
矮溲疏幼苗生理特性对淹水胁迫的响应
2021-08-16 07:33王叶滕尧张子雄陈彩霞龙秀琴
农业与技术 2021年15期
王叶滕尧张子雄陈彩霞龙秀琴
(1.贵州省山地资源研究所,贵州 贵阳 550001;2.贵州省植物园,贵州 贵阳 550004)
随着气候条件的不断变化,近些年一些地区夏季降雨量大且相对集中,易形成洼地积水,这对园林树种的选择提出了更高要求,开展园林植物淹水胁迫的研究十分必要。有学者通过测定植物生长量和营养水平来研究间断性淹水对棉花产量的影响[1]。曾建军等人对植物涝害产生机制以及植物应对水涝的相关机理研究进行了综述[2]。陈传红等人进行了淹水胁迫对西瓜幼苗生理活性物质SOD、POD和蛋白质、还原糖影响的研究[3]。仁情扎西等人从淹水所造成的危害以及植物对淹水的反应来探讨植物与水的生态关系,并从生理和形态结构方面分析了植物耐涝性的机理[4]。何嵩涛等人通过测定银杏叶片中超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等生理指标的测定来研究淹水条件下银杏膜脂过氧化作用及保护酶活性变化情况[5]。苏慧敏等人从形态和生理特征等方面对太阳扇扦插苗进行淹水研究[6]。
矮溲疏(Deatzia‘hybrida Boule’),虎耳草科(Saxifragaceae)溲疏属的双子叶植物,是溲疏的一个矮化变种,花白色或外面略带红晕且密集,是一种观赏性好、适应性强的观花小灌木,具有较好的园林绿化价值。从目前研究报道来看,对溲疏及矮溲疏的研究报道相对较少。苏婷等人通过对密花莲翘和矮溲疏新梢生长量、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)等生理指标的测定,从而进行了耐旱性差异比较[7];柴慈江等人对矮溲疏进行了试管苗的茎芽增殖和生根培养技术的研究[8];史宝胜等人对小花溲疏种子萌发条件进行了研究,并在萌发的时间、出现萌发高峰的时间和发芽率等方面均进行了差异性比较[9];邓洪平等人进行了多辐溲疏居群过氧化物酶同工酶分化研究[10];何平在光学显微镜和扫描电镜下对四川省溲疏属12种、3变种的花粉形态进行了初探[11]。但是尚未见淹水胁迫对矮溲疏生理特性影响的研究报道,因此,本项目进行了淹水胁迫对矮溲疏生理特性影响的研究,通过测定超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量、可溶性糖含量、根系活力等指标,了解其耐涝能力,为其在园林绿地中的运用提供理论依据。
1 材料与方法
1.1 试验材料
选择生长健壮的矮溲疏植株,剪取发育基本一致的新梢(保留3~4个节)进行扦插繁殖,扦插苗在珍珠岩中于小拱棚内越冬,春季萌动后,筛选生长良好的一年生矮溲疏扦插苗为试验材料,移入装有等量(约1600g)培养土(园土∶珍珠岩为4∶1)的塑料盆内,待新梢长到8cm左右进行淹水处理。
1.2 试验方法
采用盆栽方法,在水箱中进行淹水处理,水面高出培养土约1cm,以正常水分管理为对照。采用随机区组设计,每盆2株,每个小区2盆,3次重复,每处理共12株。每次取样6盆,其中3盆用于生理指标测定,3盆用于解除胁迫后的恢复性观察。设置8个处理,即淹水8d,淹水胁迫后每间隔24h取样测定。
1.3 试验指标
进行淹水处理后,每天早上8∶00进行取样,测定SOD活性、MDA含量、可溶性糖含量、根系活力等生理指标并进行植株解除胁迫后的恢复性观察。采用TTC法测定根系酶活力,蒽酮法测定植物叶片组织中可溶性糖含量,三氯乙酸(TCA)浸提的方法测定植物体丙二醛(MDA)含量,氮蓝四唑(NBT)还原比色法测定超氧化物歧化酶(SOD)活性[12]。
1.4 数据分析
试验数据采用Excel 2010软件进行统计分析,利用IBM SPSS Statistics 22.0软件做显著性分析(显著性阈值为p<0.05)。
2 结果与分析
2.1 淹水胁迫对矮溲疏外部形态的影响
淹水胁迫下矮溲疏外部形态变化和解除胁迫后的恢复性观察结果见表1。对照植株新梢生长快,叶片青绿,生长良好。淹水1~3d植株外观未表现明显的胁迫症状;第4天后,新梢生长量较对照慢,部分植株叶片出现萎焉、光泽度下降,但解除胁迫后在24h后可恢复生长状态;而淹水胁迫5d时,大部分叶片萎焉、下垂,解除胁迫后需约2d才可恢复;当淹水6d后,叶片表现黄色斑点,糜烂及脱落现象,解除胁迫后仍无法恢复。这说明淹水环境抑制了植物根系和叶片生理生化反应过程,从而表现相应的胁迫症状。

表1 淹水胁迫介导植株外部形态变化及解除胁迫时间
2.2 矮溲疏根系对淹水胁迫的响应
淹水胁迫下矮溲疏根系活力总体呈现先上升后逐渐下降的趋势。淹水处理第1天,根系活力迅速增高,极显著高于对照,而后逐渐降低,前4d均高于对照。第2天与第3天之间,第4天与第5天之间达到显著,而淹水前2d极显著的高于对照,见表2。由此说明矮溲疏在淹水处理后发生了明显的生理生化变化。
2.3 淹水胁迫下植物叶片丙二醛(MDA)含量变化
矮溲疏叶片MDA含量经过第1天的迅速增高后基本呈下降趋势。除第6天外,第1天与对照及其它各处理均达到显著,说明矮溲疏对淹水比较敏感,能在较短时间内合成较多的抗逆性物质,而后对淹水环境产生一定的适应性;第8天又显著低于第6天,这表明矮溲疏对淹水环境的适应性和忍耐力是有限的,经过6d的淹水后,植物体对淹水表现明显的胁迫响应。
2.4 淹水胁迫下叶片可溶性糖含量的变化
随淹水时间的推移,矮溲疏叶片中可溶性糖含量先上升后下降。淹水处理后,可溶性糖含量逐渐增高,第3天达到最大值,第4天显著下降,第5天低于对照。淹水3d的植物叶片中可溶性糖含量显著高于对照和其它处理,且极显著高于对照。由此说明,植物前3d对淹水环境产生忍耐力,植物通过大量分解多糖或降低光合中可溶性糖向多糖的转化速率来满足植物生长发育对营养物质的需求。
2.5 淹水胁迫下叶片超氧化物歧化酶(SOD)活性变化
矮溲疏幼苗淹水后,其叶片中超氧化物歧化酶(SOD)活性在第1天迅速增高,较对照显著,随后基本呈现上升趋势,第4天达到峰值后逐渐下降,而后在一定小范围内波动,但其活性仍高于对照。对SOD活性进行显著性测定,说明矮溲疏叶片中SOD活性对淹水环境敏感,且第4天极显著高于对照和其它试验处理,表明矮溲疏在淹水下可维持较高的SOD活性来增强耐涝能力。
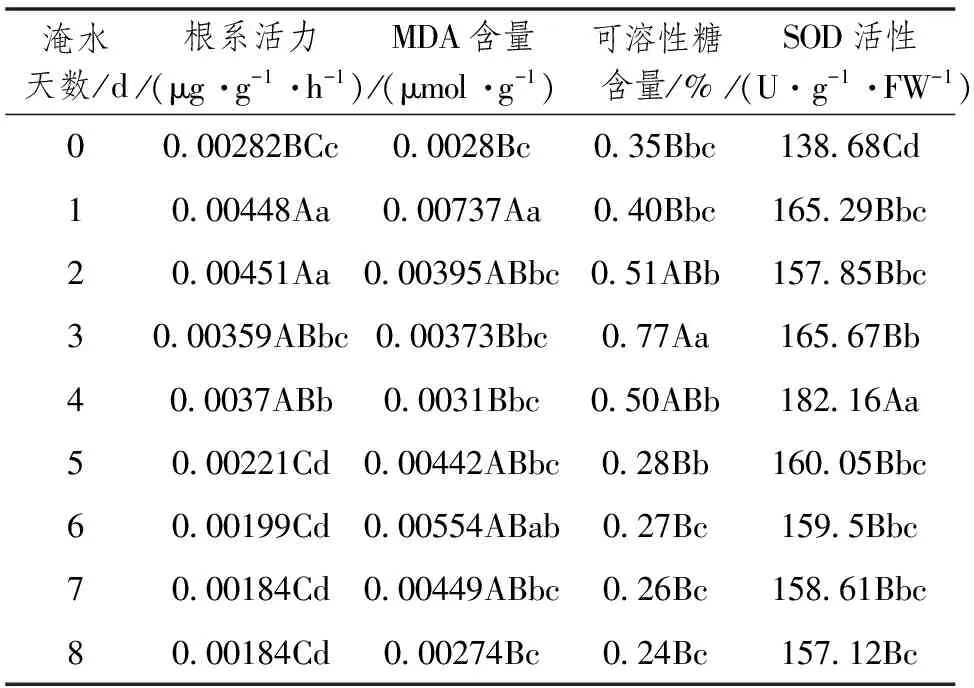
表2 矮溲疏幼苗在淹水胁迫下的生理生化变化
3 讨论与结论
淹水胁迫引起植物代谢紊乱、营养失调、乙烯增加、生长受抑[13]。其中活性氧易对蛋白质和核酸产生破坏作用,而在逆境胁迫下植物体内活性氧增加,使细胞膜中不饱和脂肪酸发生膜脂过氧化,细胞膜流动性降低,导致代谢紊乱。植物体内也有清除活性氧的酶系统,包括抗氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPX)、谷胱苷肽还原酶(GR)等[14]。矮溲疏在淹水胁迫下的超氧化物歧化酶(SOD)活性变化与杨宝铭等人对“寒富”苹果淹水胁迫下叶片和根系中SOD活性表现出先升高达到峰值,然后逐渐下降的类似趋势[15],之后持续保持一个较高的稳定水平,且远远高于对照,但与廖文燕等人对淹水胁迫下金钱松幼苗生理响应的研究结果相反[16],说明矮溲疏与金钱松可能对淹水环境有不同的耐受性。丙二醛(MDA)是植物细胞膜膜脂过氧化的产物之一,是植物受害的一个重要指标,王义强等人对淹水胁迫下银杏生理指标研究表明,随着淹水时间的推移,植物细胞膜脂过氧化增强,MDA含量急剧增加,细胞膜受害加重[17],其结果与本研究类似。SOD在植物体中将引起膜脂过氧化的超氧阴离子转化为氧化能力相对较弱的过氧化氢,而MDA是细胞膜膜脂过氧化产物。在本试验中,植物淹水第4天时,SOD活性达到峰值而MDA含量却降到最低,这与植物的生理特性相符。
可溶性糖和蛋白质含量变化是逆境胁迫中较为常见的生理现象,在胁迫条件下,为了维持体内与膨压相关的生理代谢得以正常进行,植物通过加快代谢活动来增加细胞质中溶质含量,降低渗透势维持细胞膨压[18]。淹水胁迫下矮溲疏叶片中可溶性糖含量呈现先急剧上升后下降的趋势,与陈传红等人对西瓜幼苗进行淹水胁迫的研究结果基本一致[3],而与苏慧敏等人对太阳扇扦插苗在淹水下的研究相反[6]。根系是植物水分吸收和养分摄取的主要器官,植物是否能从土壤中吸收水分取决于土壤和植物根系间的水势差,水势差的建立依赖于矿物离子的转运,而矿物离子向根内运输需要有氧呼吸释放能量[13],因此,淹水胁迫下抑制根系细胞的有氧呼吸,无氧呼吸增加,酒精等无氧代谢物积累,根系活力降低,本试验结果符合这一趋势,且在第5天显著低于对照,在“寒富”苹果的研究中也得出相同的结果[15]。
综合矮溲疏在淹水胁迫及解除胁迫后的形态变化和各生理生化指标认为,矮溲疏有较强的耐涝能力,能耐4~5d的淹水胁迫,可广泛运用于园林绿化。
猜你喜欢
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
Coco薇(2017年5期)2017-06-05
中国科技信息(2016年10期)2016-09-03
现代食品(2016年14期)2016-04-28
现代电生理学杂志(2015年4期)2015-07-18
中国粮油学报(2014年7期)2014-02-06
浙江大学学报(医学版)(2013年6期)2013-12-09
