
大丽轮枝菌木糖苷酶基因的鉴定及基于HIGS技术的功能分析
2021-08-20 01:30张小雪孙天歌张迎春陈丽华张新宇李艳军孙杰
中国农业科学 2021年15期
张小雪,孙天歌,张迎春,陈丽华,张新宇,李艳军,孙杰
大丽轮枝菌木糖苷酶基因的鉴定及基于HIGS技术的功能分析
张小雪,孙天歌,张迎春,陈丽华,张新宇,李艳军,孙杰
石河子大学农学院,新疆石河子 832003
【】从大丽轮枝菌()中鉴定木糖苷酶基因,研究其与大丽轮枝菌致病力的关系,为解析大丽轮枝菌致病分子机制提供理论依据,同时为制定更好的棉花黄萎病防治策略提供科学依据。利用生物信息学方法从大丽轮枝菌基因组数据库中鉴定全部木糖苷酶基因,并对基因编码蛋白的结构域、基因的染色体定位及进化关系等进行分析。利用实时荧光定量PCR(qRT-PCR)技术检测木糖苷酶基因在不同抗/感棉花品种根系分泌物培养0、6、12、24和48 h大丽轮枝菌中的表达量。利用寄主诱导的基因沉默(host-induced gene silencing,HIGS)技术对木糖苷酶基因在大丽轮枝菌侵染过程中的功能进行初步分析。将的目标片段转化棉花,采用伤根法接种大丽轮枝菌Vd991,观察转化植株的表型,调查病情指数,同时利用qRT-PCR技术对植株中真菌生物量和的表达量进行检测。利用生物信息学方法从大丽轮枝菌中查找出13个木糖苷酶基因(—),其编码序列长度介于1 461—2 544 bp,蛋白质分子量介于38.78—90.97 kD,理论等电点介于4.67—5.89。结构域和进化树分析发现13个木糖苷酶基因中包括9个糖苷水解酶43家族成员、1个3家族成员和3个31家族成员。染色体定位分析发现13个基因分布在6条染色体上,未形成基因簇。qRT-PCR结果发现选取的6个基因均受到根系分泌物的诱导,在一种或多种根系分泌物中培养6 h或12 h后,表达量均明显升高,然后降低。其中受海岛棉根系分泌物诱导后表达量明显升高,表明该基因的表达明显受海岛棉根系分泌物的诱导。HIGS研究结果表明,接菌14 d和21 d后转化基因干扰片段的棉花发病明显较重,其病情指数(33.3和83.9)明显高于空载体对照(21.7和66.1)。qRT-PCR分析发现转化基因干扰片段的棉花植株茎中的表达量明显低于空载体对照,真菌生物量显著多于对照。利用HIGS技术将基因沉默后,棉株的抗病性明显降低,表明在大丽轮枝菌致病及宿主-病原体互作过程中可能发挥着重要的作用。
棉花;大丽轮枝菌;黄萎病;;木糖苷酶;寄主诱导的基因沉默
0 引言
【研究意义】我国是世界上主要的棉花生产和消费国,新疆是最大的植棉省区,已连续25年保持单产、总产和调出量全国第一。2019年新疆棉花播种面积254.05万公顷,皮棉产量500.2万吨,分别占全国的76.1%和84.9%,棉花产业已成为新疆的重要经济支柱和农民收入的主要来源。棉花黄萎病是一种土传性维管束真菌病害[1],严重影响棉花的产量和纤维品质[2]。我国棉区的黄萎病主要是由大丽轮枝菌()引起[3]。大丽轮枝菌入侵寄主后,其菌丝体大量生长阻碍植物木质部对营养物质的运输[4],同时产生水解酶和毒素等物质作用于寄主,最终导致植株死亡[1,5]。由于大丽轮枝菌具有稳定存在的微菌核结构以及多变的生理型,棉花黄萎病至今难以防治。挖掘大丽轮枝菌致病相关基因并进行功能验证,对揭示大丽轮枝菌的致病分子机制以及制定更好的棉花黄萎病防治策略具有重要意义。【前人研究进展】近年来,随着大丽轮枝菌全基因组测序的完成和生物信息学工具的发展,其致病相关基因的挖掘及功能研究已取得较大进展[6-10],但由于大丽轮枝菌致病机制非常复杂,其研究仍处于探索阶段。大丽轮枝菌入侵过程中会产生胞外分泌蛋白,其中含有大量的植物细胞壁降解酶,能够降解寄主植物的细胞壁多糖以达到入侵和定殖的目的[1,5,11]。植物细胞壁的横向结构主要由半纤维素和木质素结合而成。木聚糖是半纤维素的主要成分,最高可占细胞干重的35%,是植物中第二丰富的生物聚合物,仅次于纤维素。木聚糖的降解在大丽轮枝菌的致病过程中发挥重要作用[12]。木聚糖酶和木糖苷酶是降解木聚糖的关键酶,前者将木聚糖降解成小片段,后者则利用外切的性质将小片段水解释放木糖[13-14]。已有多种病原菌分泌的木聚糖酶被发现与其致病力密切相关[15-16],然而木糖苷酶与致病力的关系尚不清楚。依据木糖苷酶水解的糖苷键不同,可分为-木糖苷酶和-木糖苷酶两种类型。依据氨基酸序列相似性,在Carbohydrate Active Enzymes数据库(CAZy,http://www.cazy.org/)中-木糖苷酶被划分为GH31家族,-木糖苷酶被归类于糖苷水解酶GH3、GH30、GH39、GH43、GH52、GH54和GH120家族。-木糖苷酶的相关报道较多,目前狭义的木糖苷酶一般指的是-木糖苷酶。-木糖苷酶在自然界中广泛存在,在高等植物和微生物中均已得到分离,在能源工业、造纸工业和医药行业中已得到广泛应用[17-18]。【本研究切入点】真菌中已有多个木糖苷酶基因被克隆[19-20],大丽轮枝菌中木糖苷酶基因的鉴定及功能研究尚未见报道。实验室前期通过对不同抗性棉花品种的根系分泌物培养大丽轮枝菌的转录组分析,发现一个木糖苷酶基因(VDAG_01866)能够响应感病棉花品种的根系分泌物,暗示其在大丽轮枝菌早期寄主识别和侵染过程中发挥着重要作用[21]。【拟解决的关键问题】对大丽轮枝菌木糖苷酶基因进行全基因组鉴定和生物信息学分析,并利用qRT-PCR技术检测木糖苷酶基因在3种根系分泌物培养不同时间大丽轮枝菌中的表达量。利用寄主诱导的基因沉默(host-induced gene silencing,HIGS)技术对在棉花致病过程中的功能进行分析,探讨木糖苷酶基因与大丽轮枝菌致病力的关系。
1 材料与方法
试验于2019—2020年在石河子大学完成。
1.1 供试材料及处理
供试材料为陆地棉()新陆早8号(感病品种)、中植棉2号(耐病品种)和海岛棉()海7124(抗病品种),均由石河子大学棉花研究所提供。大丽轮枝菌强致病菌株Vd991由石河子大学绿洲生态农业重点实验室保存。
1.1.1 棉苗的种植 将棉种在温水中浸泡8—12 h后,在28℃恒温培养箱中催芽,挑选发芽整齐一致的种子均匀种植在霍格兰营养液中,每盆种植18棵棉苗,每3 d添加一次霍格兰营养液使总体积保持5 L,培养30 d后收集根系分泌物。先用滤纸对每盆中液体进行过滤,再用细菌过滤器(直径0.22 μm)进行过滤,然后在冷冻干燥机中将过滤后的溶液浓缩至1 L,放置在4℃冰箱备用。另外,挑选发芽整齐一致的种子均匀种在等体积混合的营养土与蛭石中,培养温度26℃,相对湿度80%,光周期16 h光照/8 h黑暗,待植株长出2片真叶后用于农杆菌的注射和大丽轮枝菌的胁迫处理。
1.1.2 大丽轮枝菌样本的制备 挑取PDA培养基上生长10 d左右的菌块接种于200 mL查氏液体培养基中,26℃、150 r/min振荡培养5—7 d,分装入50 mL离心管中,12 000 r/min收集Vd991菌体。每份称取0.5 g菌体,分别加入10 mL根系分泌物,培养0、6、12、24和48 h后收集菌体,设置3次生物学重复,共收集45份菌体。
1.2 方法
1.2.1木糖苷酶基因的鉴定及亚细胞定位预测 从大丽轮枝菌数据(https://fungi.ensembl.org/ Verticillium_ dahliae/Search/ Results?Species=Verticillium%20dahliae; idx=;q=arabinosidase;site=ensemblthis)网站下载Verticillium_dahliae. ASM15067v2. GFF3、CDS、genomic和protein序列文件,利用TBtools软件对GFF3文件中所有木糖苷酶基因进行搜索,初步获得大丽轮枝菌木糖苷酶基因,同时在CDS、蛋白质、全基因组序列文件中获取基因对应的cDNA序列、蛋白质序列以及基因组序列。利用SMART在线软件(http://smart.Emblh eidelberg.de/)进一步分析基因是否具有糖苷水解酶家族的主结构域,去除不完整的短片段和重复的序列,最终确定大丽轮枝菌中全部木糖苷酶基因。利用在线软件ExPASy-ProtParam tool(https://web.expasy.org/protparam/)分析氨基酸序列的蛋白质残基数(aa)、分子量(kD)和等电点(pI)。利用在线软件ProtComp9.0(http://www.softberry.com/ berry.phtml?topic=protcomppl&group=programs&subgroup =proloc)对蛋白质的亚细胞定位进行预测。
1.2.2 木糖苷酶基因的生物信息学分析 利用SMART在线软件预测木糖苷酶基因蛋白信号肽和结构域,利用DOG2.0软件绘制蛋白的结构域图。以13个木糖苷酶基因蛋白序列的主结构域为探针序列,利用NCBI Blast P(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行序列比对,从搜索结果中选择E值小、Score值大的来自其他物种的木糖苷酶基因序列。利用MEGA6.0提供的ClustalW程序对大丽轮枝菌及其他物种中的木糖苷酶基因蛋白序列进行多重序列比对,然后通过临位相连法构建系统进化树。从大丽轮枝菌数据库下载木糖苷酶基因的CDS、基因组序列,利用在线软件Gene Structure Display Server(GSDS)(http://gsds.cbi.pku.edu.cn)分析木糖苷酶基因外显子-内含子的数目及分布。从大丽轮枝菌全基因组数据库中获得木糖苷酶基因染色体分布以及长度信息,利用Map Inspect绘制木糖苷酶基因染色体定位图。
1.2.3 大丽轮枝菌总RNA的提取及cDNA第一链的合成 利用Fungal RNA Kit(OMEGA)提取大丽轮枝菌样本总RNA,用DNase Ⅰ消解DNA后,利用1.0%琼脂糖凝胶电泳检测RNA的完整性,利用NanoDrop 2000 DNA浓度测定仪测定RNA的浓度和OD260nm/ OD280 nm值。用MMLV反转录酶合成cDNA第一链,保存于-20℃冰箱备用。
1.2.4 木糖苷酶基因的表达分析 依据进化树分类结果及实验室前期转录组数据[21],从GH3、GH31和GH43 3个糖苷水解酶家族中挑选6个基因(、、、、和),其中来自GH3家族,和来自GH43家族,、和来自GH31家族。设计这6个木糖苷酶基因的特异性引物(表1),以培养0、6、12、24和48 h大丽轮枝菌的cDNA为模板,利用qRT-PCR对基因的表达模式进行分析,大丽轮枝菌微管蛋白基因作为内参基因。qRT-PCR反应在美国罗氏 LightCycler® 480系统上进行,反应参数:94℃ 1 min;94℃ 15 s,58℃ 20 s,72℃ 20 s;40次循环,所有反应设置3次重复,按照 2-ΔΔCT法计算基因的相对表达量。
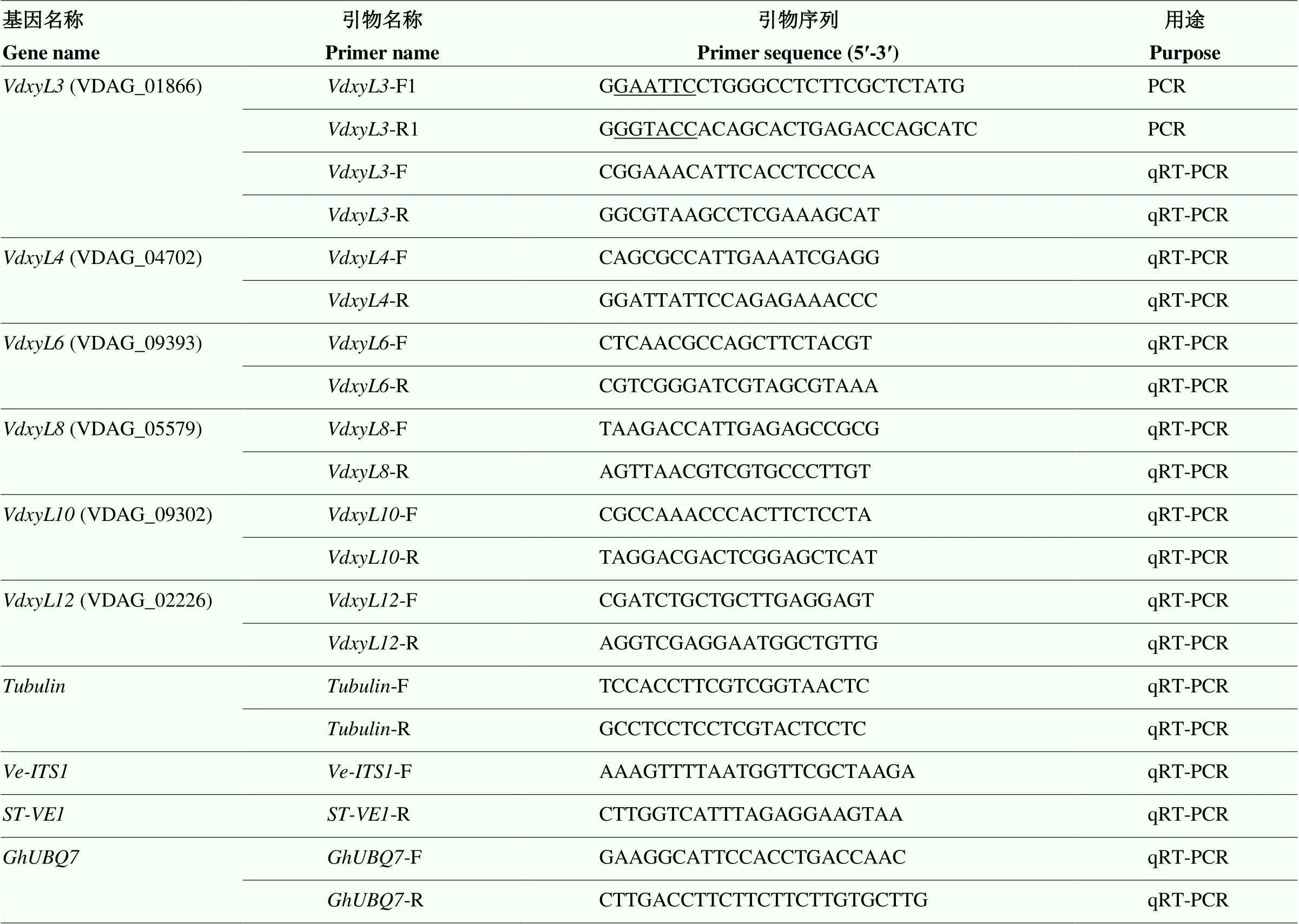
表1 引物序列
划线部分为酶切位点The underlined part is the restriction site.R I: GAATTC;I: GGTACC
1.2.5 基于HIGS技术的基因沉默 木糖苷酶基因干扰片段的克隆:根据NCBI网站Primer3-Blast在线设计的特异引物-F/R,在上下游引物中分别引入R I和I酶切位点(表1),以大丽轮枝菌cDNA为模板扩增的干扰片段。PCR反应体系(20 μL):ddH2O 13.2 μL,10×Ex Taq Buffer 2 μL,dNTP Mix(2.5 mmol·L-1)1.6 μL,cDNA模板1 μL;上游引物-F1(10 μmol·L-1)1 μL,下游引物-R1(10 μmol·L-1)1 μL,Ex Taq(5 U·μL-1)0.2 μL。扩增条件:94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 45 s,29次循环;72℃ 10 min。PCR产物经电泳检测回收后,连接至pMD19-T simple载体转化DH5感受态细胞,挑取阳性克隆送至深圳华大基因科技有限公司测序,获得T-重组质粒。
HIGS载体的构建:提取pTRV2空载体质粒和T-重组质粒,分别用R I和I进行双酶切,回收pTRV2载体线性化片段和干扰片段,采用T4DNA连接酶连接回收的两个片段,连接产物转化大肠杆菌感受态细胞,挑取单菌落摇培后提取质粒,利用PCR和双酶切对重组质粒pTRV2-进行鉴定。
棉花的注射及侵染:用电击转化法将pTRV1、pTRV2-、空载体pTRV2-和对照载体pTRV2-(该基因突变使叶片失绿)分别转化农杆菌GV3101。待棉苗两片子叶平展时,挑取含pTRV1、pTRV2-、pTRV2-和pTRV2-的农杆菌单菌落加入30 mL LB液体培养基(卡那霉素50 mg·L-1,利福平25 mg·L-1)中,28℃、200 r/min过夜培养。将摇培的菌液放入4℃离心机5 000 r/min离心10 min收集菌体,加入孢子悬浮液(10 mmol·L-1MgCl2、10 mmol·L-1MES(2-(N-吗啡啉)乙磺酸)和200 pmol·L-1AS(乙酰丁香酮),重悬后将菌液的浓度调至OD600=0.8。将准备好的含pTRV2-、pTRV2-和pTRV2-的重悬液分别与含pTRV1的重悬液按照1﹕1的比例混合,室温静置3 h后注射棉花。用1 mL无针头注射器采用压迫法注射生长10 d的棉苗,将菌液注入展平的子叶中,使菌液充满整个子叶。含pTRV2-和pTRV2-的重悬液分别注射80棵棉苗,将注射后的棉花放置于22—25℃条件下暗培养12 h,然后恢复至正常光照培养。
1.2.6 抗病性鉴定 大丽轮枝菌孢子悬浮液的制备:挑取PDA培养基上生长10 d左右的菌块接种于200 mL查氏液体培养基中,26℃、150 r/min振荡培养5—7 d,用灭菌的纱布过滤菌液后获得孢子悬浮液,在显微镜下用血球计数板计数,将孢子悬浮液调至1.0×107cfu/mL。
病情指数调查:注射农杆菌菌液后7—10 d,待pTRV2-处理的棉苗真叶完全呈现黄白色时,制备浓度为1×107cfu/mL的大丽轮枝菌孢子悬浮液,采用伤根法侵染棉花感病品种新陆早8号,每株接种20 mL孢子悬浮液。选取20株pTRV2-处理过的棉苗作为水处理对照(Mock)。侵染15—30 d后,对新陆早8号的感病情况进行统计[22]。
真菌生物量检测:接Vd991后14 d,分别采集pTRV2-和pTRV2-处理棉株的茎,提取总DNA(CTAB法),利用棉花内参基因(DQ116441.1)和黄萎病特异引物ITS1-F和ST-Ve1- R[23]进行qRT-PCR,测定其中大丽轮枝菌的生物量。
目标基因表达量检测:采用EASYspinPlus植物RNA提取试剂盒(Aidlab,北京,中国)提取接菌后14 d棉株茎的总RNA,反转录为cDNA,以为内参基因,-F和-R为引物,利用qRT-PCR对的表达量进行分析。
大丽轮枝菌恢复培养:接菌后14 d,随机选取pTRV2-和pTRV2处理棉苗各10株,用于大丽轮枝菌的恢复试验。从暴露于蛭石表面的茎基部切断植株,向上每隔2 cm取一个茎段,每个植株取3段。将茎段放入75%酒精浸泡30 s,然后在0.1%的升汞溶液中浸泡消毒5 min,无菌水冲洗3—5次,将处理的茎段均匀摆放在PDA平板上,25℃培养7—10 d。待大丽轮枝菌菌落长出后,观察菌落的生长情况。
2 结果
2.1 大丽轮枝菌木糖苷酶基因的鉴定
通过大丽轮枝菌基因组数据库的搜索结合在线工具SMART结构域分析,共鉴定出13个木糖苷酶基因,定名为—(表2)。这些木糖苷酶基因的编码序列(coding sequence,CDS)长度为1 461— 2 544 bp,编码的氨基酸为335—834个,蛋白质分子量介于38.78—90.97 kD,理论等电点介于4.67—5.89。亚细胞定位预测发现这些基因大多是细胞膜外的蛋白质,部分定位于细胞质、膜结合的溶酶体、细胞核或膜结合高尔基体上。13个基因中包含4个木糖苷酶/阿拉伯糖苷酶基因(),1个-葡糖苷酶/-木糖苷酶(),5个-木糖苷酶基因()和3个-木糖苷酶基因()。
2.2 大丽轮枝菌木糖苷酶基因编码蛋白的结构域分析
利用SMART在线工具(http://smart.embl-heidelberg. de/smart/batch.pl)对13个木糖苷酶基因编码蛋白进行了信号肽和结构域预测,分析发现VdxyL6含有糖苷水解酶Glyco_hydro_3(红色)和Glyco_hydro_3_C结构域(黄色);VdxyL4、VdxyL8和VdxyL12均含有糖苷水解酶Glyco_hydro_31结构域(绿色);其中VdxyL8还含有Gal_mutarotas_2结构域(橙色);VdxyL1、VdxyL2、VdxyL3、VdxyL5、VdxyL7、VdxyL9、VdxyL10、VdxyL11和VdxyL13均含有糖苷水解酶Glyco_hydro_43结构域(蓝色);其中VdxyL3含有两个Glyco_hydro_43结构域。13个基因编码的蛋白中VdxyL3、VdxyL6、VdxyL7、VdxyL9和VdxyL11含有不同长度的信号肽(黑色)(图1)。

表2 大丽轮枝菌中木糖苷酶基因的鉴定

黑色表示信号肽;蓝色表示Glyco_hydro 43结构域;绿色表示Glyco_hydro 31结构域;红色表示Glyco_hydro 3结构域;黄色表示Glyco_hydro 3_C结构域;橙色表示Gal_mutarotas_2结构域Black indicates the signal peptide; Blue indicates the Glyco_hydro 43 domain; Green indicates the Glyco_hydro 31 domain; Red indicates the Glyco_hydro 3 domain; Yellow indicates the Glyco_hydro 3_C domain; Orange indicates the Gal_mutarotas_2 domain
2.3 木糖苷酶基因多重序列比对及进化树的构建
通过在线软件NCBI中多序列比对查找并选取其他物种中12个木糖苷酶基因编码的蛋白序列,其中XP 018236340.1(尖镰孢)、RMZ44074.1(黄曲霉)、TFL04453.1(细翼蕨)、TDZ59664.1(三叶炭疽菌)、TEA21443.1(紫苏炭疽菌)和THX71209.1(出芽短梗霉菌)属于糖苷水解酶43家族;CVK94262.1(芒果镰孢)和CVL11996.1(层出镰孢)属于糖苷水解酶3家族;RKK75644.1(尖镰孢)、KPM38392.1(新蜜环菌)、XP 003721488.1(稻瘟病菌)和XP 028469785.1(碱性钠酵母菌)属于糖苷水解酶31家族;将12个木糖苷酶基因与大丽轮枝菌13个木糖苷酶基因编码的蛋白进行系统进化树分析(图2),25个蛋白被清晰地分成3组。VdxyL1、VdxyL2、VdxyL3、VdxyL5、VdxyL7、VdxyL9、VdxyL10、VdxyL11和VdxyL13与其他物种中43家族成员聚为一类;VdxyL6与其他物种中3家族成员聚为一类;VdxyL4、VdxyL12、VdxyL8与其他物种中31家族成员聚为一类;该结果与结构域预测结果一致,表明大丽轮枝菌13个木糖苷酶基因中,包括9个糖苷水解酶43家族成员,1个糖苷水解酶3家族成员和3个31家族成员。

红色表示Glyco_hydro 3家族;紫色表示Glyco_hydro 31家族;蓝色表示Glyco_hydro 43家族
2.4 木糖苷酶基因结构分析
利用在线软件GSDS对13个木糖苷酶基因结构进行分析,结果如图3所示和含有5个外显子和4个内含子;含有4个外显子和3个内含子;、和含有3个外显子和2个内含子;、、、和含有2个外显子和1个内含子;和含有1个外显子,无内含子。
2.5 大丽轮枝菌木糖苷酶基因染色体分布
大丽轮枝菌共有8条染色体,13个木糖苷酶基因分布在6条染色体上。Chr02、Chr03和Chr07上分别含有3个木糖苷酶基因,Chr02上分布的基因为、和,Chr03上分布的基因为、和,Chr07上分布的基因为、和。Chr04上分布2个基因和。Chr01和Chr08上各含有一个木糖苷酶基因,分别为和(图4)。根据200 kb核苷酸中含3个以上基因为1个基因簇的定义,木糖苷酶基因没有形成基因簇。

图3 大丽轮枝菌木糖苷酶基因结构分析

图4 大丽轮枝菌木糖苷酶基因在染色体上的分布
2.6 木糖苷酶基因在根系分泌物诱导下的表达分析
qRT-PCR结果发现6个基因均受到根系分泌物的诱导,在一种或多种根系分泌物中培养6 h或12 h后,表达量均明显升高,然后降低。在海7124根系分泌物诱导6 h的表达量明显高于其他样本,表明该基因的表达明显受海岛棉根系分泌物的诱导。、、、和在耐病品种(中植棉2号)根系分泌物诱导6 h样本中的表达量明显高于其他样本,其中的表达仅受到耐病品种根系分泌物的诱导,的表达也受感病陆地棉品种(新陆早8号)根系分泌物的诱导,、和受3种不同抗/感品种根系分泌物的诱导(图5)。该研究结果表明大丽轮枝菌木糖苷酶基因在响应棉花根系分泌物的过程中可能发挥着重要作用。根据qRT-PCR结果结合实验室前期转录组数据,选取用于进一步的功能研究。
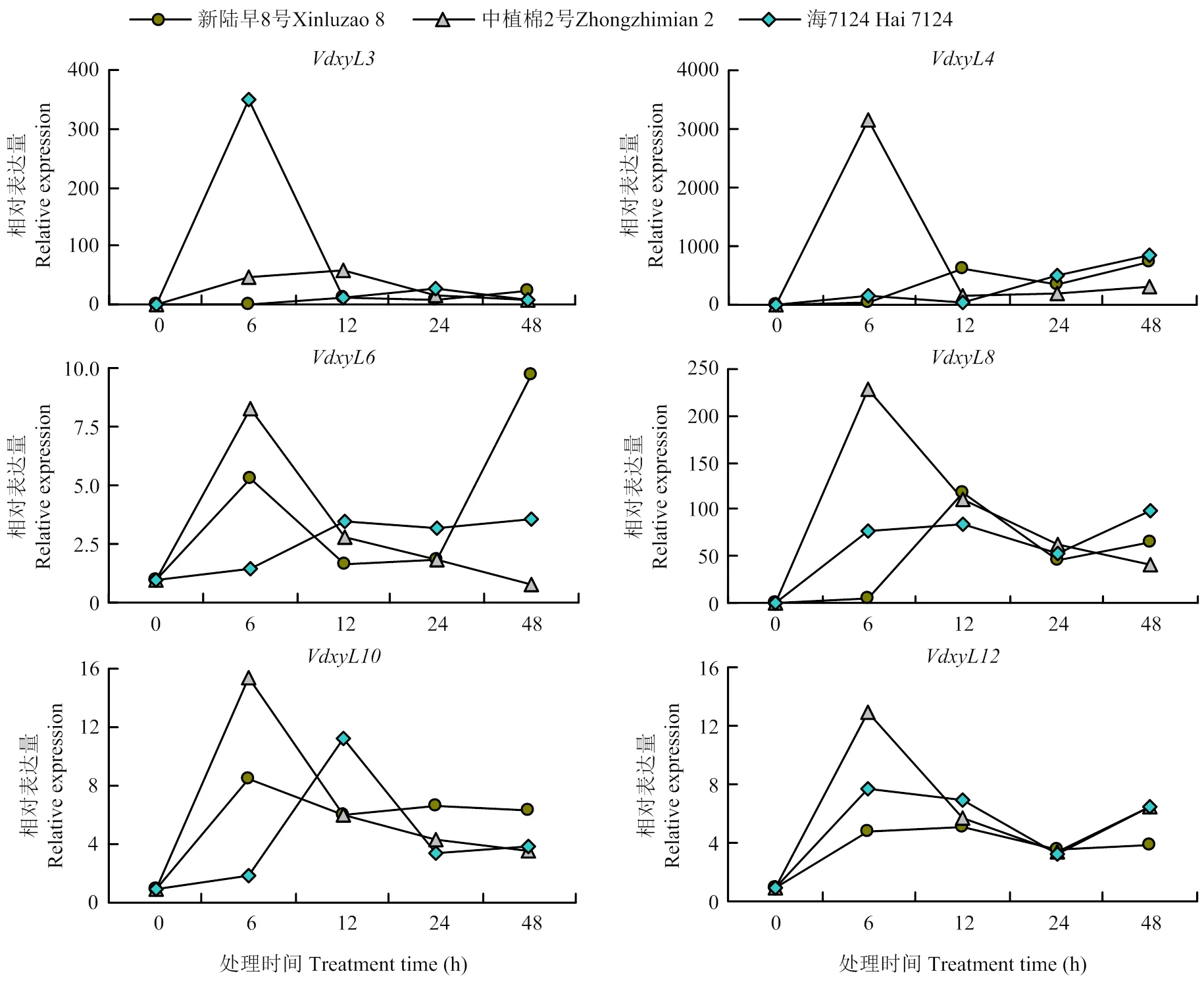
图5 木糖苷酶基因在不同棉花品种根系分泌物培养的大丽轮枝菌中表达模式
2.7 HIGS沉默效果检测
选取进行HIGS沉默研究。将注射pTRV2-的棉花作为阴性对照,注射pTRV2-的棉花为阳性对照。注射10 d后,pTRV2-处理植株真叶出现黄化现象(图6-A),表明HIGS体系可以成功抑制目标基因表达。制备浓度为1.0× 107cfu/mL大丽轮枝菌孢子悬浮液,采用伤根法侵染棉花。接菌14 d后,采集pTRV2-和pTRV2-处理的棉花茎,提取RNA并反转录为cDNA,以-F/R为引物,利用qRT-PCR分析的表达水平,结果发现pTRV2-处理的棉花茎中的表达量明显低于pTRV2-(图6-B),表明pTRV2-处理棉株中大丽轮枝菌的表达成功受到抑制。
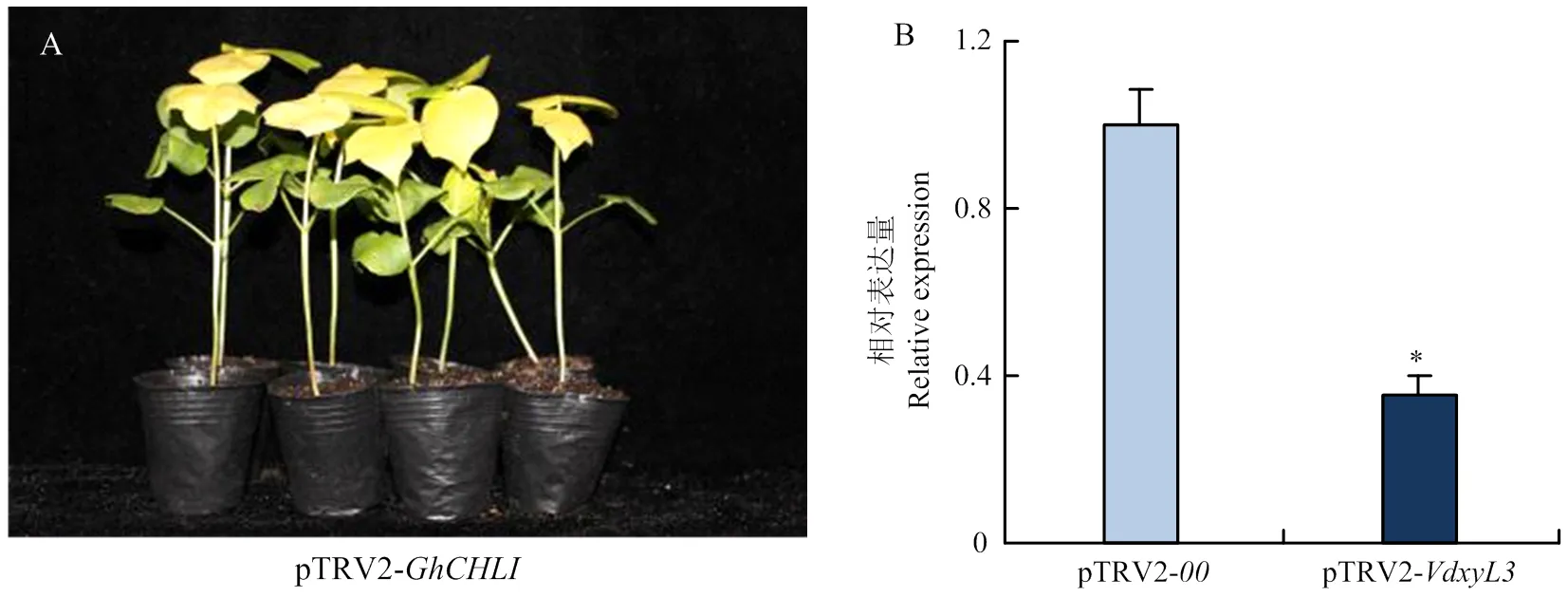
A:注射pTRV2-GhCHLI 10 d后感病品种新陆早8号的表型Phenotypes ofsusceptible variety Xinluzao 8 after 10 days of injection with pTRV2-GhCHLI;B:qRT-PCR 检测VdxyL3在pTRV2-00 和pTRV2-VdxyL3处理植株中的表达量。从接菌14 d后植株茎秆中提取棉花总RNA。tubulin为内参基因qRT-PCR analysis of VdxyL3 expression in the pTRV2-00 and pTRV2-VdxyL3 treated plants. Total RNA was isolated from stems at 14 dpi. tubulin was used as the control
2.8 病情调查
接菌后14 d和21 d,分别对pTRV2-和pTRV2-处理植株的发病情况进行调查。接菌后14 d,pTRV2-处理植株出现轻微的叶片黄化现象,而pTRV2-处理植株呈现明显的黄化和萎蔫现象;接菌后21 d,pTRV2-处理植株叶片出现明显的黄化、萎蔫和脱落现象,而pTRV2-处理植株发病严重,叶片脱落较多(图7-A)。两种处理植株的正常生长均受到影响,但pTRV2-处理植株的株高略矮于pTRV2-处理植株(图7-B)。病情指数调查发现pTRV2-处理植株在接菌14 d和21 d的病情指数(33.3和83.9)均显著高于pTRV2-植株14 d和21 d(21.7和66.1)(图7-C)。剖杆试验发现pTRV2-处理植株褐化程度明显较高(图7-D)。
2.9 大丽轮枝菌恢复培养和真菌相对生物量检测
接菌后14 d,分别采集pTRV2-和pTRV2-处理棉株的茎,提取总DNA,利用qRT-PCR检测真菌生物量。同时,收取pTRV2-和pTRV2-处理棉苗的茎用于大丽轮枝菌的恢复试验。大丽轮枝菌恢复培养和真菌生物量检测发现pTRV2-处理植株茎中的真菌生物量显著多于pTRV2-处理植株(图8-A、8-B),表明抑制的表达增强了大丽轮枝菌的致病力。
3 讨论
木糖苷酶基因对于降解植物细胞壁中的木聚糖至关重要,筛选大丽轮枝菌致病相关木糖苷酶基因,可为防治由该菌引起的植物黄萎病提供理论依据。木糖苷酶基因属于糖苷水解酶(glycoside hydrolases,GH)家族,该家族编码的酶能够以内切或外切的方式水解含糖化合物中的糖苷键。根据CAZy数据库依据氨基酸序列相似性对于糖苷水解家族的分类,共有168个公认的家族(GH1-168)。本研究从大丽轮枝菌中鉴定了13个木糖苷酶基因,它们分别属于GH3、GH31和GH43家族。真菌的-木糖苷酶大都属于GH3家族糖苷水解酶[24]。与前人研究不同,本研究13个基因中大多数(、、、、、、、和)属于GH43家族,仅有1个()属于GH3家族。有的-木糖苷酶是双功能酶,具有双重酶催化活性[25-26],本研究中和具有-木糖苷酶和阿拉伯糖苷酶双重酶活性,具有-葡糖苷酶/-木糖苷酶活性。-木糖苷酶在低聚木糖的末端还原端催化末端未经取代的木糖甙水解,13个基因中包括3个-木糖苷酶基因(、和),均属于GH31家族成员。

A:pTRV2-00和pTRV2-VdxyL3植株接种大丽轮枝菌14 d和21 d表型Disease symptom of the pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 and 21 dpi;B:接种大丽轮枝菌14 d后pTRV2-00和pTRV2-VdxyL3处理植株的株高Plant height of the pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 dpi;C:pTRV2-00和pTRV2-VdxyL3植株接种大丽轮枝菌14 d和 21 d病情指数统计Disease index of the pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 and 21 dpi;D:pTRV2-00和pTRV2-VdxyL3植株接种大丽轮枝菌14 d后茎秆的表型Disease symptom in stems of the pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 dpi

A:接种大丽轮枝菌14 d后pTRV2-00和pTRV2-VdxyL3处理植株茎秆中病原菌相对含量测定Quantification of the relative fungal biomass in stems of the pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 dpi;B:接种大丽轮枝菌14 d后pTRV2-00和pTRV2-VdxyL3处理植株茎秆大丽轮枝菌恢复培养。将茎秆置于PDA培养基,25℃培养箱中培养7 d后拍照Fungal isolation in the stem sections from pTRV2-00 and pTRV2-VdxyL3 treated plants at 14 dpi. Stems were plated on PDA medium. Photos were taken at 7 dpi of culture at 25℃
了解宿主与病原体的相互作用对于黄萎病的防控具有重要意义。在这个病理系统中,病害循环始于大丽轮枝菌微菌核对宿主根系分泌物刺激的响应和萌发[27]。感病品种根系分泌物促进大丽轮枝菌的生长,而抗病品种根系分泌物抑制其生长[28-29]。EI-Bebany等[27]研究发现,感病马铃薯品种根系分泌物诱导后大丽轮枝菌中差异基因的数量多于中抗品种根系分泌物诱导后的数量,高侵染力病原菌中受感病品种根系分泌物诱导表达的基因被认为与致病相关;Zhang等[21]研究发现,感病棉花品种根系分泌物诱导后大丽轮枝菌中差异基因的数量明显多于耐病和抗病品种根系分泌物诱导后的数量,一些编码水解酶和跨膜转运蛋白的基因受感病品种根系分泌物诱导后明显上调,被认为与致病相关;Xu等[9]研究表明,受棉花根诱导后表达量明显上调,敲除和回补试验发现该基因在大丽轮枝菌的产孢、菌丝体的发育、微菌核的形成和致病过程中发挥着重要作用,该基因HIGS沉默棉株的抗病性明显增强。上述研究表明棉花根系分泌物能够影响大丽轮枝菌中基因的表达,受其诱导表达的基因在大丽轮枝菌致病及宿主-病原体互作过程中可能发挥着重要的作用。本研究利用qRT-PCR对不同抗/感棉花品种根系分泌物诱导后大丽轮枝菌中6个木糖苷酶基因的表达量变化进行了分析,发现其表达均受根系分泌物的诱导,因此可将其作为研究大丽轮枝菌致病及宿主-病原体互作机制的候选基因。
大丽轮枝菌的致病分子机制非常复杂,近年来其致病相关基因的研究取得了阶段性的进展。大丽轮枝菌致病相关基因包括效应因子和细胞壁降解酶相关基因,如[30]、[8]、[31]和[32]等;生长-致病相关基因,如[33]、[34]、[35]和[36]等。尽管认为大丽轮枝菌分泌蛋白中的细胞壁降解酶能够参与植物细胞壁的降解,促进病原菌的定殖,但相关基因的鉴定及功能研究仅有少量报道。如果胶代谢相关基因和被敲除后,大丽轮枝菌的致病力降低[37]。调控果胶酶和半乳糖酶活性的被敲除后,大丽轮枝菌的致病力降低[31]。毒力因子与植物细胞壁降解有关,该基因的缺失突变体致病力降低[32]。赵玉兰等[38]利用HIGS技术快速筛选获得了4个细胞壁降解相关基因,转化4个靶标基因的烟草以及混合注射的烟草病情指数均降低,抗病性增强。上述研究表明将编码细胞壁降解酶的基因敲除或沉默导致大丽轮枝菌的致病力降低。本研究利用HIGS技术沉默后大丽轮枝菌的致病力明显增强,导致处理棉株的病情指数明显增高,这与前人细胞壁降解酶基因的功能研究结果不一致[31-32,37-38],推测可能参与诱导植物体内的抗性机制,增强植株的抗性,而该基因的沉默表达导致植株抗病性减弱。鉴于大丽轮枝菌致病分子机制的复杂性,的具体作用机制有待进一步研究。
4 结论
利用生物信息学方法从大丽轮枝菌基因组数据库中鉴定出13个木糖苷酶基因,包括9个糖苷水解酶43家族成员,1个3家族成员和3个31家族成员。6个木糖苷酶基因的表达均受到棉花根系分泌物的诱导,暗示其在大丽轮枝菌致病及宿主-病原体互作过程中可能发挥着重要的作用。通过HIGS技术对木糖苷酶基因在大丽轮枝菌侵染过程中的功能进行分析,发现当被干扰后,转化棉株的病情指数增高,抗病性减弱,表明与大丽轮枝菌的致病性相关。
[1] KLOSTERMAN S J, ATALLAH Z K, VALLAD G E, SUBBARAO K V. Diversity, pathogenicity, and management of Verticillium species. Annual Review of Phytopathology, 2009, 47: 39-62.
[2] 马存, 简桂良, 孙文姬. 我国棉花抗黄萎病育种现状、问题及对策. 中国农业科学, 1997, 30(2): 58-64.
MA C, JIAN G L, SUN W J. Current status, problem and countermeasure on resistance breeding to verticillium wilt of cotton in China. Scientia Agricultura Sinica, 1997, 30(2): 58-64. (in Chinese)
[3] 张绪振, 张树琴, 陈吉棣, 李庆基, 陈壁, 姚跃文. 我国棉花黄萎病菌“种”的鉴定. 植物病理学报, 1981, 11(3): 13-20.
ZHANG X Z, ZHANG S Q, CHEN J D, LI Q J, CHEN B, YAO Y W. Identification of verticillium wilt pathogen of cotton in China. Acta Phytopathologica Sinica, 1981, 11(3): 13-20. (in Chinese)
[4] GERIK J S, HUISMAN O C. Study of field-grown cotton roots infected withusing an immunoenzymatic staining technique. Phytopathology, 1988, 78(9): 1174-1178.
[5] FRADIN E F, THOMMA B P H J. Physiology and molecular aspects of verticillium wilt diseases caused byand. Molecular Plant Pathology, 2006, 7(2): 71-86.
[6] GUI Y J, CHEN J Y, ZHANG D D, LI N Y, LI T G, ZHANG W Q, WANG X Y, SHORT D P, LI L, GUO W, KONG Z Q, BAO Y M, SUBBARAO K V, DAI X F.manipulates plant immunity by glycoside hydrolase 12 proteins in conjunction with carbohydrate-binding module 1. Environmental Microbiology, 2017, 19(5): 1914-1932.
[7] QIN J, WANG K L, SUN L F, XING H Y, WANG S, LI L, CHEN S, GUO H S, ZHANG J. The plant-specific transcription factors CBP60G and SARD1 are targeted by asecretory protein VDSCP41 to modulate immunity. eLife, 2018, 7: e34902.
[8] ZHANG L S, NI H, DU X, WANG S, MA X W, NRNBERGER T, GUO H S, HUA C L. The-specific protein VdSCP7 localizes to the plant nucleus and modulates immunity to fungal infections. New Phytologist, 2017, 215(1): 368-381.
[9] XU J, WANG X Y, LI Y Q, ZENG J G, WANG G L, DENG C Y, GUO W Z. Host-induced gene silencing of a regulator of G protein signaling gene () confers resistance to verticillium wilt in cotton. Plant Biotechnology Journal, 2018, 16(9): 1629-1643.
[10] ZHAO Y L, ZHANG T, GUO H S. Penetration assays, fungal recovery and pathogenicity assays for. Bio-Protocol, 2017, 7(4): DOI: 10.21769/BioProtoc.2133.
[11] HOGENHOUT S A, VAN DER HOORN R A, TERAUCHI R, KAMOUN S. Emerging concepts in effector biology of plant- associated organisms. Molecular Plant-Microbe Interactions, 2009, 22(2): 115-122.
[12] 张志东. 棉花黄萎病致病相关基因的挖掘与功能分析[D] . 上海: 上海交通大学, 2017.
ZHANG Z D. Mining and function analysis of genes related to cotton verticillium wilt[D]. Shanghai: Shanghai jiaotonguniversity, 2017. (in Chinese)
[13] HUANG X L, LI Z, DU C Y, WANG J F, LI S. Improved expression and characterization of a multidomain xylanase fromSCUT27 in. Journal of Agricultural and Food Chemistry, 2015, 63(28): 6430-6439.
[14] ZHANG S Y, WANG H M, SHI P J, XU B, BAI Y G, LUO H Y, YAO B. Cloning, expression, and characterization of a thermostable-xylosidase from thermoacidophilicsp. A4. Process Biochemistry, 2014, 49(9): 1422-1428.
[15] NGUYEN Q B, ITOH K, VU B V, TOSA Y, NAKAYASHIKI H. Simultaneous silencing of endo--1,4 xylanase genes reveals their roles in the virulence of. Molecular Microbiology, 2011,81(4):1008-1019.
[16] BritoN, Espino J J, GonzálezC. The endo--1,4-xylanase Xyn11A is required for virulence in. Molecular Plant-Microbe Interactions, 2006, 19(1):25-32.
[17] MUSSATTO S I, MANCILHA I M. Non-digestible oligosaccharides: A review. Carbohydrate Polymers, 2007, 68(3): 587-597.
[18] VAN RENSBURG P, STRAUSS M L A, LAMBRECHTS M G, CORDERO OTERO R R, PRETORIU I S. The heterologous expression of polysaccharidase-encoding genes with oenological relevance in. Journal of Applied Microbiology, 2007, 103(6): 2248-2257.
[19] BOSETTO A, JUSTO P I, ZANARDI B, VENZON S S, GRACIANO L, DOS SANTOS E L, DE CASSIA GARCIA SIMAO R. Research progress concerning fungal and bacterial-xylosidases. Applied Biochemistry and Biotechnology, 2016, 178(4): 766-795.
[20] MUSTAFA G, KOUSAR S, RAJOKA M I, JAMIL A. Molecular cloning and comparative sequence analysis of fungal-xylosidases. AMB Express, 2016, 6(1): 30.
[21] ZHANG X Y, CHENG W H, FENG Z D, ZHU Q H, SUN Y Q, LI Y J, SUN J. Transcriptomic analysis of gene expression ofupon treatment of the cotton root exudates. BMC Genomics, 2020, 21(1): 155.
[22] 熊显鹏.和在棉花抗黄萎病中的功能研究[D]. 石河子: 石河子大学, 2020.
XIONG X P. Functional analysis ofand
[23] ELLENDORFF U, FRADIN E F, DE JONGE R, THOMMA B P. RNA silencing is required fordefence against verticillium wilt disease.Journal of Experimental Botany, 2009, 60(2): 591-602.
[24] YANG X Z, SHI P J, HUANG H Q, LUO H Y, WANG Y R, ZHANG W, YAO B. Two xylose-tolerant GH43 bifunctional-xylosidase/-arabinosidases and one GH11 xylanase fromand their synergy in the degradation of xylan. Food Chemistry, 2014, 148: 381-387.
[25] JORDAN D B, LI X L. Variation in relative substrate specificity of bifunctional-d-xylosidase/-l-arabinofuranosidase by single-site mutations: roles of substrate distortion and recognition. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics, 2007, 1774(9): 1192-1198.
[26] LEE R C, HRMOVA M, BURTON R A, LAHNSTEIN J, FINCHER G B. Bifunctional family 3 glycoside hydrolases from barley with-l-arabinofuranosidase and-D-xylosidase activity. Journal of Biological Chemistry, 2003, 278(7): 5377-5387.
[27] El-BEBANY A F, HENRIQUEZ M A, BADAWI M, ADAM L R, HADRAMI A E, DAAYF F. Induction of putative pathogenicity- related genes inin response to elicitation with potato root extracts. Environmental and Experimental Botany, 2011, 72(2):251-257.
[28] 袁虹霞, 李洪连, 王烨, 房卫平, 王振跃. 棉花不同抗性品种根系分泌物分析及其对黄萎病菌的影响. 植物病理学报, 2002, 32(2): 127-131.
YUAN H X, LI H L, WANG Y, FANG W P, WANG Z Y. The root exudates of cotton cultivars with the different resistance and their effects on. Acta Phytopathologica Sinica, 2002, 32(2): 127-131. (in Chinese)
[29] 郑倩, 李俊华, 危常州, 褚贵新. 不同抗性棉花品种根系分泌物及酚酸类物质对黄萎病菌的影响. 棉花学报, 2012, 24(4): 363-369.
ZHENG Q, LI J H, WEI C Z, CHU G X. Effects of root exudates and phenolic acids from differently resistant cotton cultivars on. Cotton Science, 2012, 24(4): 363-369. (in Chinese)
[30] KOMBRINK A, ROVENICH H, SHI-KUNNE X, ROJAS-PADILLA E, DOMAZAKIS E, VAN DEN BERG G C M, Domazakis E, DE JONGE R, VALKENBURG D J, SANCHEZ-VALLET A, SEIDL M F, THOMMA B P H J.LysM effectors differentially contribute to virulence on plant hosts. Molecular Plant Pathology, 2017, 18(4): 596-608.
[31] TZIMA A K, PAPLOMATAS E J, RAUYAREE P, OSPINA- GIRALDO M D, KANG S., the sucrose non-fermenting protein kinase gene of, is required for virulence and expression of genes involved in cell-wall degradation. Molecular Plant-Microbe Interactions, 2011, 24(1): 129-142.
[32] LIU S Y, CHEN J Y, WANG J L, LI L, XIAO H L, ADAM S M, DAI X F. Molecular characterization and functional analysis of a specific secreted protein from highly virulent defoliating. Gene, 2013, 529(2): 307-316.
[33] SANTHANAM P, THOMMA B P.Sge1 differentially regulates expression of candidate effector genes. Molecular Plant-Microbe Interactions, 2013, 26(2): 249-256.
[34] LI Z F, LIU Y J, FENG Z L, FENG H J, KLOSTERMAN S J, ZHOU F F, ZHAO L H, SHI Y Q, ZHU H Q., encoding CYC8 glucose repression mediator protein, is required for microsclerotia formation and full virulence in. PLoS One, 2015, 10(12): e0144020.
[35] ZHANG Y L, LI Z F, FENG Z L, FENG H J, SHI Y Q, ZHAO L H, ZHANG X L, ZHU H Q. Functional analysis of the pathogenicity- related gene. PLoS One, 2016, 11(11): e0166000.
[36] QI X L, SU X F, GUO H M, QI J C, CHENG H M. VdThit, a thiamine transport protein, is required for pathogenicity of the vascular pathogen. Molecular Plant-Microbe Interactions, 2016, 29(7): 545-559.
[37] CHEN J Y, XIAO H L, GUI Y J, ZHANG D D, LI L, BAO Y M, DAI X F. Characterization of theexoproteome involves in pathogenicity from cotton-containing medium. Frontiers in Microbiology, 2016, 7: 1709.
[38] 赵玉兰, 苏晓峰, 程红梅. 利用寄主诱导的基因沉默技术验证大丽轮枝菌糖代谢相关基因的致病力. 中国农业科学, 2015, 48(7): 1321-1329.
ZHAO Y L, SU X F, CHENG H M. Verification ofpathogenicity of glycometabolism related genes by using host-induced gene silencing method. Scientia Agricultura Sinica, 2015, 48(7): 1321-1329. (in Chinese)
Identification of Xylosidase Genes fromand Functional Analysis Based on HIGS Technology
ZHANG XiaoXue, SUN TianGe, ZHANG YingChun, CHEN LiHua, ZHANG XinYu, LI YanJun, SUN Jie
College of Agriculture, Shihezi University, Shihezi 832003, Xinjiang
【】The objective of this research is to identify xylosidase genes fromand study the relationship between xylosidase genes and pathogenicity of, which will provide a theoretical basis for exploring the molecular mechanism of pathogenicity ofand a scientific basis for formulating better control strategies for verticillium wilt.【】All xylosidase genes were identified from the genome database ofby bioinformatics, and their protein domain, chromosomal location and phylogenetic relationship were analyzed. Quantitative real-time PCR (qRT-PCR) was used to detect the expression pattern of xylosidase genes inculturedwith different root exudates from resistant and susceptible cotton varieties for 0, 6, 12, 24 and 48 h. The function of one of the xylosidase geneswas analyzed by using host-induced gene silencing (HIGS) method. The target fragment ofwas injected into cotton, and thenVd991 was inoculated to those plants injected with target fragment ofby using root-dip approach. The phenotype of transformed plants was observed and the disease index was counted, meanwhile, the biomass of fungi and the expression level of【】Bioinformatics analysis showed that there were 13 xylosidase genes (-) in, whose coding sequences ranged from 1 461 to 2 544 bp, molecular weight of the encoded proteins ranged from 38.78 to 90.97 kD, and theoretical isoelectric point ranged from 4.67 to 5.89. Protein domain phylogenetic relationship analysis showed that there were 9 glycoside hydrolase 43 family members, 1 glycoside hydrolase 3 family member and 3 glycoside hydrolase 31 family members included in xylosidase genes. The chromosomal location analysis showed that the 13 genes were distributed on 6 chromosomes and no gene clusters were formed. qRT-PCR analysis showed that the expression of 6 xylosidase genes was induced by cotton root exudates. After being cultured in one or more root exudates for 6 h or 12 h, expression levels of these genes were significantly increased and then decreased. Among 6 genes, the expression level ofincreased significantly after sensing root exudates from sea island cotton. The results based on HIGS technology showed that after 14 and 21 days of inoculation, the disease symptom of cotton plants transformed withinterfering fragment was more serious, and the disease index (33.3 and 83.9) was significantly higher than that of empty vector control (21.7 and 66.1). qRT-PCR analysis showed that cotton plants transformed withinterfering fragment had higher fungal biomass but lower expression level ofcompared to the empty vector control.【】Thegene silencing by using HIGS technology lead to a significant decrease of cotton resistance to, indicating thatmay play a certain role in the pathogenesis ofand host-pathogen interaction.
cotton;; verticillium wilt;; xylosidase; host-induced gene silencing (HIGS)

10.3864/j.issn.0578-1752.2021.15.007
2020-11-09;
2020-12-18
国家自然科学基金(31460360)、石河子大学高层次人才科研启动项目(RCZK202019)
张小雪,E-mail:943124507@qq.com。通信作者李艳军,E-mail:lyj20022002@sina.com
(责任编辑 岳梅)
猜你喜欢
中草药(2022年20期)2022-11-15
食品科学(2022年20期)2022-10-31
现代食品科技(2022年9期)2022-10-09
湖北农业科学(2022年11期)2022-07-18
中国土壤与肥料(2021年5期)2021-12-02
实用肿瘤学杂志(2020年4期)2020-12-08
伴侣(2019年5期)2019-05-27
人人健康(2017年23期)2017-12-26
食品工业科技(2014年23期)2014-03-11
医学综述(2011年12期)2011-12-09
