
凤阳山常绿阔叶林优势种空间格局及关联性
2021-08-17 01:21季新良周红敏夏春平张弓乔
中国野生植物资源 2021年7期
季新良,彭 辉,周红敏,夏春平,瞿 虹,张弓乔
(1. 浙江凤阳山自然保护区管理处,浙江 龙泉 323700; 2. 浙江龙泉市林科院,浙江 龙泉 323700; 3. 龙泉市林业局,浙江 龙泉 323700;4. 中国林业科学研究院林业所,北京100091)
群落组成与空间格局为研究物种共存机制提供了重要信息,是研究种群特征、种群间相互作用及种群与环境关系的重要基础[1]。种群空间格局主要受到种内种间竞争及环境过滤等多因素共同作用[2],从物种空间分布格局推测多样性的维持机制即从格局推导的过程,是生态学研究的重要方面[3]。森林中树种的空间格局及关联性对树种生长、死亡、繁殖、资源利用及对干扰的抵抗有显著的影响,特别是研究不同树种空间关联性,可以更好的解析该群落形成和维持机制。大量研究表明,树种的空间格局与尺度密切关联,例如树种在小尺度上呈现聚集分布,而在大尺度上则可能为随机分布或均匀分布[4]。
点格局分析方法如Ripley’sK函数以树种分布的空间坐标为基础将个体视为2维平面的一个点[5],可分析不同尺度下的种群格局及种间(种内)关系[6]。但因其具有累积性,会混淆大、小尺度的效应。双相关函数g(r)是K函数的衍生,因其能有效剔除K函数在小尺度上的累积效应,在国内外已有大量研究应用,涵盖了如热带天然针叶林[7]、亚热带常绿落叶阔叶混交林[8]、温带落叶阔叶林[9]等类型,发表了大量的文献取得了众多成果[10]。
凤阳山国家级自然保护区位于浙江省南部,其地带性植被为亚热带常绿阔叶林,因海拔高度的变化,在相应的气候垂直分布带上形成森林植被的垂直带谱系列[11],加之地形和人为活动影响,出现了如针叶林、针阔混交林、常绿落叶阔叶混交林、山地矮林等多种类型的植被[12]。但目前关于凤阳山保护区常绿阔叶林优势树种的空间分布和空间关联的研究尚不多见。因此,依托凤阳山0.8 hm2常绿阔叶林固定监测样地,本研究分析其主要空间优势树种的空间分布格局及其关联性,旨在探讨该区域种群空间分布特点,推断群落演替方向,为深入研究凤阳山保护区生物多样性和物种共存提供参考。
1 研究地区与研究方法
1.1 研究区概况
凤阳山自然保护区位于浙江省龙泉市南部(119°06′~119°15′E、27°46′~27°58′N),面积151 km2,属于武夷山系洞宫山山脉的中段,悬崖峭壁多处可见。土壤以黄壤土为主,地带性植被为典型的中亚热带常绿阔叶林,且植被垂直分布明显。区内年平均气温12.3℃,年日照1 515.5 h,平均相对湿度80%,年蒸发量1 171.0 mm,年降雨量2 438.2 mm。有种子植物164科、659属、1 488种,以栲属(Castanopsis)、木荷属(Schima)和青冈属(Cyclobalanopsis)为主要优势树种,分布着红豆杉(Taxuschinensis)、伯乐树(Bretschneiderasinensis)、南方红豆杉(Taxuschinensisvar.mairei)等珍贵树种[13]。
1.2 数据采集
在前期全面实地踏查基础上,选择典型的常绿阔叶林群落,依照史密森热带森林研究中心(Center for tropical forest science,CTFS)林生物多样性监测规范设立80 m×100 m的固定样地(坡度20°,海拔1 300 m)划分为5 m×5 m的小样方,调查胸径(Diameter at breast height,DBH)≥1 cm的所有存活木本植物个体,挂牌并记录物种名称、坐标(x,y)、胸径、树高、冠幅等。
1.3 点格局分析
以恵刚盈提出的树种优势度(DSP)替代重要值[14],对样地内的树种空间优势度进行比较:
(1)
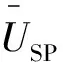
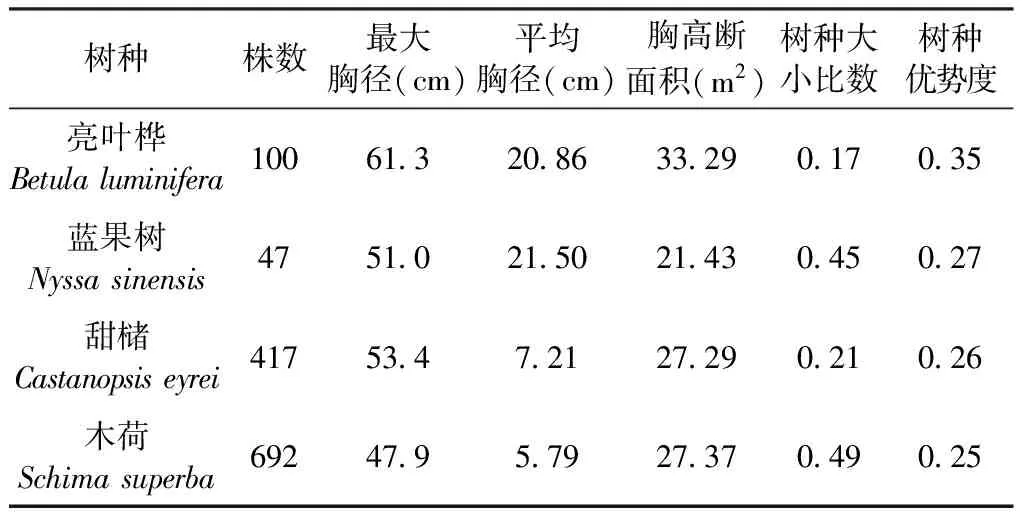
表1 样地主要空间优势树种Table 1 Dominant species in the forest plot
由于亮叶桦和蓝果树株数较少,而点格局分析对样本量有一定要求,因此采用单变量双相关函数g(r)分析4个优势树种整体空间格局,采用双变量双相关函数g12(r)分析4优势树种两两之间的空间关联性[15-16],而不对各个龄级进行空间关联性检验。双相关g(r)函数主要以环代替K函数中的圆,计算过程没有累积效应[17],即:
(2)
当g(r)=1.0时,种群个体在r尺度上为随机分布,g(r)>1.0时为聚集分布,g(r)<1.0时为均匀分布。
双变量双相关函数g12(r)可以进行双变量点格局分析:
(3)
当g12(r) 值高于置信区间的上限,则2个种在r尺度为正关联;若低于置信区间的下限,则呈负关联;若在置信区间内,则表明2个种之间相互独立或没有关联性。用199次Monte Carlo随机模拟,产生置信度为99%的包迹线,以检验两个树种的分布格局和关联性是否显著[18]。
1.4 零模型检验
选择不同的零假设模型(Null model)进行点格局分析,单变量g(r)函数采用完全随机模型(Complete spatial randomness,CSR)和异质泊松模型(Heterogeneous poisson process,HP),双变量g12(r)函数采用完全空间随机模型和先决条件模型(Antecedent condition,AC),其中CSR模型实质是均质泊松过程,假设每个点在样地中的出现几率都是等同且相互独立,而HP模型排除了生境异质性的零模型。AC模型在生成双变量g12(r)函数的包迹线时,假设对照个体的位置不变,案例中的个体随机分布。数据分析及制图使用软件R 3.6.3完成。
2 结果与分析
2.1 径级结构
参照亚热带常绿阔叶林相关研究[19],对0.8 hm2样地内的4个主要树种的径级结构进行分析如图1,甜槠与木荷的径级结构均呈倒J型,43.4%的甜槠个体和53.6%的木荷个体DBH在0~4 cm之间,幼树数量丰富,种群更新良好,属增长型种群。而亮叶桦和蓝果树虽然在样地中空间优势度较高,但现存基本是大树和老龄树且个体较少,种群由成熟个体维持,缺乏幼苗储备,更新困难,从而导致种群趋于衰退。
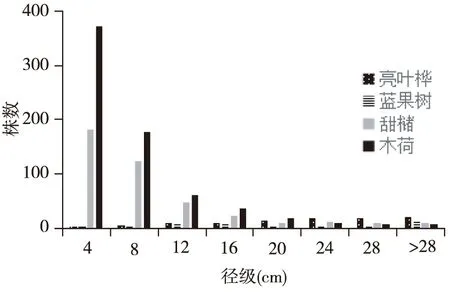
图1 主要空间优势树种的径级分布Fig. 1 Diameter at breast heightclass of 4 main tree species
2.2 优势树种的空间格局分析
通过g(r)函数对空间格局进行分析,结果如图2所示,首先采用完全随机模型分析样地4个空间优势树种的空间格局,在0~30 m的尺度上,甜槠和木荷分布趋势相似,均在<10 m尺度聚集性分布,随着尺度增大而呈聚集强度逐渐减弱,转为随机分布。亮叶桦在0~25 m尺度为聚集分布,后转为随机分布,蓝果树在0~30 m尺度表现为随机分布。采用异质泊松模型排除生境异质性后发现(图3)亮叶桦和蓝果树均表现为随机分布,甜槠在0~5 m、10~12 m,木荷在5~7 m为聚集分布,而甜槠在22~25 m,木荷在20~30 m呈弱的均匀分布趋势。
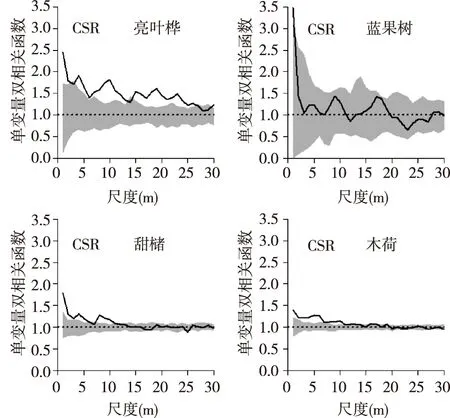
图2 完全随机模型下4个优势树种的空间格局Fig. 2 Spatial distribution pattern of 4 tree species under CSR null model
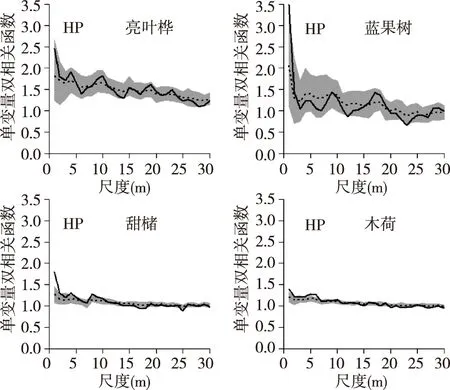
图3 异质泊松模型下4个优势树种的空间格局Fig. 3 Spatial distribution pattern of 4 tree species under HP null model
2.3 主要优势树种的空间关联性
4个优势树种的空间关联性情况如图4,在完全随机零模型下,亮叶桦与蓝果树种间的关联性为不相关,亮叶桦与甜槠在20~25 m正相关,亮叶桦和木荷在0~20 m尺度上表现为正相关,蓝果树和甜槠在8~10 m正相关,蓝果树与木荷0~5 m正相关,甜槠和木荷在0~10 m正相关,但除去亮叶桦与木荷正相关明显外,其他正相关性均不明显。而在先决条件零模型下,除亮叶桦与甜槠在20~25 m弱正相关外,其他树种两两之间的空间关联均为不相关。
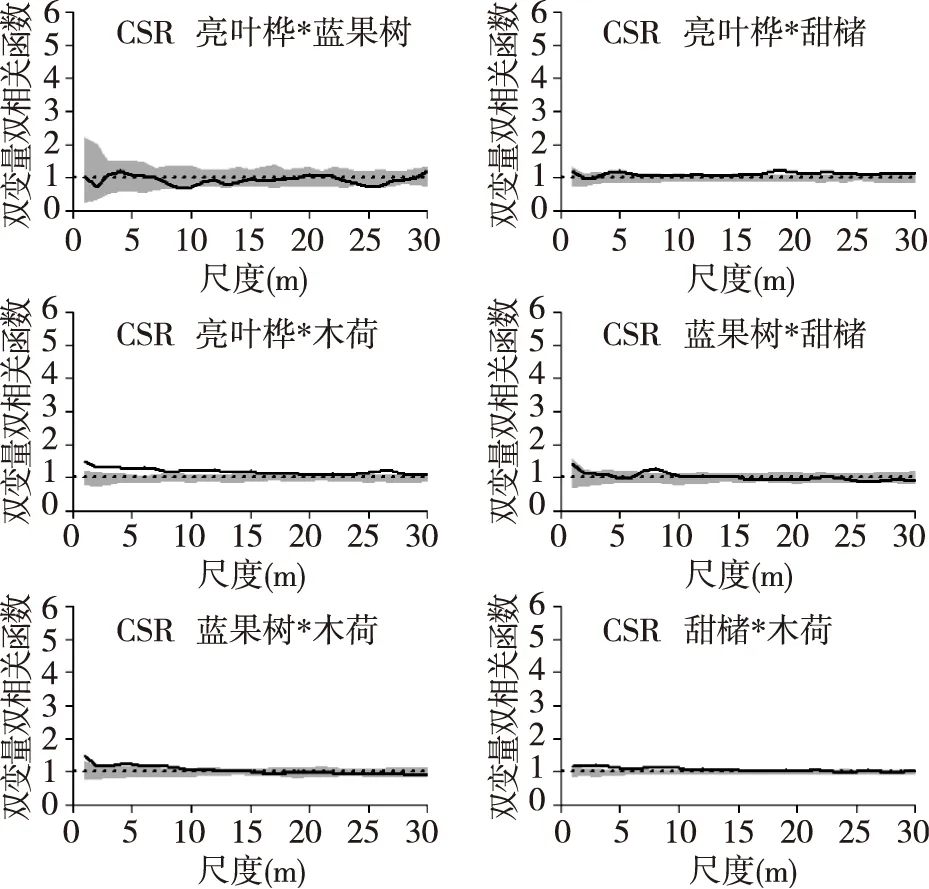
图4 完全随机模型下4个优势树种的空间关联Fig. 4 Spatial associations of 4 tree species under CSR null model
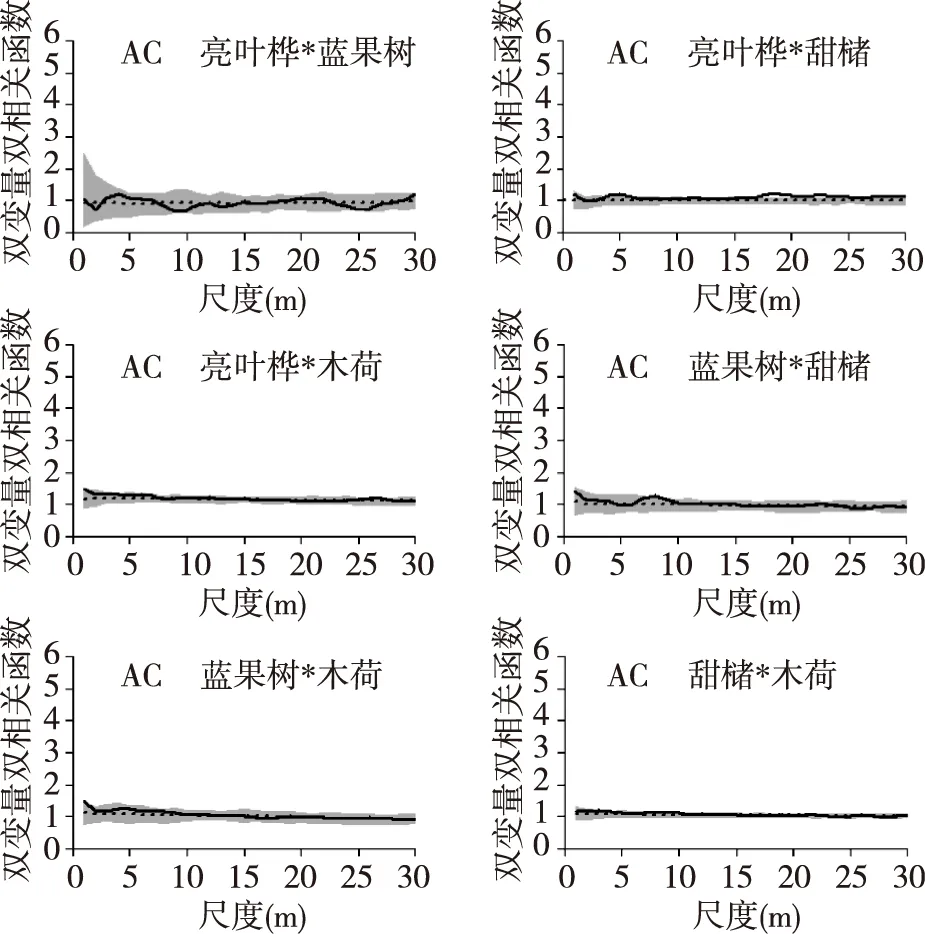
图5 先决条件模型下4个优势树种的空间关联Fig. 5 Spatial associations of 4 tree species under AC null model
3 讨 论
优势树种是森林群落结构形成和维持中的骨架和主导,对样地中优势种(成年个体,特别是大树)的分布格局及空间关联进行研究对于理解不同物种或功能类群之间的相互作用和群落的组成及潜在的生态学过程具有重要意义[20-21]。样地中亮叶桦和蓝果树属于阳性树种,是演替先锋种[22],其幼龄个体对林下光照条件要求高。由于保护区内天然次生林逐渐向老龄林演替,群落中现存的亮叶桦和蓝果树均以老龄树和大树为主,如蓝果树,样地中只有47株,但平均胸径为21.50 cm,亮叶桦100株,平均胸径20.86 cm。这种情况下导致群落中的亮叶桦和蓝果树逐渐被小径级个体充足,种群径级结构呈倒“J”形,自然更新良好,属增长型种群的优势种甜槠和木荷所替代。
在自然森林群落中,大部分物种种群呈聚集分布。其中小尺度上的同种聚集分布多因种子扩散限制及物种自身生物学特性作用导致[23]。本研究中在不排除生境异质性的影响时,样地中除蓝果树因个体较少在0~30 m尺度下为随机分布外,亮叶桦、甜槠和木荷的分布格局均呈小尺度聚集分布,随尺度增大转为随机分布,结果符合自然界中观察到的一般规律。其中亮叶桦个体少且多为大径级个体,却仍在0~25 m尺度呈现聚集分布,表明在天然林中同种个体聚集属普遍现象[24]。剔除生境异质性后,亮叶桦从聚集分布转为随机分布,而甜槠和木荷的总体来看,聚集性减弱,且在20 m尺度后出现了均匀分布趋势,这说明在常绿阔叶林样地中生境异质性显著影响树种的分布特征,但不同物种受影响的程度并不一样[25]。甜槠和木荷作为亚热带常绿阔叶林中常见的阳性先锋树种,在样地中分布更加广泛,有更多的幼苗储备。甜槠种子较大,可能受重力的作用而造成小树在母树周围聚集分布。而木荷种子有种翅,能传播较远,受重力限制作用较小,在排除生境异质性后,木荷在20~30 m尺度出现了趋于均匀分布的趋势。研究发现,随着群落由演替初期的低稳定性先锋群落演替为高稳定性的顶极群落,物种间的正相关性会逐渐加强[26]。本研究中4个空间优势树种,在完全随机零模型下,除亮叶桦与蓝果树种间的关联性为不相关外,其他3个树种均表现为在某一尺度上呈现出两两之间的正相关性,但大部分尺度总体表现为空间不相关特点,这说明主要树种间存在一定依赖性,种间竞争相对缓和。亮叶桦和木荷在0~20 m呈显著正相关,说明它们之间有相似的资源利用策略及生态重叠。而在先决条件零模型下,除亮叶桦与甜槠在20~25 m弱正相关外,其他树种两两之间的空间关联均为不相关。
4 结 论
4个主要树种的空间分布格局和种间的关联性随空间尺度的不同而变化,生境异质性显著影响树种的分布特征,但不同物种受影响的程度并不一样。群落由演替初期的低稳定性先锋群落演替为高稳定性的顶极群落,群落处于稳定发展阶段。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
防护林科技(2019年12期)2019-04-14
中成药(2017年3期)2017-05-17
中国环境监察(2016年12期)2016-10-24
花卉(2016年14期)2016-07-31
