
鱼藤酮处理人神经母细胞瘤细胞的氧化水平研究
2021-08-25 02:04张萍邵靓
中国当代医药 2021年20期
张 萍 邵 靓
1.江西省人民医院神经内科,江西南昌 330000;2.江西省人民医院心血管内科,江西南昌 330000
多巴胺能神经元损伤和凋亡导致多巴胺分泌减少,是帕金森病的直接原因[1]。帕金森病多巴胺能神经元损伤及减少的原因和保护策略是目前帕金森病研究的重点及热点。多种因素均可引起多巴胺能神经元损伤,其中环境因素所导致的帕金森病日益增多,引起越来越多的重视[2]。如何减轻环境所致的多巴胺能神经元损伤,减少有机农药相关的帕金森病的发生具有现实意义。本研究从细胞氧化应激与帕金森病多巴胺能神经元减少的机制入手,从细胞氧化应激、细胞内抗氧化物及氧化物的水平干预等方面探讨鱼藤酮与多巴胺能神经元损伤的相关关系,可能为临床帕金森病的治疗提供切实有效的依据。
1 材料与方法
1.1 一般资料
1.1.1 实验细胞 人神经母细胞瘤细胞(human neuroblastoma cells,SH-SY5Y)细胞购于武汉大学中国典型培养物保藏中心。
1.1.2 仪器与试剂 本研究采用的主要仪器与试剂见表1。

表1 仪器与试剂
1.2 细胞培养
在37°C,5% CO2恒定条件下,用含1%抗生素(青霉素和链霉素)的10%胎牛血清(美国Gibco)培养SH-SY5Y 细胞2 d。
1.3 实验分组
根据细胞不同处理方式,将细胞分为正常组和鱼藤酮组:正常组中1 mL 培养基加入1 μL 二甲基亚砜(dimethyl sulfoxide,DMSO)溶液;鱼藤酮组中使用DMSO 稀释10 mmol/L 鱼藤酮溶液为不同浓度(25、50、100、200、500、1000、2000 nM)(nM:nmol/L)进行干预。
1.4 MTT 检测
3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT),能与活性较强的细胞线粒体中的琥珀酸脱氢酶反应,被还原为不溶于水的蓝紫色结晶并沉积在细胞中。使用酶标仪比色对比,在490 nm 波长处测定吸光值,可以间接反应存活细胞数量。
1.5 光镜观察细胞形态变化
活性较好的细胞呈椭圆形,突起较多;活性较差或死亡的细胞,突起消失变圆。
1.6 细胞内氧化应激水平检测
利用流式细胞术,在37°C 下保持20 min,使用2′-7′-二氯二氢荧光素二乙酸酯(DCFH-DA)染色正常组和鱼藤酮组细胞。其激发波长485 nm,发射波长530 nm。硫代巴比妥酸活性物质(thiobarbituric acid reactive substances,TBARS)和超氧化物根据实验试剂盒步骤评估超氧化物歧化酶(superoxide dismutase,SOD)水平。
1.7 统计学方法
采用SPSS 13.0 统计学软件进行数据分析,计量资料用均数±标准差(±s)表示,两组间比较采用t检验;计数资料用率表示,两组间比较采用χ2检验,以P<0.05 为差异有统计学意义。
2 结果
2.1 不同浓度鱼藤酮处理SH-SY5Y 细胞存活率的比较
分别使用25、50、100、200、500、1000、2000 nM 鱼藤酮干预SH-SY5Y 细胞后,以正常组细胞活性为100%,其他各组细胞与正常组比较,活性分别为(98.19±3.43)%、(95.79±3.95)%、(92.45±4.76)%、(86.63±3.89)%、(78.02±4.97)%、(62.50±5.21)%、(49.37±5.69)%。当鱼藤酮浓度达到200 nM 及以上时,细胞活性明显下降(P<0.05),与正常组比较,差异均有统计学意义(P<0.05)(图1)。
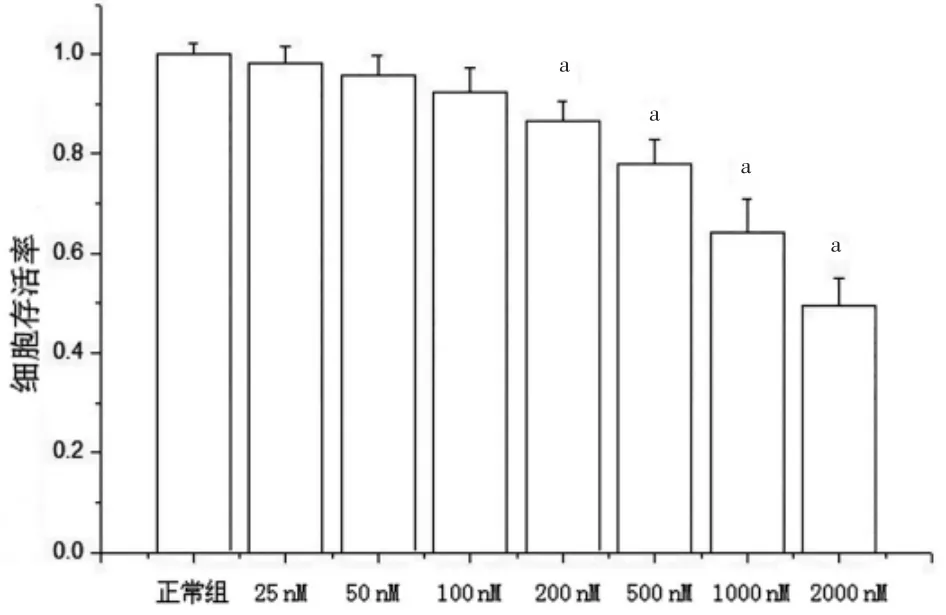
图1 不同浓度鱼藤酮处理SH-SY5Y 细胞存活率的比较
2.2 不同鱼藤酮处理的SH-SY5Y 细胞光镜结果
分别使用25、50、100、200、500、1000、2000 nM 鱼藤酮干预SH-SY5Y 细胞后,光镜下可见从200 nM开始细胞生长受限,随着浓度的改变,细胞形态学及细胞数量减少更为明显,2000 nM 的鱼藤酮加入培养基后细胞几乎停止生长,突起消失变圆,细胞生长与鱼藤酮浓度呈浓度依赖的负相关(图2)。
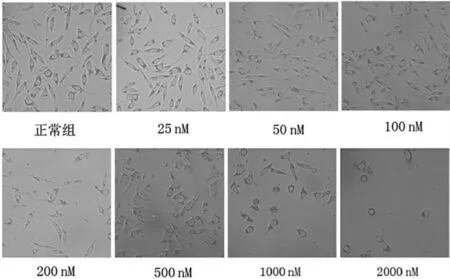
图2 不同鱼藤酮处理的SH-SY5Y 细胞光镜结果(×400 倍)
2.3 鱼藤酮干预后SH-SY5Y 细胞氧化应激水平的改变
选取500 nM 鱼藤酮处理的SH-SY5Y 细胞24 h后,利用流式细胞术测定细胞内氧化簇(reactive oxygen species,ROS)水平,结果显示,鱼藤酮组细胞内ROS 水平高于正常组,差异有统计学意义(P<0.05)(图3)。通过酶联反应比色检测发现,500 nM 鱼藤酮处理后细胞内SOD 表达水平低于正常组,差异有统计学意义(P<0.05);细胞内脂质过氧化表达水平高于正常组,差异有统计学意义(P<0.05)(图4)。
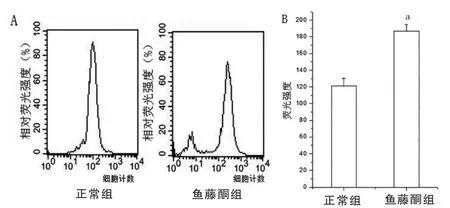
图3 500 nM 鱼藤酮处理细胞24 h 细胞ROS 水平
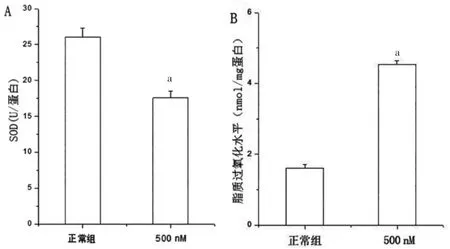
图4 500 nM 鱼藤酮处理的SH-SY5Y 细胞24 h 细胞SOD 水平及脂质过氧化水平
3 讨论
帕金森病又称震颤麻痹,是最常见的神经退行性疾病之一,主要病理表现为Lewy 体形成,中脑多巴胺能神经元变性死亡,纹状体多巴胺含量显著减少[3]。研究表明氧化应激是帕金森病发病机制中最常见的原因之一[4]。氧化应激通常指抗氧化物和氧化物间的失衡而导致的细胞损伤,包括抗氧化物如过氧化氢酶、超氧化物歧化酶、过氧化物还原酶家族、谷胱甘肽过氧化物酶等,以及氧化物如丙二醛、硫代巴比妥酸反应物质、丙烯醛等[5]。ROS 可以攻击核酸导致DNA 单链和双链断裂,DNA 蛋白交联、嘌呤和嘧啶碱基改变以及导致蛋白质编码RNAs 和非编码RNAs 的氧化修饰,可以攻击蛋白质的骨干和侧链,从而导致蛋白质的错误折叠和聚集[6]。同时,ROS 系统具有多种生物学作用,参与了激酶级联反应的活化,钙动员和信号传导、微调控氧化还原敏感性基因的表达、在神经干细胞的分化以及神经发生中起作用[7-13]。在帕金森病患者的血液中还原型谷胱甘肽显著高于正常组,而其它抗氧化因子浓度和过氧化氢酶的活性均有下降[14]。
长时间接触农药证实与神经系统变性疾病相关,这些农药包括百草枯、代森锰、艾氏剂、狄氏剂、异狄氏剂、甲氰菊脂以及有机磷等。这些农药均可诱导细胞氧化应激。已被确定与帕金森病发病相关的环境毒素1-甲基-4-苯基-1,2,3,6-四氢吡啶就可通过增加多巴胺能神经元内的氧化应激[15]。鱼藤酮是一种线粒体呼吸链复合体Ⅰ的抑制剂,能够通过抑制线粒体复合物Ⅰ,产生能量危机,而增加ROS 和OS 的产生,从而抑制线粒体呼吸链电子传递作用,造成跨膜电位差改变及线粒体膜通透性增强,导致细胞线粒体功能丧失,氧化应激激活,细胞凋亡坏死,最终造成在黑质致密部多巴胺能神经元的损失[16]。本课题组既往研究也显示触发氧化应激可以使暴露于氧化剂或淀粉样蛋白-B 的肽(抗体)诱导的半胱氨酸残基有利于分子间二硫键和随后形成细胞质聚集体的不可逆氧化,这种不溶性蛋白质可能最终促进细胞死亡[17]。本研究通过鱼藤酮处理多巴胺能神经母细胞系SH-SY5Y 细胞,证明其结果能造成氧化应激,这一定程度上可以模仿农药等毒物造成帕金森病的机制。
许多抗氧化剂的研究也证实对帕金森病有神经保护作用。如银杏叶提取物可对抗MPP+的神经毒性作用[18],另一项研究表明银杏叶提取物对MPTP 引起的氧化应激有保护作用[19]。促甲状腺激素释放激素因其抗氧化应激、抗谷氨酸盐的毒性作用、抗caspase诱导的细胞死亡、阻止DNA 片段化和抗炎作用而被研究于神经系统变性疾病如阿尔茨海默病与帕金森病[20]。既往本课题组研究也证实,通过敲低GAPDH 或者给予银杏叶提取物可以抑制氧化应激水平,从而改善鱼藤酮诱导的多巴胺能神经细胞死亡[21]。
综上所述,本研究通过鱼藤酮处理多巴胺能神经细胞,结果造成细胞内氧化应激,细胞内ROS 水平升高,脂质过氧化增强,SOD 水平下降,而抑制鱼藤酮通过氧化应激途径诱导的多巴胺能神经元损伤是治疗帕金森病的潜在策略。本研究仅限于体外细胞研究,有待进一步进行动物研究和人群研究证实降低氧化应激水平治疗帕金森病的有效性和安全性。
猜你喜欢
中老年保健(2022年5期)2022-08-24
中老年保健(2022年4期)2022-08-22
保健医苑(2022年6期)2022-07-08
昆明医科大学学报(2022年4期)2022-05-23
上海交通大学学报(医学版)(2022年3期)2022-05-05
读者(2022年9期)2022-04-22
老年医学研究(2021年5期)2022-01-19
心肺血管病杂志(2020年5期)2021-01-14
疯狂英语·新读写(2020年3期)2020-06-06
时代英语·高一(2019年5期)2019-09-03
