
lncRNA TMPO-AS1靶向miR-1224-5p调控肝癌细胞的增殖和凋亡
2021-08-18 02:14金春英
河北医药 2021年15期
金春英
肝癌被列为全球第四大最常见的恶性肿瘤和第三大癌症相关死亡原因[1]。尽管肝癌的治疗方法取得了长足的进步,包括肝切除、化学疗法和放射疗法或分子靶向疗法,但肝癌患者的预后仍然很差[2]。因此,迫切需要了解肝癌发生的分子机制,以探索新型肝癌预后生物标志物,促进肝癌患者治疗策略的发展。长链非编码RNA(long non-coding RNA,lncRNA)是具有200个以上核苷酸的非蛋白质RNA[3]。越来越多的证据表明,lncRNA在病理过程扮演关键角色,包括癌症的发生和发展[4]。lncRNA TMPO antisense RNA 1(TMPO-AS1)在膀胱癌组织和细胞中高表达,促进膀胱癌细胞的生长、迁移和侵袭[5]。TMPO-AS1有助于肺癌发生,这可能是通过上调TMPO引起的[6]。但lncRNA TMPO-AS1在肝癌进展中的确切功能和潜在分子机制需要进一步探索。微小RNA(microRNA,miRNA)是约22个核苷酸的小型非编码RNA,在癌症的发生和发展中起着至关重要的作用。最近研究表明,microRNA-1224-5p(miR-1224-5p)的异常表达与几种人类癌症有关,例如miR-1224-5p在骨肉瘤组织和细胞中低表达[7],可能是前列腺癌的肿瘤抑制因子[8]。Chao等[9]学者报告指出,miR-1224-5p表达的上调可能会减少人肝癌肿瘤内皮细胞的增殖,诱导凋亡,抑制迁移和侵袭并抑制管形成,并可能在肝癌中起潜在的肿瘤抑制作用。至于miR-1224-5p在肝癌中的表达和功能仍然不清楚。本研究针对lncRNA TMPO-AS1、miR-1224-5p在肝癌组织中的表达和对肝癌细胞增殖、凋亡的影响进行探讨,并分析它们之间的靶向调控,为肝癌发病机制提供新的见解。
1 材料与方法
1.1 细胞与试剂 肝癌细胞株HCCLM3购自美国典型培养物保藏中心,Roswell Park Memorial Institute(RPMI-1640)培养基购自美国Gibco公司,Lipofectamine 2 000购自Thermo Fisher Scientific公司,Prime Script TM RT Master Mix试剂盒、Prime Script TM RT Master Mix试剂盒购自大连TaKaRa公司,噻唑蓝(methyl thiazolyl tetrazolium,MTT)购自美国Sigma公司,FITC Annexin V细胞凋亡检测试剂盒购自美国BD Biosciences公司,RIPA缓冲液、二辛可宁酸(bicinchoninic acid,BCA) 蛋白质分析试剂盒、SDS-PAGE凝胶购自上海Beyotime公司,抗细胞周期蛋白D1(CyclinD1)、抗p21、抗B细胞淋巴瘤/白血病-2(B cell lymphoma/lewkmia-2,Bcl-2)、抗Bcl-2相关X蛋白(Bcl-2 Associated X Protein,Bax)和抗甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体购自美国Abcam公司。
1.2 临床组织标本 2016年2月至2018年8月从本院的肝癌手术中获得了35对肝癌组织和相邻的癌旁组织样本(距离肿瘤组织>3 cm)。这项研究的所有肿瘤组织均经病理学确诊,并获得患者的同意。切除后将组织在液氮中冷冻,直至用于实时荧光定量PCR(qRT-PCR)测定。
1.3 细胞培养与转染 将HCCLM3细胞培养于RPMI-1640培养基中,向其中添加10%胎牛血清、100 U/ml青霉素和100 mg/ml链霉素。在37℃,含有5% CO2的培养箱中培养。lncRNA TMPO-AS1过表达质粒(pcDNA-TMPO-AS1),pcDNA质粒(pcDNA),针对lncRNA TMPO-AS1的小干扰RNA(si-TMPO-AS1),小干扰RNA阴性对照(si-NC),miR-1224-3p模拟物(miR-1224-3p),模拟物阴性对照(miR-NC),miR-1224-3p抑制剂(anti-miR-1224-5p),抑制剂阴性对照(anti-miR-NC)由苏州GENEWIZ公司合成。使用Lipofectamine 2000用上述质粒或寡核苷酸转染HCCLM3细胞(接种于6孔板中,融合度为70%)。将细胞分为pcDNA组(转染pcDNA),pcDNA-TMPO-AS1组(转染pcDNA-TMPO-AS1),si-NC组(转染si-NC),si-TMPO-AS1组(转染si-TMPO-AS1),miR-NC组(转染miR-NC),miR-1224-3p组(转染miR-1224-3p),si-TMPO-AS1+anti-miR-NC组(共转染si-TMPO-AS1和anti-miR-NC),si-TMPO-AS1+anti-miR-1224-5p组(共转染si-TMPO-AS1和anti-miR-1224-5p)。转染48 h后检测各项指标。
1.4 qRT-PCR测定肝癌组织和细胞中lncRNA TMPO-AS1和miR-1224-5p表达 使用TRIzol试剂从肝癌组织、癌旁组织和HCCLM3细胞。使用Prime Script TM RT Master Mix进行RNA逆转录,以得到的互补DNA(cDNA)为模板,使用SYBR Premix Ex Taq II于Applied Biosystems 7500实时PCR系统进行qPCR。根据2-ΔΔCt方法分析lncRNA TMPO-AS1和miR-1224-5p表达。引物如下:lncRNA TMPO-AS1 5,-AGCCCACACACTACAGGCA-3,(F)和5,-GCACAAAAGCAGTACGACCT-3,(R)。内参β肌动蛋白(β-actin) 5,-CCTCTCCCAAGTCCACACAG-3,(F)和5,-GGGCACGAAGGCTCATCATT-3,(R)。miR-1224-5p 5,-TGCGCGTGAGGACTCGGGA-3,(F)和5,-CTCAAGTGTCGTGGAGTCGGCAA-3,(R)。内参U6 5,-CGCTTCGGCAGCACATATAC-3,(F)和5,-AAATATGGAACGCTTCACGA-3,(R)。
1.5 双荧光素酶报告实验测定肝癌HCCLM3细胞荧光素酶活性 将包含假定miR-1224-5p结合位点的HCCLM3野生型(WT)片段或突变(MUT)序列插入pmirGLO双荧光素酶载体中,分别命名为WT-HCCLM3或MUT-HCCLM3。使用Lipofectamine 2000,在HCCLM3细胞中共转染WT-HCCLM3或MUT-HCCLM3和miR-1224-5p模拟物或miR-NC。转染后48 h收获细胞。根据制造商的说明,使用双荧光素酶报告基因检测系统检测荧光素酶活性。
1.6 MTT比色法测定肝癌HCCLM3细胞增殖 用上述质粒或寡核苷酸转染的HCCLM3细胞分别在96孔板中接种24 h、48 h和72 h,加入20 μl/孔的MTT溶液(5 mg/ml),并继续温育4 h。然后,除去上清液,并将反应得到的甲瓒晶体溶解在200 μl的二甲基亚砜中。晶体在室温下溶解10 min。通过使用Bio Tek酶标仪测量450 nm的吸光度(OD)来评估细胞增殖。
1.7 流式细胞术测定肝癌HCCLM3细胞凋亡 使用FITC Annexin V细胞凋亡检测试剂盒分析细胞凋亡。收集用上述质粒或寡核苷酸转染的HCCLM3细胞,在室温下黑暗中用10 μl Annexin V-FITC和5 μl碘化丙啶处理细胞15 min。通过具有Cell Quest软件的FACSCalibur流式细胞仪检测细胞凋亡率。
1.8 免疫印迹实验(western blot)测定肝癌HCCLM3中CyclinD1、p21、Bcl-2和Bax蛋白表达 用上述质粒或寡核苷酸处理的HCCLM3细胞在RIPA缓冲液中裂解。使用BCA 蛋白质分析试剂盒对蛋白裂解物进行定量,然后通过SDS-PAGE进行分离。将蛋白转移到聚偏二氟乙烯膜中,然后用5%脱脂牛奶封闭。将膜与一抗在4℃孵育过夜,与以辣根过氧化物酶(HRP)标记的二抗(1∶5 000)孵育2 h。一抗包括抗CyclinD1(1∶1 000)、抗p21(1∶1 000)、抗Bcl-2(1∶1 000)、抗Bax(1∶1 000)和抗GAPDH(1∶2 000)。通过增强的化学发光系统观察印迹。以GAPDH为对照,分析蛋白的表达水平。

2 结果
2.1 lncRNA TMPO-AS1和miR-1224-5p在肝癌组织中的表达 与癌旁组织比较,肝癌组织中lncRNA TMPO-AS1的表达有所增加,miR-1224-5p的表达明显降低,差异有统计学意义(P<0.05)。见表1。

表1 lncRNA TMPO-AS1 和miR-1224-5p 在肝癌组织中的表达
2.2 lncRNA TMPO-AS1靶向调控miR-1224-5p的表达 采用StarBase工具(http://starbase.sysu.edu.cn/)预测lncRNA TMPO-AS1与miR-1224-5p之间的相互作用,发现miR-1224-5p可以作为lncRNA TMPO-AS1的靶基因,双荧光素酶报告基因检测结果显示,与miR-NC组比较,含有WT-TMPO-AS1的荧光素酶报告质粒与miR-1224-5p在肝癌HCCLM3细胞中共转染降低了荧光素酶活性(P<0.05),而含有MUT-TMPO-AS1的荧光素酶报告质粒与miR-1224-5p共转染,荧光素酶活性差异无统计学意义(P>0.05)。qRT-PCR数据发现,miR-1224-5p在pcDNA-TMPO-AS1组的表达水平低于pcDNA组,si-TMPO-AS1组的表达水平高于si-NC组(P<0.05)。见图1,表2、3。

图1 lncRNA TMPO-AS1的序列中含有与miR-1224-5p互补的核苷酸序列

表2 双荧光素酶报告实验
表3 lncRNA TMPO-AS1 调控miR-1224-5p 的表达
2.3 敲低lncRNA TMPO-AS1表达对肝癌HCCLM3细胞增殖和凋亡的影响 相较于转染si-NC,肝癌HCCLM3细胞中转染si-TMPO-AS1后,lncRNA TMPO-AS1的表达被抑制(P<0.05),细胞24 h、48 h和72 h的OD值明显降低,凋亡率显著升高,并且CyclinD1蛋白和Bcl-2蛋白表达水平明显减少,p21蛋白和Bax蛋白表达水平显著增加,差异有统计学意义(P<0.05)。见图2,表4。

图2 敲低lncRNA TMPO-AS1表达对肝癌HCCLM3细胞增殖和凋亡的影响;A 增殖和凋亡相关蛋白表达;B 细胞凋亡流式图
表4 敲低lncRNA TMPO-AS1 表达对肝癌HCCLM3 细胞增殖和凋亡的影响
2.4 抑制miR-1224-5p表达逆转了敲低lncRNA TMPO-AS1表达对肝癌HCCLM3细胞增殖和凋亡的作用 与si-NC组比较,si-TMPO-AS1组肝癌HCCLM3细胞的miR-1224-5p表达被促进,细胞48 h和72 h的OD值明显减少,凋亡率有所增加,CyclinD1蛋白和Bcl-2蛋白表达水平显著降低,p21蛋白和Bax蛋白表达水平明显升高,差异有统计学意义(P<0.05)。与si-TMPO-AS1+anti-miR-NC组比较,si-TMPO-AS1+anti-miR-1224-5p组肝癌HCCLM3细胞的miR-1224-5p表达被抑制,细胞24 h、48 h和72 h的OD值明显增加,凋亡率显著减少,CyclinD1蛋白和Bcl-2蛋白表达水平显著升高,p21蛋白和Bax蛋白表达水平明显降低,差异有统计学意义(P<0.05)。见图3,表5。

图3 抑制miR-1224-5p表达逆转了敲低lncRNA TMPO-AS1表达对肝癌HCCLM3细胞增殖和凋亡的作用;A 增殖和凋亡相关蛋白表达;B 细胞凋亡流式图
表5 抑制miR-1224-5p 表达逆转了敲低lncRNA TMPO-AS1 表达对肝癌HCCLM3 细胞增殖和凋亡的作用
2.5 miR-1224-5p过表达对肝癌HCCLM3细胞增殖和凋亡的影响 肝癌HCCLM3细胞转染miR-1224-3p后,同转染miR-NC组比较,miR-1224-3p组大幅增加miR-1224-3p的表达水平(P<0.05),明显减少细胞24 h、48 h和72 h的OD值,显著增加细胞的凋亡率,以及明显降低CyclinD1蛋白和Bcl-2蛋白表达水平,显著升高p21蛋白和Bax蛋白表达水平,差异有统计学意义(P<0.05)。见图4,表6。
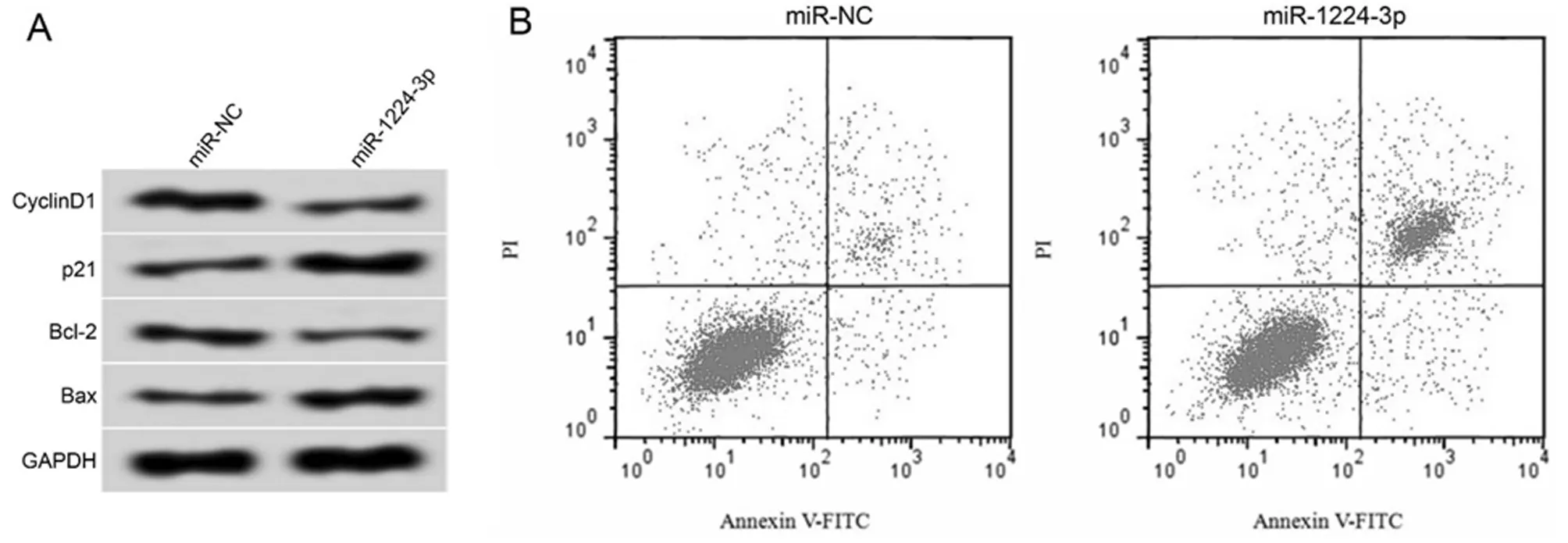
图4 miR-1224-5p过表达对肝癌HCCLM3细胞增殖和凋亡的影响;A 增殖和凋亡相关蛋白表达;B 细胞凋亡流式图
表6 miR-1224-5p 过表达对肝癌HCCLM3 细胞增殖和凋亡的影响
3 讨论
越来越多的证据表明,包括lncRNA和miRNA在内的非编码RNA是肝癌发展中的重要调节剂,具有潜在的诊断和治疗价值[10]。本研究旨在探讨lncRNA TMPO-AS1在肝癌细胞中的作用和机制。结果显示,在肝癌组织中,lncRNA TMPO-AS1被上调,而miR-1224-5p被下调。敲低lncRNA TMPO-AS1表达或miR-1224-5p过表达可以抑制肝癌进程。值得注意的是,miR-1224-5p是lncRNA TMPO-AS1的靶标。
近年来,lncRNA被公认为包括肝癌在内的重要癌症生物标志物。因此,鉴定肝癌预后的lncRNA至关重要。根据报道lncRNA TMPO-AS1是一种与癌症相关的lncRNA,在多种癌症中充当癌基因,例如,lncRNA TMPO-AS1在胃癌组织中表达上调,上调其表达可促进胃癌细胞的增殖、迁移和侵袭[11]。非小细胞肺癌组织和细胞中的lncRNA TMPO-AS1过表达,TMPO-AS1的下调在体外显著抑制细胞增殖,集落形成,迁移和侵袭以及体内肿瘤生长[6]。TMPO-AS1可能是前列腺癌的有用诊断和预后标志物,TMPO-AS1过表达通过促进细胞周期进程和促进迁移而增加了前列腺癌细胞增殖,但降低了细胞的凋亡[12]。lncRNA TMPO-AS1在宫颈癌细胞中表达上调,上调其表达可促进宫颈癌细胞的增殖、迁移和侵袭[13]。在当前的研究中,为了解lncRNA TMPO-AS1在肝癌中的作用,通过qRT-PCR分析检测了肝癌组织及其相应的癌旁组织中lncRNA TMPO-AS1的表达水平。结果观察到,与癌旁组织相比,肝癌组织中lncRNA TMPO-AS1表达水平显著增加。为了调查肝癌中lncRNA TMPO-AS1表达的生物学功能,利用小干扰RNA敲低lncRNA TMPO-AS1的表达,结果发现,其敲低抑制了肝癌细胞的增殖和增殖相关蛋白CyclinD1、抗凋亡蛋白Bcl-2的表达,促进了细胞的凋亡、增殖相关蛋白p21和促凋亡蛋白Bax的表达。表明与前述研究一致,lncRNA TMPO-AS1在肝癌中起着致癌lncRNA的作用。
miR-1224-5pw位于人类3q27.1染色体,可抑制多种癌症的发展,包括黑色素瘤[14]和结直肠癌[15]。资料显示,miR-1224-5p在胶质瘤组织中低表达,具有抑制胶质瘤细胞增殖的作用[16]。上调miR-1224-5p的表达可抑制人肝癌血管内皮细胞的增殖能力,促进肝癌血管内皮细胞的凋亡并降低其迁移、侵袭及血管形成能力[17]。目前的研究证明,miR-1224-5p在肝癌组织中显著下调。miR-1224-5p过表达可降低细胞增殖能力、CyclinD1和Bcl-2蛋白表达,并诱导细胞凋亡、p21和Bax蛋白表达。这些结果表明,miR-1224-5p在肝癌中发挥了抑癌作用,与前述报道[7-9]相符。
作为基因调节剂,lncRNA对生物过程的调控具有多种分子机制,例如RNA-RNA,RNA-蛋白质和RNA-DNA相互作用[18],其主要机制之一是作为miRNA海绵来调控miRNA的表达[19]。在这项研究中,生物信息学分析表明lncRNA TMPO-AS1和miR-1224-5p互补结合位点。荧光素酶测定法证实miR-1224-5p是lncRNA TMPO-AS1的直接靶标,并且,qRT-PCR检测到上调lncRNA TMPO-AS1时miR-1224-5p的表达受到抑制,下调lncRNA TMPO-AS1时其表达被促进,证明lncRNA TMPO-AS1可以靶向负调控miR-1224-5p的表达。进一步探索lncRNA TMPO-AS1在肝癌中发挥作用的机制发现,抑制miR-1224-5p表达能逆转敲低lncRNA TMPO-AS1表达抑制肝癌HCCLM3细胞增殖的作用,并逆转敲低lncRNA TMPO-AS1表达促进细胞凋亡的作用。证实靶向调控miR-1224-5p是lncRNA TMPO-AS1影响肝癌发生发展的重要途径之一。
总之,lncRNA TMPO-AS1在肝癌组织中表达上调,发挥肿瘤促进作用。敲低lncRNA TMPO-AS1的表达可以抑制肝癌细胞的增殖,和诱导细胞凋亡。此外,lncRNA TMPO-AS1的作用机制与靶向miR-1224-5p有关,这提示lncRNA TMPO-AS1可能成为肝癌的候选生物标志物。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
天津医科大学学报(2021年4期)2021-08-21
江西农业学报(2021年4期)2021-04-20
中日友好医院学报(2021年1期)2021-04-14
昆明医科大学学报(2021年2期)2021-03-29
广东蚕业(2021年1期)2021-03-18
三农资讯半月报(2020年11期)2020-06-21
