内脂肪含量的有效途径。大量研究已证明,饲粮中能量/蛋白质水平、碳水化合物水平、氨基酸含量等对肌内脂肪的发生和沉淀具有重要影响作用[51-54]。
3.1 能量/蛋白水平
在饲粮对肌肉组织的调节中,蛋白质(氨基酸)与能量之比决定了猪机体蛋白质和脂肪沉积的比例。低蛋白质、低能量水平饲料可导致更高的胴体和肌内脂肪水平,对肉质产生正面影响。Gondret等[55]的研究中,低能低蛋白质饲粮(消化能12.96 MJ/kg,粗蛋白质13.1%)和标准饲粮相比较,增加了肌内脂肪含量而没有过多的胴体脂肪沉积。他们认为低能低蛋白质饲粮是增加猪肌内脂肪而不增加胴体脂肪的有效方式。Goerl等[56]研究饲粮中蛋白质水平对猪肉品质的影响时发现,随着饲粮中蛋白质水平增加,28~104 kg猪胴体背膘下降,瘦肉率增加,同时肌肉大理石纹减少,肉嫩度下降;而当饲粮中蛋白质不足时,能显著提高猪肉食用品质,可能是低蛋白质饲粮促使体内蛋白质周转加快和肌内脂肪含量升高所致。但是,未经赖氨酸含量调整的低蛋白质饲粮会降低商业杂交基因型猪的背最长肌面积[57]。Da等[58]给仔猪饲喂9~12周传统饲粮和试验饲粮(缺乏20%蛋白质和7%的能量)后发现,限饲使得红肌和白肌中肌内脂肪的含量显著积累。Leheska等[59]在类似试验中也发现,限饲和禁食可使猪有更长的背最长肌,更深的肌肉颜色和更高的大理石纹评分。Wood等[53]在考察品种、营养对猪生长速度和猪肉的脂肪程度、感官性状和脂肪酸组成影响时发现,饲料能量/蛋白质水平对不同品种猪的生长速度、胴体脂肪程度、肌肉大理石纹和脂肪酸组成的影响程度不同。因此饲粮能量/蛋白质水平对肌内脂肪的沉淀有积极影响并存在基因型的差异。
3.2 氨基酸
氨基酸不仅是合成蛋白质的底物,还能通过体内物质代谢转化为脂肪。据Hyun等[60-61]报道,添加亮氨酸和低水平赖氨酸能提高肌内脂肪含量。亮氨酸作为一种酮酸,在动物体内可以进一步氧化脱羧,生成酰基辅酶A衍生物并进入三羧酸循环,促进脂肪酸发合成,在猪饲粮中添加亮氨酸可以增加肌内脂肪;然而,整个文章中的数据是不一致的,他只是发现在低水平赖氨酸(0.5%)的饲粮中,肌内脂肪含量增加,但是当赖氨酸添加到0.7%时,没有任何影响。Madeir等[62]未发现在杜洛克皮特兰饲粮中添加亮氨酸还原蛋白对肌内脂肪有影响;Tous等[63]也没发现在低蛋白质水平或正常蛋白质水平饲粮中添加亮氨酸对肌内脂肪有影响。由于亮氨酸和其他支链脂肪酸之间存在拮抗作用,增加亮氨酸含量还可能损害其他氨基酸的性能[64]。关于低水平赖氨酸饲粮能提高肌内脂肪含量的结果得到了支撑。Pettigrew等[65]指出,当氨基酸摄入不足以使蛋白质积累率最大化,猪生长缓慢并产生较胖的屠体与更多的大理石花纹脂肪。Wang等[66]发现,随着赖氨酸水平从0.98%降至0.43%,肌内脂肪含量呈线性增加。但是也有不同的结果发现,Tous等[67]的研究结果显示,杜洛克长白猪的饲粮蛋白质水平降低、维持赖氨酸水平与对照饲粮相同或降低饲粮赖氨酸水平维持与对照饲粮相同的蛋白质水平,导致肌内脂肪增加;而当参数蛋白质和赖氨酸同时降低时对肌内脂肪含量不产生影响。进一步研究蛋白质与赖氨酸在肌内脂肪沉淀过程中的相互作用关系,发现膳食蛋白质限制引起的肌内脂肪增加是通过赖氨酸缺乏介导的[68-70]。与亮氨酸和赖氨酸一样,精氨酸在饲料中使用的研究结果也不尽相同。Guo等[71]发现,在饲粮中补充1.0%的L-精氨酸和谷氨酸能显著增加脂肪酸以及多种脂肪酸氧化衍生的挥发性化合物的形成的同时增加脂肪酸在肌肉中的积累和向肌内脂肪的转化,改善肉的嫩度、多汁性和整体食用品质。Go等[72]表示没有发现精氨酸对猪肌内脂肪有影响。有人还认为,添加精氨酸会减少肥育猪的脂肪沉淀,因为精氨酸在调节能量底物(脂肪酸、葡萄糖、氨基酸)代谢中的作用,部分通过产生一氧化氮来实现,而一氧化氮能增加脂肪酸氧化,减少三酰甘油酯的合成,增加基础脂肪的分解;Tous等[63]的研究结果显示,在正常蛋白质水平的饲粮中添加精氨酸可降低猪腰肉中大理石纹和肌内脂肪含量(P<0.05),也能降低腹部脂肪重量(P<0.01),增加瘦肉率(P<0.05)。但是在Tan等[73-74]的研究中有不同的发现,他们的结果显示,饲粮中补充L-精氨酸对猪脂肪组织和骨骼肌中脂质代谢基因的表达有不同的调节作用,有利于肌肉中脂肪的生成,同时有利于脂肪组织中的脂肪分解;在不增加或减少胴体脂肪的情况下增加肌内脂肪含量。由此可见,氨基酸能通过参与体内物质代谢或影响脂质代谢相关基因的表达影响肌内脂肪的沉淀,但其作用效果受饲粮蛋白质、赖氨酸水平和猪品种的影响。
3.3 碳水化合物
碳水化合物是动物饲粮中最为重要的三大营养物之一,不仅可以满足动物能量代谢的需要,还是动物合成一些大分子的前体物来源[75]。研究表明,肌内脂肪在合成过程中主要以葡萄糖或乳酸的碳原子作为脂肪合成的碳链来源[76];在肌内脂肪细胞表面发现较高葡萄糖转运担保-4(GLUT4)的表达,以及葡萄糖转化为长链脂肪酸所需要的磷酸果糖激酶和ATP-柠檬酸裂解酶类;此外,在肌内脂肪中葡萄糖合成脂肪酸的速率是乙酸合成肌内脂肪速率的2倍[77],这些研究结果说明,通过增加饲粮中葡萄糖或葡萄糖前体物质(如淀粉)的供应量可增加动物体内肌内脂肪的沉淀量。Kern等[78]在关于肉牛生长阶段对胴体组成影响以及与肌内脂肪沉淀相关基因的报道中提到,PPARγ mRNA和蛋白质的表达在摄食中期(111 d)和后期(188 d)高于早期(35 d)。考虑到PPARγ是脂肪生成的积极调节因子,与肌内脂肪沉淀相关的脂肪生成活动随着饲养时间的增加而增加[79],建议早期断奶和饲喂高淀粉饲粮,以改善肉牛的大理石纹。Moisá等[80]研究也发现,早期断奶-早期高淀粉喂养牛的脂肪细胞增生转录因子(PPARγ和C/EBPα)和成脂激活物(甲状腺激素反应、甾醇调节元件结合转录因子1和胰岛素诱导基因1)的mRNA水平以及几种产脂酶[FASN、硬脂酰辅酶A去饱和酶(SCD)、长链脂肪酸家族6的延伸酶和二酰甘油酰基转移酶2]在处理后96~167 d逐渐升高并达到峰值;正常断奶和饲喂方案下的肉牛在222 d时没有类似的表达水平,他们认为早期断奶-早期高淀粉喂养可促使PPARγ及其参与脂肪生成和脂肪生成的靶基因的早熟和持续激活,从而导致肌内脂肪(IMF)沉积增加,胴体等级提高。Guo等[81]研究也表明,低蛋白质/碳水化合物比例的饲粮(11.2%蛋白质和68.2%碳水化合物;10.1%蛋白质和69.3%碳水化合物)对PPARγ基因的表达量有积极影响。另有研究报道,PPARγ的表达能进一步促进C/EBPα的表达,为C/EBP和PPARγ协同作用于脂肪细胞的终末分化提供一个自我强化的调节环;C/EBPα又与过氧化物酶体增殖物激活受体γ共同刺激脂肪细胞分化[19]。脂肪细胞分化、增殖是影响肌内脂肪沉淀的关键因素。由此可见,生长发育早期高碳水化合物的饲料能促进脂肪的生成。
3.4 脂肪来源
饲料中脂肪会改变组织脂肪的含量与组成,特别是富含多不饱和脂肪酸的饲粮[82]。Bożena等[83]研究发现,猪饲料中脂肪来源对脂肪和水分含量有显著影响,亚麻籽油较菜籽油、猪油相比能显著提高n-3脂肪酸含量,减少n-6和n-9脂肪酸,提高以n-6/n-3比值表示的脂肪质量指标。Luo等[84]在研究亚麻籽对猪肌内脂肪含量及成脂相关基因影响是发现,随着含10%亚麻籽饲粮饲养时间的延长,肌内脂肪含量呈线性增加,同时,n-3多不饱和脂肪酸(PUFA)含量及过氧化物酶体增殖物激活受体d(PPARd)、PPARc、脂肪细胞脂肪酸结合的表达蛋白(aP2)和脂蛋白脂酶(LPL)也呈线性增加(P<0.01),而无翼相关MMTV整合位点10b(Wnt10b)的表达呈线性下降(P<0.01)。此外,n-3多不饱和脂肪酸的富集与这些基因的表达之间存在显著的(P<0.01)二次或线性关系,同时PPARc、aP2orWnt10b的表达与这些基因的表达和肌内脂肪含量之间存在显著的(P<0.01)二次或线性关系。这些数据显示,饲料中使用n-3脂肪酸含量高的脂肪能提高肌肉中n-3多不饱和脂肪酸的富集,进而增加肌内脂肪的沉积量,提高猪肉品质和营养价值。另外,饲粮中添加共轭亚油酸(CLA)能增加肉猪肌内脂肪细胞的大小,实现在减少猪背膘厚度的基础上增加肌肉的大理石纹,以提高猪肉的食用品质[85]。
3.5 其他添加剂
牛至精油是从植物牛至中提取的挥发性精油,具有抗氧化、改善肉质等功效。Cheng等[24]研究表明,从生长期到屠宰期间添加牛至精油能提高猪肉的肌内脂肪含量、嫩度、总体接受度;牛至精油的添加还能提高肌内脂肪中n-3多不饱和脂肪酸和单不饱和脂肪酸的百分比,提高腰背最长肌的氧化稳定性,减少腰背最长肌的滴水损失。
维生素A是基因表达的关键调节因子,已经证实维生素A通过调节参与脂肪生成和脂质代谢的几个基因的表达对脂肪组织产生影响;另外,维生素A对脂肪细胞的分化有负面影响,对处于增生过程中的组织细胞影响更大[86]。皮下脂肪细胞的分化早于肌内脂肪细胞,产后猪皮下脂肪组织的增加主要是肥大,增生是产后早期发生在肌内脂肪组织的主要事件[55],因此生长肥育期限饲维生素A对肌内脂肪含量没有影响。Kruk等[46]研究表明,限制维生素A,可在不明显影响皮下脂肪的同时使肌内脂肪显著增加。Ayuso等[86]研究发现,从仔猪断奶到出栏,饲喂去除维生素A的混合饲料可使伊比利亚猪的半膜肌肉中肌内脂肪含量增加47%(从4.60%增加到6.78%)。限制维生素A还具有重要的互补优势,维生素A和E都是亲油性维生素,它们在细胞和组织中的积累存在竞争,因此限制维生素A有利于维生素E的积累,这对肉的稳定性和货架期有重要的好处。并且维生素A和维生素E是猪饲料使用维生素中最昂贵的,减少预混合料中维生素A的含量,还能降低饲料成本[87]。
4 总结
肌内脂肪的功能体现是肉质,其含量直接影响猪肉酸度、肉色、大理石纹、滴水损失、水分等多个肉品质指标。胴体肌内脂肪含量可以通过控制生前猪不同阶段的饲料营养水平和添加特殊物质调节,但某些物质效果受猪品种和基础饲粮营养水平的影响,使用前还需作特有的验证。更多与肌内脂肪含量调节相关添加剂的开发应依托肌内脂肪沉积的生物学研究,生产安全、有效、无污染、无残留的调节剂,贴合绿色、健康的养殖理念。
(参考文献87篇略,有需要者请函索)
猜你喜欢
动物营养学报(2022年8期)2022-09-01 03:20:52
今日农业(2021年5期)2021-11-27 17:22:19
中国生殖健康(2020年5期)2021-01-18 03:00:06
今日农业(2020年17期)2020-12-15 12:34:28
中国化肥信息(2019年12期)2020-01-16 08:40:04
中国生殖健康(2018年5期)2018-11-06 07:15:56
动物营养学报(2017年8期)2017-08-16 09:40:00
数学小灵通·3-4年级(2017年5期)2017-06-05 09:12:37
动物营养学报(2015年9期)2016-01-07 11:29:44
中国洗涤用品工业(2015年8期)2015-02-28 19:02:49
