
小鼠神经细胞中胱硫醚β合成酶催化生成的H2S对脑微血管内皮细胞增殖、迁移的影响及与VEGFR2的关系
2021-08-10 00:34陈佳妮陈志武
中国药理学通报 2021年8期
陈佳妮,陈志武
(安徽医科大学基础医学院药理教研室)
血管内皮生长因子(vascular endothelial growth factor,VEGF),是一种重要的血管生成调节因子,可增强内皮细胞有丝分裂、增殖和迁移及侧支血管的形成,在生理和病理性的血管生成中发挥着基本作用[1]。VEGF作用是通过其特异的受体血管内皮生长因子受体(vascular endothelial growth factor receptor,VEGFR)来发挥的。VEGFR分为VEGFR1、VEGFR2和VEGFR33个亚型,其中,VEGFR2主要参与内皮细胞有丝分裂的发生,介导了VEGF在内皮细胞中的生物学效应[2]。
缺血后神经损伤修复过程中,脑血管再生直接关系到神经组织损伤的发展和修复。在血管再生过程中,血管内皮细胞的迁移与增殖是十分关键的环节[3]。内源性硫化氢(hydrogen sulfide,H2S)是一种气态信号分子,在机体的生理和病理过程中发挥着重要的作用[4],主要由胱硫醚β合成酶(cystathionine-β-synthetase,CBS)和胱硫醚-γ-裂解酶(cystathionine-γ-lyase,CSE)催化L-半胱氨酸(L-cysteine,L-Cys)来生成。CBS和CSE有组织分布特异性,CBS分布在神经细胞中,而CSE分布在血管组织中[5]。因此,神经细胞主要通过CBS催化产生H2S。脑组织中的H2S参与了海马长时程增强、神经元保护等功能的调节,发挥了一种非典型神经递质的作用[6]。但内源性H2S也是一种重要的血管活性物质,参与血管的生理和病理过程。有研究表明H2S可促进人脐静脉内皮细胞的增殖和迁移[7],并有研究[8]显示H2S可激活VEGFR2,siRNA介导的VEGFR2基因敲低可抑制H2S诱导的人血管内皮细胞的迁移。但是由脑细胞产生的神经源性H2S是否对脑血管内皮细胞的迁移与增殖产生一定的影响呢?尚未见这方面的研究报道。因此,本文在体外观察了神经元CBS催化产生的H2S对小鼠脑微血管内皮细胞的增殖、迁移的影响,并初步探讨了其作用与VEGFR2的关系。
1 材料与方法
1.1 细胞小鼠海马神经元(HT22细胞株)、人脐静脉内皮细胞(HUVEC细胞株),均购自上海名劲生物科技有限公司;小鼠脑微血管细胞(bEnd.3细胞株),购自湖南丰晖生物科技有限公司。
1.2 主要试剂硫氢化钠NaHS购自美国Sigma公司;司马沙尼 (semaxinib,SU5416),购自美国MCE公司;重组小鼠血管内皮生长因子VEGFA/VEGF164购自美国Abbkine公司;胎牛血清FBS购自美国Gibco公司;高糖DMEM和RPMI 1640培养基购自美国Hyclone公司;CCK-8检测试剂盒购自美国Glpbio公司;4%多聚甲醛,0.1%结晶紫水溶液购自Biosharp公司;H2S含量检测试剂盒购自Solarbio公司;24孔Transwell小室(直径6.5 mm;孔径:8 μm;ref.3422)和6孔Transwell小室(直径24 mm;孔径:0.4 μm;ref.3450),购自美国Corning公司;L-Cys购自Macklin公司;氨基氧乙酸AOAA购自源叶生物公司;NaHS于使用前用生理盐溶液PBS溶解配制,并避光保存。
1.3 方法
1.3.1细胞培养 在37 ℃的5% CO2培养箱中,HUVEC细胞用含有10%FBS和1%青霉素-链霉素的RPMI 1640培养基培养,bEnd.3细胞和HT22细胞分别用含有10%FBS和1%青霉素-链霉素的高糖DMEM培养基培养,待细胞融合度达90%时进行传代培养,并分别加入测试药进行试验。
1.3.2HT22细胞和bEnd.3细胞共培养 在六孔板的Transwell小室的上室接种内皮细胞(bEnd.3细胞),下室接种神经细胞(HT22细胞)置入37 ℃的5% CO2培养箱中进行共培养。实验分为对照组、AOAA组(1 mmol·L-1)、SU5416组(10 μmol·L-1)、L-Cys组(100 μmol·L-1)、L-Cys+AOAA组、L-Cys+SU5416组,各组分别加入溶媒或相应的测试药物孵育24 h后,进行各种实验指标的检测。
1.3.3CCK-8增殖试剂盒检测增殖 在培养的HUVEC细胞或bEnd.3细胞培养孔中,分别加入不同浓度的NaHS、SU5416及阳性对照药VEGF164孵育一定时间后,弃除培养基,各孔细胞中分别加入100 μL新鲜培养基和10 μL的CCK-8测试液继续在37 ℃下孵育2 h后,用酶标仪读取各培养孔在450 nm处的吸光度值(A值),计算细胞增殖率:增殖率/%=(给药后实验组A值-空白组A值) / (给药前实验组A值-空白组A值)×100%。
1.3.4细胞划痕实验 将HUVEC细胞、bEnd.3细胞分别接种到24孔板,当细胞达到100%汇合时,使用无菌的200 μL移液器吸头在各孔单层细胞中心划一直线“伤口”,并用无菌PBS洗涤两次以去除死细胞。然后分别加入不同浓度的NaHS或SU5416或VEGF164以及500 μL含2%FBS的新鲜培养基(RPMI 1640和高糖DMEM)继续孵育24 h。在倒置显微镜下,拍照0、24 h细胞划痕伤口,并用ImageJ图像软件测定伤口面积。
1.3.5Transwell细胞迁移实验 在Transwell上室中接种100 μL无血清细胞悬液(bEnd.3细胞或HUVEC细胞),实验分为对照组、SU5416组(10 μmol·L-1)、NaHS组(10-3.5mol·L-1)、NaHS+SU5416处理组、VEGF164组(10 ng·mL-1)、VEGF164+SU5416处理组,下室加入含10% FBS的RPMI 1640和高糖DMEM培养基各500 μL;共培养组在上室接种bEnd.3细胞(100 μL无血清细胞悬液),下室接种HT22细胞。在37 ℃下孵育24 h后,将上室膜在室温下用4% 多聚甲醛固定30 min,使用棉签除去残留在上室膜上表面的未迁移细胞,并用0.1 % 结晶紫染色在室温下放置15 min。拍照,用Image-Pro Plus软件计数。
1.3.6内源性H2S含量的甲基蓝法检测 将神经细胞和内皮细胞的共培养基取出250 μL置于离心管中,用250 μL 1% 醋酸锌和10% 三氯乙酸处理,然后与20 μmol·L-1N,N-二甲基-对苯二胺硫酸盐和30 μmol·L-1氯化铁在1.2 mol·L-1盐酸中混合。在室温下孵育25 min后,用酶标仪在665 nm处检测吸光度,并使用空白管调零。根据NaHS标准曲线计算H2S含量。
1.3.7细胞内游离Ca2+浓度的钙荧光成像法检测 如前所述在神经细胞和内皮细胞共培养中,在Transwell小室的上室中放置一直径为14 mm的圆形玻璃盖玻片,将bEnd.3细胞接种在该盖玻片上。在37 ℃的5% CO2培养箱中培养一定时间后,在细胞中加入4 μmol·L-1Fluo-8 / AM和0.02% Pluronic F-127,继续孵育30 min。用正常生理溶液 (NPSS:145 mmol·L-1NaCl,3 mmol·L-1KCl,1 mmol·L-1CaCl2,2 mmol·L-1MgCl2,10 mmol·L-1glucose,10 mmol·L-1HEPES,pH 7.4) 洗涤后,将含有细胞的盖玻片安装到Ca2+成像系统记录室中记录荧光强度(Fluorescence intensity,FT),用波长Ex/Em=490/520 nm来进行检测,以FT来反映细胞内游离钙浓度的变化。
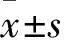
2 结果
2.1 NaHS促进小鼠脑微血管和人脐静脉内皮细胞的增殖Fig 1A所示,与对照组比较,加入H2S 供体NaHS(1×10-8—1×10-3.5mol·L-1)培养24 h时,小鼠脑微血管内皮细胞bEnd.3细胞和人脐静脉内皮细胞 HUVEC细胞增殖有明显的增强(P<0.01)。VEGF164(10 μg·L-1)有类似的增强作用(P<0.01)。结果表明外源性H2S对小鼠脑微血管和人脐静脉内皮细胞的增殖有明显的促进作用。Fig 1B与对照组相比,加入1×10-3.5mol·L-1NaHS 1 h时,bEnd.3细胞和HUVEC细胞的增殖作用不明显,但加入1×10-3.5mol·L-1NaHS 6 h时,bEnd.3细胞和HUVEC细胞增殖作用开始增强,而加入48 h时,NaHS增殖作用进一步增加。

Fig 1 Effects of NaHS on proliferation ability of bEnd.3 and HUVEC
2.2 SU5416对NaHS诱导的小鼠脑微血管和人脐静脉内皮细胞的增殖的影响如Fig 2所示,在bEnd.3细胞和HUVEC细胞中分别加入VEGFR2阻断剂SU5416 10 μmol·L-1培养24 h对这两种细胞的增殖均无明显的影响,但可明显地抑制1×10-3.5mol·L-1NaHS引起的bEnd.3细胞和HUVEC细胞的增殖,提示外源性H2S促进小鼠脑微血管和人脐静脉内皮细胞增殖作用可能与VEGFR2有关。

Fig 2 Effects of SU5416 on NaHS-promoted proliferation of bEnd.3 and HUVEC
2.3 NaHS促进小鼠脑微血管和人脐静脉内皮细胞迁移及SU5416对其的影响Fig 3结果中,与对照组相比, NaHS(1×10-6—1×10-3.5mol·L-1)加入24 h可明显并呈浓度依赖性地增加bEnd.3细胞和HUVEC细胞的迁移面积(P<0.01);Fig 4的Transwell实验结果表明与溶媒对照组相比,加入NaHS 1×10-3.5mol·L-124 h可明显地增加bEnd.3细胞和HUVEC细胞从Transwell上室膜内迁移到膜外的细胞数(P<0.01)。10 μg·L-1VEGF164对bEnd.3细胞和HUVEC细胞的迁移面积和迁移细胞数有类似的作用(P<0.01)。结果表明外源性H2S可明显地促进小鼠脑微血管及人脐静脉内皮细胞的迁移。Fig 4还表明SU5416单独加入24 h对bEnd.3细胞和HUVEC细胞的迁移细胞数无明显的影响,但可明显地减少1×10-3.5mol·L-1NaHS或10 μg·L-1VEGF164引起的bEnd.3细胞和HUVEC细胞的迁移细胞数的增加,提示外源性H2S促进小鼠脑微血管及人脐静脉内皮细胞的迁移可能与VEGFR2有关。

Fig 3 Influence of NaHS on migration of bEnd.3 and HUVEC cells(×100)
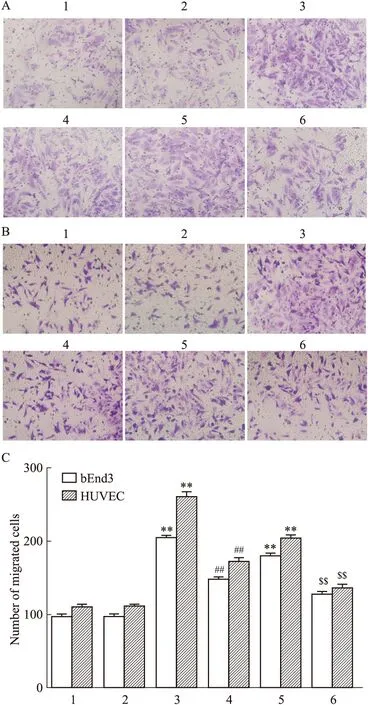
Fig 4 NaHS-promoted migration of bEnd.3 and HUVEC cells and effects of SU5416 on
2.4 神经元CBS催化产生的H2S促进小鼠脑微血管内皮细胞的增殖和迁移结果如Fig 5和Fig 6所示,在小鼠海马神经元HT22细胞与脑微血管bEnd.3内皮细胞的共培养体系中,H2S合酶CBS底物L-cys(100 μmol·L-1)可明显地增加bEnd.3细胞增殖和迁移细胞数(P<0.01);CBS抑制剂AOAA(1 mmol·L-1)单独使用对bEnd.3细胞增殖和迁移细胞数无明显的影响,但可明显地减弱L-cys引起的bEnd.3细胞增殖和迁移细胞数的增加。结果提示神经元CBS催化生成H2S可促进小鼠脑微血管内皮细胞的增殖和迁移。
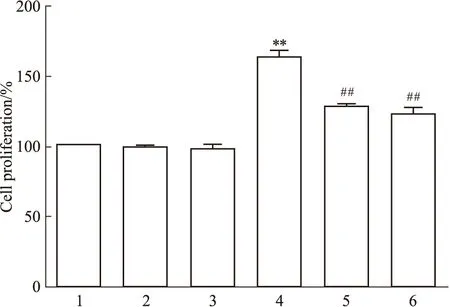
Fig 5 Effect of neuro CBS-derived H2S on proliferation of bEnd.3cells in Transwell co-culture
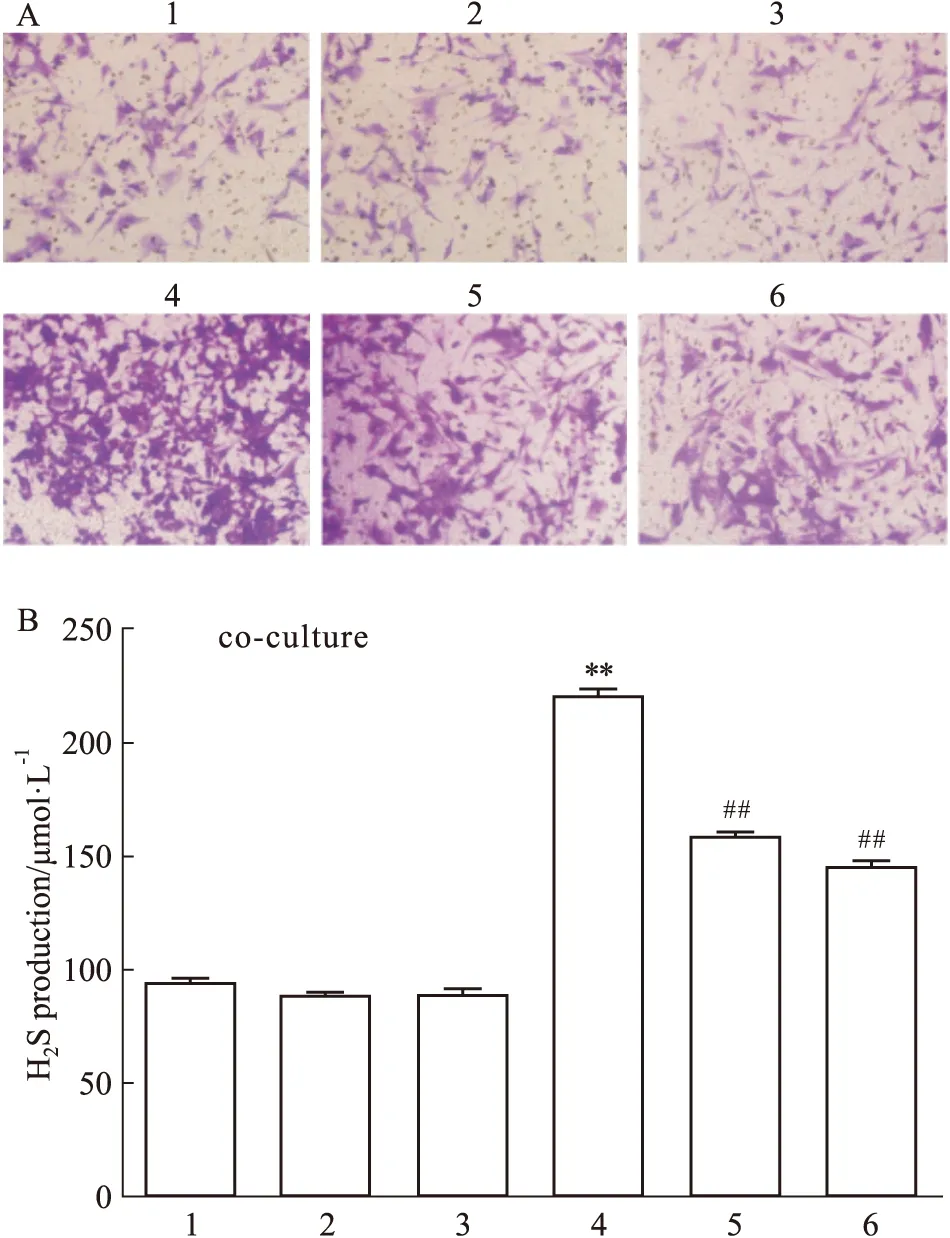
Fig 6 Effect of neuro CBS-derived H2S on migration of bEnd.3 cells in Transwell co-culture system(×200)
2.5 阻断VEGFR2对神经元CBS催化产生的H2S促小鼠脑微血管内皮细胞的增殖及迁移的影响如Fig 5和Fig 6所示,VEGFR2阻断剂SU5416单用对bEnd.3细胞增殖和迁移无明显的影响,但可明显地减弱L-Cys促进bEnd.3细胞增殖和迁移作用,提示HT22神经元CBS催化产生的H2S促小鼠脑微血管bEnd.3内皮细胞的增殖和迁移作用可能与VEGFR2有关。
2.6 L-Cys单用及合用AOAA或SU5416对小鼠海马神经元和内皮细胞共培养中H2S生成的影响
结果如Fig 7所示,CBS底物L-Cys可明显提高小鼠海马神经元HT22细胞与脑微血管内皮细胞bEnd.3细胞共培养中的H2S含量(P<0.01);CBS抑制剂AOAA和VEGFR2阻断剂SU5416单用对共培养中的H2S含量均无明显影响,但AOAA可明显地降低L-Cys引起的H2S含量的提高,而SU5416却无明显的影响。结果提示,L-Cys可通过海马神经元CBS催化生成H2S来提高共培养中的H2S含量。

Fig 7 Effect of L-Cys alone or in combination with AOAA or SU5416 on H2S content in co-culture system of bEnd.3 cells and HT22
2.7 L-Cys单用及合用AOAA或SU5416对共培养中小鼠脑微血管内皮细胞的胞内Ca2+荧光强度的影响结果如Fig 8所示,在小鼠海马神经元HT22细胞与脑微血管内皮细胞bEnd.3细胞共培养中,加入CBS底物L-Cys可明显提高内皮细胞内Ca2+荧光强度(P<0.01);加入CBS抑制剂AOAA和VEGFR2阻断剂SU5416单用对内皮细胞内Ca2+荧光强度均无明显影响,但AOAA 和SU5416能够降低L-Cys引起的内皮细胞内Ca2+荧光强度的提高。结果提示L-Cys可通过小鼠海马神经元CBS催化生成H2S来提高共培养中的内皮细胞内游离Ca2+浓度,并可能与VEGFR2有关。
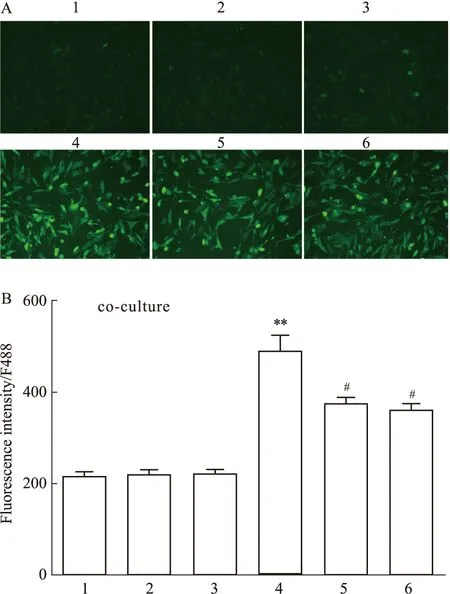
Fig 8 Effect of L-Cys alone or in combination with AOAA or SU5416 on Ca2+ fluorescence intensity in bEnd.3 cells co-cultured with HT22
3 讨论
缺血性脑损伤既有神经组织的损伤,也有脑血管的损伤,并且脑血管损伤或堵塞导致的脑血流减少甚至停止,是神经组织损伤的最直接原因[9]。因此,缺血后脑损伤修复中,恢复脑血流供应是至关重要的。缺血后脑血流供应的恢复不仅可通过脑血管的舒张,新的脑血管生成对恢复缺血区的血流供应和促进神经细胞损伤的修复也是非常重要的。
血管生成是指从已经存在的毛细血管或其后静脉形成新的血管,包括血管内皮基质降解、内皮细胞移行与增殖、内皮细胞管道化形成血管环等极其复杂的过程[10]。其中,血管内皮细胞及其增殖与迁移能力在血管生成中是至关重要的[11]。本研究在人脐静脉内皮细胞的HUVEC细胞上,观察到了H2S供体NaHS促进细胞增殖和迁移作用,这与Longchamp等[12]报道的H2S可促进人脐静脉内皮细胞增殖和迁移是一致的。本研究进一步在小鼠脑微血管内皮细胞的bEnd.3细胞中,首次发现NaHS可明显地增加bEnd.3细胞的增殖及迁移面积与迁移细胞数,表明H2S可促进小鼠脑微血管内皮细胞的增殖和迁移。
血管内皮细胞迁移和增殖受到多种细胞信号调控[13]。其中,VEGF/VEGFR2信号最为重要[14]。H2S可促进小鼠脑微血管内皮细胞的增殖和迁移可能涉及有VEGFR2机制。有研究[15]表明,siRNA介导的VEGFR2基因敲低可抑制H2S诱导的人血管内皮细胞的迁移。本研究发现,VEGFR2阻断剂SU5416不仅可以降低H2S对人脐静脉内皮细胞的增殖和迁移的促进,也可明显地抑制H2S促进小鼠脑微血管内皮细胞的增殖和迁移。结合文献研究报道H2S可激活VEGFR2[8],我们的结果提示外源性H2S可能通过激活VEGFR2来促进小鼠脑微血管及人脐静脉内皮细胞的增殖和迁移。
神经细胞中H2S合酶CBS催化产生的H2S可参与神经系统的生理和病理过程的调节。本研究将小鼠海马神经元(HT22细胞)与小鼠脑微血管内皮(bEnd.3细胞)进行共培养,发现H2S合酶CBS底物L-cys可明显地增加bEnd.3细胞增殖、迁移及共培养中的H2S含量,而CBS抑制剂AOAA虽然单独使用对bEnd.3细胞增殖、迁移无明显影响,但可明显地减弱L-Cys引起的bEnd.3细胞增殖、迁移并减少H2S的含量。由于CBS是脑细胞特有的H2S合酶,结果表明小鼠海马神经元CBS可催化L-Cys生成H2S来促进小鼠脑微血管内皮细胞的增殖和迁移,也即小鼠海马神经元CBS催化产生的H2S可促进脑微血管内皮细胞的增殖和迁移。
本研究在HT22细胞与bEnd.3细胞共培养中,进一步发现虽然VEGFR2阻断剂SU5416本身对bEnd.3细胞增殖和迁移无明显的影响,但可明显地减弱L-Cys经CBS催化生成H2S引起的bEnd.3细胞增殖和迁移的增加,并且此时共培养中的H2S含量无明显的变化,结果表明神经元CBS催化生成的H2S增加脑微血管内皮细胞的增殖和迁移作用可能与VEGFR2可能有关。结合上述的外源性H2S可通过激活VEGFR2来促进bEnd.3细胞的细胞增殖和迁移,结果提示神经元CBS催化生成的H2S可激活VEGFR2促进脑血管内皮细胞增殖和迁移,至于H2S如何激活VEGFR2及其详细机制有待于今后的进一步研究。
Ca2+是参与调节细胞功能的一种重要信使离子,可参与基因转录、细胞分泌、细胞增殖及迁移等多种生理功能的调节。有研究[15]表明,抑制钙库操纵性钙内流能够明显抑制内皮细胞的增殖、迁移。本研究在小鼠海马神经元与脑血管内皮细胞共培养模型中,发现CBS底物L-Cys可明显提高内皮细胞内Ca2+荧光强度,并可被CBS抑制剂AOAA明显减弱,表明L-Cys可通过神经元CBS催化生成H2S来提高内皮细胞的胞内游离Ca2+浓度。并且进一步观察到L-Cys引起的提高内皮细胞内Ca2+荧光强度的提高可被VEGFR2阻断剂明显减弱,结合上述的VEGFR2在外源性H2S或神经元CBS催化生成的H2S在内皮细胞增殖和迁移中的作用,提示神经元CBS催化生成的H2S可能通过激活VEGFR2,导致脑血管内皮细胞内游离Ca2+浓度升高来促进细胞增殖和迁移。
综上所述,神经元CBS催化生成的H2S可促进脑血管内皮细胞增殖和迁移,其可能是与激活VEGFR2,升高内皮细胞的胞内游离Ca2+浓度有关。
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
昆明医科大学学报(2021年3期)2021-07-22
世界最新医学信息文摘(2021年12期)2021-06-09
宁夏医学杂志(2020年3期)2021-01-21
创新作文(小学版)(2019年10期)2019-09-25
天然产物研究与开发(2018年4期)2018-05-07
小学生学习指导(低年级)(2017年5期)2017-05-04
中国医药导报(2015年26期)2015-02-28
郑州大学学报(医学版)(2015年2期)2015-02-27
中国医疗美容(2015年5期)2015-02-03
