
3种治理模式对石漠化地区土壤微生物群落结构的影响及治理能力比较
2021-08-02 15:52黄磊曾亚军郭金鹏许才万
江苏农业科学 2021年12期
黄磊 曾亚军 郭金鹏 许才万

摘要:为探寻较为合理且高效的石漠化植被修复的治理模式,为今后选择治理石漠化的植被组合提供参考依据。在贵州省黔东南苗族侗族自治州施秉县设置样地,选择3种治理模式:模式一为针叶林(马尾松、柏木),模式二为阔叶林(壳斗类,杂灌),模式三为灌木林(荚蒾、杂灌),分析不同治理模式下的土壤养分状况、土壤微生物群落结构、土壤微生物与环境因子相关性。结果表明,3种模式下土壤中微生物数量为细菌>真菌>放线菌;模式二治理下的土壤呈弱碱性,土壤养分含量高于其他2种模式,土壤微生物具有更高的多样性,罗尔斯通氏菌菌属(H16)、Gaiella、红游动菌属(Rhodoplanes)、芽单胞菌属(Gemmatimonas)、Haliangium、子囊菌门(Ascomycoat)、担子菌门(Basidiomycete)、接合菌门(Zygomycota)是其优势菌属,且与土壤中大多数营养物质具有显著正相关性;石漠化治理效果表现为模式二>模式一>模式三,所以模式二的石漠化治理能力更好。
关键词:石漠化;植被修复;治理模式;土壤养分;土壤微生物群落结构;土壤微生物多样性
中图分类号:S154.3 文献标志码: A
文章编号:1002-1302(2021)12-0201-08
收稿日期:2020-08-25
基金项目:贵州黎平石漠化生态系统国家定位观测研究站运行补助专项经费。
作者简介:黄 磊(1987—),男,四川凉山人,硕士,主要从事森林培育研究。E-mail:465125951@qq.com。
通信作者:曾亚军,硕士,副研究员,主要从事森林培育理论与技术方面的研究。E-mail:yjz410@163.com。
石漠化也称为石质荒漠化,指由于水土流失造成的地表植被减少和土壤损失、基岩裸露,最终导致土地丧失农业利用价值和生态环境退化的现象[1]。我国西南地区石漠化面积达13 000 km2,是世界上最大的喀斯特地貌分布区之一[2-3]。石漠化已成为严重阻碍我国西南地区经济增长的主要生态灾害[4],石漠化地区的生态环境更加脆弱,人均可耕地减少;由于在该条件下植被生长困难,从而导致土壤储水功能降低,造成人畜饮水困难等问题[5]。全国范围内石漠化最为严重的是贵州省,其次是云南和广西地区,资料显示,目前贵州省还有396.33万hm2土地适宜进行生态修复,任务十分艰巨[6]。贵州省石漠化严重阻碍了当地生态环境及可持续经济发展。因此,石漠化治理在贵州地区一直都是政府关注和环境科研工作者研究的重点。
1990年以来,石漠化地区的生态退化环境一直靠植被修复的方式来治理[7]。目前,关于石漠化生态环境修复的研究主要集中在不同治理模式下植物群落组成及多样性、造林保存率、土壤肥力、土壤酶活力等方面[8-10]。近年来,研究发现土壤微生物对改善土壤质量具有重要作用[11],土壤微生物多样性对塑造陆地生态系统的功能具有重要作用[12]。一方面,可以通过植被固定土壤来改善石漠化程度,另一方面,该过程中土壤营养物质不断积累,能促进植被生长。此外,土壤微生物对维持土壤活力具有重要意义[13],作为土壤中物质转化的动力,土壤微生物在土壤的形成及改变土壤肥力等方面有着不可替代的作用[14]。土壤微生物对土壤环境变化十分敏感,研究显示在石漠化植被恢复过程中细菌是土壤微生物中的优势类群[15-17],因此土壤中细菌多样性与石漠化生态系统恢复过程中土壤营养物质变化密切相关,可作为评价土壤质量变化即石漠化修复效率的指标[18]。
目前关于石漠化治理的研究主要集中在探讨一种治理模式或该治理模式下植被演替过程中土壤中微生物群落结构变化,而在不同的治理模式下土壤中的微生物群落结构及治理效果比较的研究则鲜见报道。因此,本研究拟从3种不同的治理模式入手,分别为模式一:针叶林,马尾松、柏木;模式二:阔叶林,壳斗类、杂灌;模式三:灌木林,荚蒾、杂灌。以上3种模式中的马尾松[19-20]、柏木[21]、壳斗类植物[22-23]、荚蒾[24]、杂灌植物[25]都被研究报道过是石漠化治理的先锋树种。因此,探究以上3种不同治理模式下土壤养分含量及土壤微生物群落结构的异同,旨在找到较为合理且高效的石漠化植被修复的治理模式,为今后石漠化治理工作提供一定的选择依据,提高石漠化治理效果从而达到保护生态环境的目的。
1 材料与方法
1.1 研究地区概况
研究地区位于贵州省黔东南苗族侗族自治州施秉县(108° 06′00″~108° 07′12″ E、27°12′00″~27°10′12″ N),屬亚热带湿润季风气候,年均温度 16 ℃,年均降水 1 220 mm[26]。其喀斯特石漠化面积282.95 km2,是典型的石漠化代表地区[27]。采样地点如下:编号2 (108°14′38″ 3E、26°57′31″ 3N)即治理模式一,海拔850 m,其植被类型为针叶林,主要树种为马尾松、柏木;编号12(108°14′26″ E、26°57′57″ N)即治理模式二,海拔833 m,其植被类型为阔叶林,主要树种为壳斗类、杂灌;编号26(108°15′32″ E 、26°57′46″ N)即治理模式三,海拔887 m,其植被类型为灌木林,主要树种为荚蒾、杂灌。
1.2 采样
采样时间为2018年秋季,每样地设置3个采样点,根据实际情况尽量挖深度为0~20 cm的土壤剖面,每个采样点取2份土壤样品,各500 g,1份用于测定土壤养分含量,1份用于微生物群落结构分析。
1.3 土壤养分含量测定
测定土壤pH值及有机碳、硝态氮、铵态氮、全钾、有机质、全氮、速效钾、有效磷含量。
1.4 DNA提取及高通量测序
宏基因组DNA提取,并测序分析。
1.5 数据分析
利用QIIME (V1.9.1)、Canoco 4.5、MEGA 5.0、Mothur、SPSS 25.0等软件对数据进行分析整理。
2 结果与分析
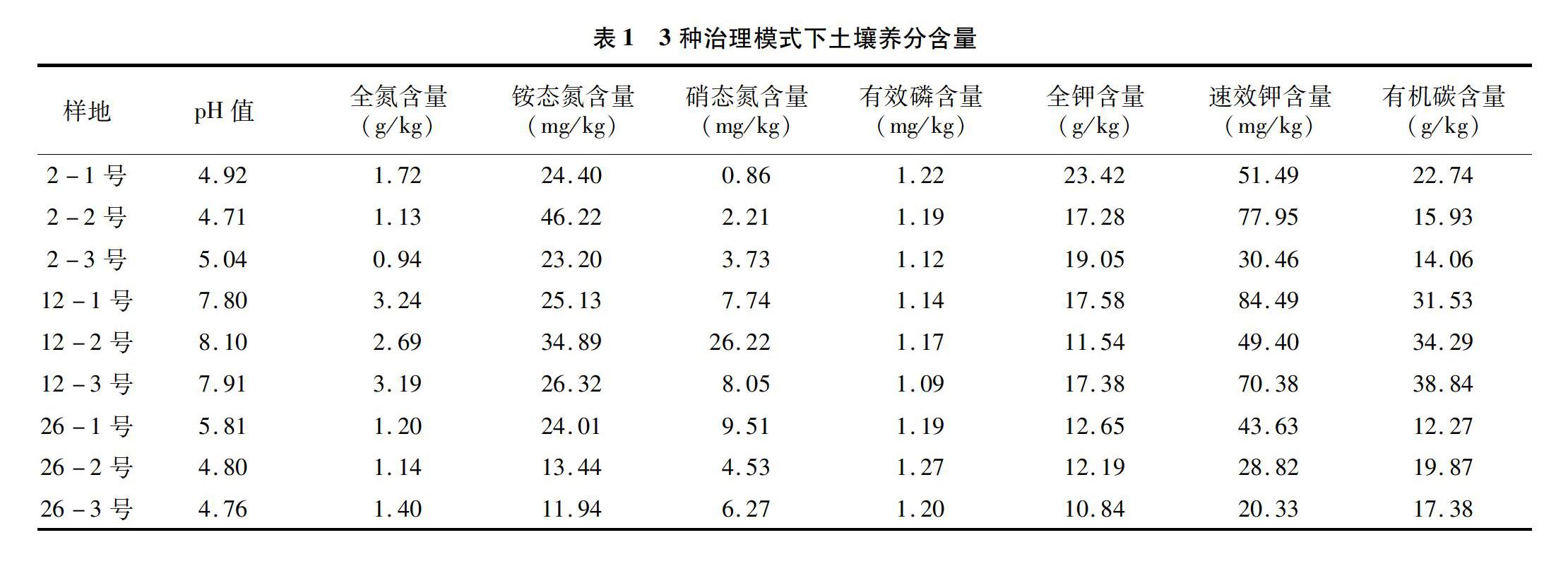
2.1 3种治理模式下各土壤养分含量
由表1可以看出,除了在模式二治理下土壤呈弱碱性外其余2种治理模式下土壤都呈酸性,这表明不同治理模式会对土壤的酸碱性产生不同的影响。在模式二治理下,土壤的全氮、铵态氮、硝态氮及有机碳含量都较其他2种治理模式高。有效磷含量在3种治理模式下并无明显差异。而全钾含量在模式一治理下的土壤中含量最高,速效钾在模式二下含量最高。
土壤有机质、氮、磷、钾等养分含量与石漠化程度密切相关,综上分析结果表明整体上在第2种治理模式下土壤具有更高的养分,其次是模式一、模式三。所以石漠化治理效果为模式二>模式一>模式三。
2.2 3种治理模式下土壤细菌群落结构
不同治理模式下土壤的营养成分即恢复效果不同,可能是由于在不同的植被治理下其土壤微生物群落结构组成差异造成的。土壤微生物是土壤的重要组成部分,在土壤中动植物残体和有机质及其有害物质的分解、生物化学循环和土壤结构的形成过程起着重要的作用。因此,本文对所采用的3种治理模式下的土壤微生物群落结构进行研究,目的在于通过解析土壤微生物群落结构为不同石漠化治理模式恢复效率,为更好地选择石漠化治理模式提供理论依据。
在石漠化治理模式植被恢复过程中开展土壤微生物区系的数量和组成研究,有助于诊断石漠化地区土壤质量和植物营养的修复状况,土壤微生物菌落统计结果显示细菌>真菌>放线菌数量。
2.2.1 土壤细菌群落结构
由物种相对丰度(图1)可以看出,在3种治理模式下的土壤微生物中优势菌群都是变形菌门(Proteobacteria,46.256%、35.727%、31.570%)、酸杆菌门(Acidobacteria,22.145%、22.988%、32.584%)以及疣微菌门(Verrucomicrobia,8.353%、14.071%、32.584%)。根据所有样品在属水平的物种注释及丰度信息,选取丰度排名前35的属,根据其在每个样品中的丰度信息进行聚类,结果显示(图2)在3种治理模式下其微生物的群落结构明显不同。在模式一植被治理下,根瘤菌属(Rhizobium,0.749%)、溶杆菌属(Lysobacter,1.344%)、马赛菌属(Massilia,0.456%)、土微菌属(Pedomicrobium,0.562%)、寡养单胞菌属(Stenotrophobacter,0.261%)以及分支杆菌属(Mycobacterium,0.367%)等是主要优势菌属。在模式二治理下,罗尔斯通氏菌菌属(H16,3.557%)、Gaiella(0.382%)、红游动菌属(Rhodoplanes,3.613%)、芽单胞菌属(Gemmatimonas,0.612%)及Haliangium(0.982%)是优势菌属。而在模式三治理土壤中根微菌属(Rhizomicrobium,0.936%)、苔藓杆菌属(Bryobacter,0.553%)及酸杆菌属(Acidibacter,0.925%)等属于优势菌属。同时,由于土壤理化性质结果显示模式二治理下土壤肥力较高,因此可推测在模式二的植被治理下其土壤中的优势微生物比其他微生物在石漠化治理过程中具有更重要的作用,为今后选择治理石漠化植被提供参考依据。
2.2.2 土壤细菌多样性
总体上来看,各个样品的覆盖率均大于98%,说明样品的采样率足够获得最丰富的OTU(表2)。选择香农指数(Shannon)、辛普森多样性指数(Simpson)和Chao1指数,对不同样品在97%一致性阈值下的α多样性进行分析。从表2可以看出,3种治理模式下土壤微生物的香农指数分别为9.20、9.29、9.33,辛普森多样性指数均为0.99,Chao1指數分别为3 707.239、3 988.579、4 128.411。表明这几种土壤样品的群落结构差异不显著,具有较高的微生物多样性。
选用Weighted Unifrac距离和Unweighted Unifrac距离这2个指标来衡量样品之间的相异系数。由图3可知,以Weighted Unifrac距离作为标准(上方),内生细菌群落相异系数最大的为模式一与模式三,为0.226。结果表明模式二与模式一、模式三之间内生菌的物种多样性差异低于模式一与模式三之间的内生菌多样性。以Unweighted Unifrac距离作为标准(下方),相异系数最大的为模式一与模式三,为0.303;其次为模式一与模式二,为0.287。结果表明3种治理模式下土壤微生物群落结构具有差异。
2.2.3 3种治理模式下土壤细菌组成与环境因子相关性
微生物菌属与环境因子相关性(图4)结果表明,全钾含量与模式一中优势菌属根瘤菌属(Rhizobium,0.828,P<0.01)、寡养单胞菌属(Stenotrophobacter,0.867,P<0.01)以及分支杆菌属(Mycobacterium,0.833,P<0.01)等菌属具有极显著正相关关系。有效磷含量仅与模式三中优势菌属酸杆菌属(Acidibacter,0.921,P<0.01)呈现极显著正相关关系。
其余的环境因子与模式二中优势菌属具有显著正相关性。如有机碳含量与Gaiella(0.683,P<0.05)显著正相关;速效钾与Gaiella(0.767,P<0.05)、红游动菌属(Rhodoplanes,0.739,P<0.05)具有显著正相关关系;硝态氮与芽单胞菌属(Gemmatimonas,0.867,P<0.01)及Haliangium(0.850,P<0.01)表现出极显著正相关关系;铵态氮与Gaiella(0.883,P<0.01)、红游动菌属(Rhodoplanes,0.840,P<0.01)具有极显著正相关性;全氮含量与芽单胞菌属(Gemmatimonas,0.700,P<0.05)具有显著正相关性;最后是pH值与Gaiella(0.683,P<0.05)、Haliangium(0.700,P<0.05)具有显著正相关性。由此可以看出,土壤中的大多数营养物质与模式二治理下的土壤微生物具有正相关性,意味着在模式二的治理下土壤具有更高的营养,有利于植被生长,石漠化治理效果最好。
2.3 3种治理模式下土壤真菌群落结构
2.3.1 土壤真菌群落结构
由图5可知,在3种治理模式下真菌群落结构各有不同。在模式一治理下子囊菌门(Ascomycoat,71.263%)>担子菌门(Basidiomycete,14.972%)>接合菌门(Zygomycota,10.331%),为主要优势菌群。模式二治理下子囊菌门(Ascomycoat,56.731%)>担子菌门(Basidiomycete,38.729%)>接合菌门(Zygomycota,3.702%)。而在模式三治理下担子菌门(Basidiomycete,95.005%)为优势菌群。根据所有样品在属水平的物种注释及丰度信息,选取丰度排名前35的属,根据其在每个样品中的丰度信息进行聚类。结果(图6)显示在3种治理模式下其微生物的群落结构明显不同。在模式一治理下木霉属(Trichoderma,6.107%)、Ilyonectria(4.351%)、Idriella(2.859%)、青霉菌属(Penicillium,1.256%)、枝顶孢霉属(Acremonium,0.199%)、镰刀菌属 (Fusarium,0.584%)、Cyphellophora(0.095%)等为优势菌属。模式二治理下短梗蠕孢属(Trichocladium,3.637%)、块菌属(Tuber,1.906%)、大团囊菌属(Elaphomyces,2.074%)、Gloeoporus(0.150%)、Campylocarpon(0.139%)、蘑菇属(Clavaria,0.358%)、明梭孢属(Monographella,0.103%)、多毛菌属(Hirsutella,0.850%)、鹅膏菌属(Amanita,0.123%)、革菌属(Thelephora,0.600%)等为优势菌属。模式三治理下乳菇属(Lactarius,39.970%)、濕伞属(Hygrocybe,6.904%)为优势菌属。由于不同治理模式下土壤微生物优势菌属明显不同,推测在不同治理模式下土壤营养物质即土壤肥力不同,同时检测土壤理化性质发现在模式二治理下土壤具有较高的营养物质,因此模式二治理下土壤中的优势菌属在石漠化恢复过程中具有重要作用,这可为今后选择治理石漠化植被提供参考依据。
2.3.2 土壤真菌多样性
总体上来看,各组样品覆盖率均大于99%,说明样品的采样率足够获得最丰富的OUT(表3)。选择香农指数(Shannon)、辛普森多样性指数(Simpson)和Chao1指数,对不同样品在97%一致性阈值下的α多样性进行分析。从表3可以看出,3种治理模式下土壤微生物的香农指数分别为为5.371、5.908、3.946,辛普森多样性指数分别为0.941、0.957、0.858,Chao1指数分别为685、788、397。表明在模式二治理下土壤微生物具有更高的多样性,其次是模式一、模式三。
选用Weighted Unifrac距离和Unweighted Unifrac距离这2个指标来衡量样品之间的相异系数。由图7可知,以Weighted Unifrac距离作为标准(上方),内生细菌群落相异系数最大的为模式一与模式三,为1.867;其次为模式二与模式三,为1.756。结果表明模式一与模式三之间内生菌的物种多样性差异高于模式二与模式一、模式三的内生菌多样性。以Unweighted Unifrac距离作为标准(下方),相异系数最大的为模式一与模式三,为0.736,其次为模式一与模式二,为0.671。结果表明3种治理模式下土壤微生物群落结构具有差异。
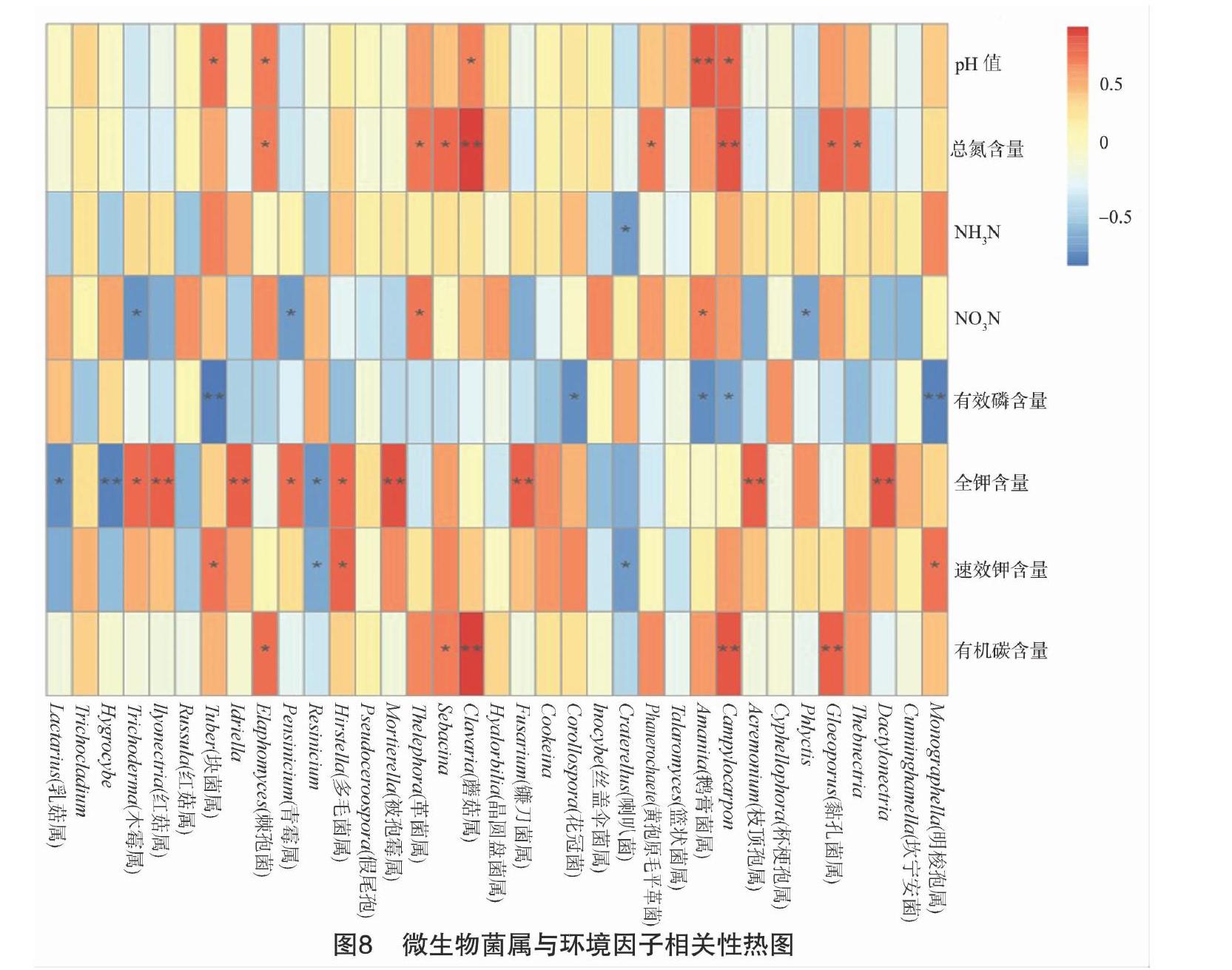
2.3.3 不同治理模式下土壤真菌群落与环境因子相关性
真菌属与环境因子相关性(图8)结果表明:有机碳含量与模式二下优势菌属Gloeoporus(0.800,P<0.01)、Campylocarpon(0.821,P<0.01)、
蘑菇属(Clavaria,0.928,P<0.01)具有极显著相关性。速效钾与模式二下优势菌属明梭孢属(Monographella,0.722,P<0.05)、多毛菌属(Hirsutella,0.780,P<0.05)、块菌属(Tuber,0.728,P<0.05)具有显著相关性。全钾含量与模式一下优势菌属枝顶孢属(Acremonium,0.800,P<0.01)、镰刀菌属(Fusarium,0.800,P<0.01)、Idriella(0.800,P<0.01)及Ilyonectria(0.800,P<0.01)具有极显著正相关性。硝态氮含量与模式二下优势菌属鹅膏菌属(Amanita,0.672,P<0.05)及革菌属(Thelephora,0.700,P<0.05)具有显著正相关性。全氮含量与模式二下优势菌属Campylocarpon(0.821,P<0.01)及蘑菇属(Clavaria,0.928,P<0.01)具有极显著正相关性。pH值与模式二下优势菌属鹅膏菌属(Amanita,0.808,P<0.01)具有极显著正相关性。有效磷含量与铵态氮基本上与所有菌属均无显著相关性。
由上述结果可看出,土壤中大多数营养物质均与模式二下土壤微生物菌属具有显著正相关性,说明在模式二的治理下土壤营养元素更多,养分更充足,更有利于植被的生长及石漠化的修复。综上所述,在模式二即阔叶林植被为壳斗类,杂灌的治理下,土壤的肥力更佳,石漠化治理能力更好。
2.3.4 真菌营养类型预测
营养类型预测结果(图9)表明,在模式一治理下的真菌主要为未分类腐生真菌(Undefined Saprotroph,27.249%),模式二下为未分类腐生真菌(Undefined Saprotroph,13.280%)及外生根菌(Ectomycorrhizal,6.786%),模式三下为未分类腐生真菌(Undefined Saprotroph,8.329%)及外生根菌(Ectomycorrhizal,57.461%)。腐生真菌能够分解落叶、动物粪便,在森林生态系统养分循环中具有重要作用。同时试验表明外生根菌能使树木提前4~5年成材,在发展林业育苗、促进树木的生长及绿化荒山等方面扮演着重要角色。这2种菌在3种治理模式下都属于优势菌,该菌有利于增加土壤肥力,促进植物生长及石漠化地区植被的恢复。
2.4 3种治理模式下土壤样品与环境因子相关性
在石漠化治理过程中,土壤营养物质的变化与石漠化恢复情况密切相关。在石漠化恢复过程中,土壤营养成分不断增加,土壤肥力增加,微生物生长繁殖快,有利于石漠化的恢复。图10结果显示,3种治理模式下土壤样品与环境因子的相关性。如模式一治理下的土壤样品与全钾、速效钾含量以及铵态氮含量呈正相关关系,模式二土壤样品与除有效磷含量外的全部环境因子都呈正相关关系,模式三则仅与有效磷和硝态氮含量具有正相关关系。此结果表明,模式二治理下土壤具有较高的肥力,而其他2种治理模式下土壤肥力由于受到环境因子的限制而低于模式二治理下,因此模式二治理下土壤的各营养物质总体上高于另外2种。
3 结论与讨论
贺祥等研究认为,土壤容重随着石漠化程度的加剧而增大,与原生林地相比,石漠化区域的土壤有机质、全氮等养分含量较低,速效养分含量与其相差不显著[28-29]。土壤中有机质、氮、磷、钾等养分含量与土地退化的演替过程一致,石漠化程度越高,土壤有机质、氮、磷、钾等养分含量越低。李孝良等研究认为,随着石漠化程度的增加,土壤全氮、有机碳含量均呈降低的趋势。微生物量碳与有机质之间有强烈的依赖关系,同时与土壤肥力紧密相关,可以作为指示肥力的重要指标[30]。土壤酸碱度受母质、生物气候及管理措施等条件的制约,是土壤肥力的重要影响因素之一。氮是成土过程中由于生物作用积累的,氮含量高意味着土壤中微生物的生物作用活跃。同时有机碳含量能直接影响土壤肥力,它们之间为正相关关系。磷参与组成植物体内许多重要的化合物,是植物体生长代谢过程不可缺少的。通过对不同治理模式下土壤样品的各理化性质进行分析比较得出,模式二治理下土壤的各营养物质整體上较其他的要高,说明不同模式的石漠化修复能力有差别,其中模式二的修复效果相对较好,土壤肥力较高。
不同治理模式下土壤微生物均表现为变形菌门、酸杆菌门以及疣微菌门相对丰度较高。物种相对丰度聚类热图结果显示在不同的治理模式下土壤中占优势的菌属各不相同,推测这可能是由于受环境因子影响的关系。土壤微生物物种不同可能是造成土壤肥力差异的原因,意味着在不同的治理模式下其土壤微生物群落结构的差异造成对石漠化的恢复能力不同。同时,将不同模式治理下的土壤与环境因子作相关性分析,结果表明在模式二治理下的土壤跟与土壤肥力相关的全氮、有机碳含量等都具有较高的相关性。
不同的治理模式造成石漠化地区土壤微生物群落结构不同,而不同的微生物群落结构对土壤的肥力即石漠化的恢复能力具有不同的效果。一系列的分析比较结果显示土壤中的优势菌属为罗尔斯通氏菌菌属、芽孢单菌属以及Gaiella的模式二即阔叶林植被为壳斗类,杂灌的治理下,土壤的肥力更佳,石漠化治理能力更好。
参考文献:
[1]Li B Q,Li Z,Sun X X,et al. DNA-SIP reveals the diversity of chemolithoautotrophic bacteria inhabiting three different soil types in typical karst Rocky desertification ecosystems in Southwest China[J]. Microbial Ecology,2018,76(4):976-990.
[2]Feng D L,Huang X H,Liu Y,et al. Growth and changes of endogenous hormones of mulberry roots in a simulated rocky desertification area[J]. Environmental Science and Pollution Research International,2016,23(11):11171-11180.
[3]Huang Q H,Cai Y L,Xing X S. Rocky desertification,antidesertification,and sustainable development in the karst mountain region of Southwest China[J]. Ambio,2008,37(5):390-392.
[4]Jiang Z C,Lian Y Q,Qin X Q. Rocky desertification in Southwest China:impacts,causes,and restoration[J]. Earth-Science Reviews,2014,132(3):1-12.
[5]苏维词,朱文孝,熊康宁.贵州喀斯特山区的石漠化及其生态经济治理模式[J]. 中国岩溶,2002,21(1):19-24.
[6]刘建忠,郭 颖,王六平,等. 贵州喀斯特石漠化地区生态修复现状及治理措施[J]. 安徽农业科学,2011,39(19):11684-11686.
[7]Qi X K,Wang K L,Zhang C H. Effectiveness of ecological restoration projects in a karst region of southwest China assessed using vegetation succession mapping[J]. Ecological Engineering,2013,54(2):245-253.
[8]李隽宜.不同石漠化治理模式的林分生长、植物多样性及土壤肥力比较[D]. 南宁:广西大学,2016.
[9]周晓果,李隽宜,朱宏光,等. 不同石漠化治理模式的造林保存率及幼林生长分析[J]. 广西科学,2017,24(2):175-181.
[10]温远光,夏承博,周晓果,等. 不同石漠化治理模式下群落的植物组成及多样性[J]. 广西科学,2017,24(2):168-174,181.
[11]龙海飞,苏维词,夏 春.石漠化地区土壤有机质与微生物数量关系研究[J]. 环境科学与技术,2013,36(10):57-62.
[12]Bardgett R D,van der Putten W H. Belowground biodiversity and ecosystem functioning[J]. Nature,2014,515(7528):505-511.
[13]Ling N,Zhu C,Xue C,et al. Insight into how organic amendments can shape the soil microbiome in long-term field experiments as revealed by network analysis[J]. Soil Biology and Biochemistry,2016,99(1):137-149.
[14]陈 莹,王 茜,王子苑,等. 不同石漠化程度对灌丛草地土壤细菌遗传多样性的影响[J]. 草地学报,2017,25(4):756-761.
[15]罗海波,宋光煜,何腾兵,等. 贵州喀斯特山区石漠化治理过程中土壤质量特性研究[J]. 水土保持学报,2004,18(6):112-115.
[16]唐 政,李继光,李 慧,等. 喀斯特土壤微生物和活性有机碳对生态恢复的快速响应[J]. 生态环境学报,2014(7):1130-1135.
[17]魏 媛.退化喀斯特植被恢复过程中土壤生物学特性研究——以贵州花江地区为例[D]. 南京:南京林业大学,2008.
[18]覃勇荣,苏 盛,黄小梅,等. 不同植被对石漠化地区土壤微生物数量的影响[J]. 河池学院学报,2016,36(5):1-8.
[19]覃勇荣,王 燕,刘旭辉,等. 马尾松对喀斯特石漠化地区土壤有机质的影响[J]. 中国农学通报,2009,25(5):104-109.
[20]秦平书.石漠化对马尾松幼林生长的影响[J]. 湖南林业科技,2015,42(6):82-85.
[21]陈 瑞,童方平,李 贵,等. 石漠化区枫香、刺槐和柏木的抗逆性初步研究[J]. 湖南林业科技,2014,41(6):18-22.
[22]董旭杰,彭继庆,曹福祥,等. 湘中、湘南石漠化地区植物多样性研究[J]. 中南林业科技大学学报,2015,35(12):102-107.
[23]司 彬,姚小华,任华东,等. 滇东喀斯特植被恢复演替过程中物种多样性研究[J]. 西南大学学报(自然科学版),2009,31(1):132-139.
[24]宋海燕,张 静,赵雅洁,等. 不同程度石漠化对金山荚蒾末端小枝的生长和生物量積累及分配的影响[J]. 植物科学学报,2018,36(1):103-111.
[25]马 骏,阚丹妤,沙 敏,等. 滇池流域石漠化地区植被恢复技术研究[J]. 安徽农业科学,2014(33):11782-11783,11807.
[26]顾秉谦,周明忠,肖时珍,等. 贵州施秉世界自然遗产地喀斯特地貌的地质控制研究[J]. 西南大学学报(自然科学版),2016,38(10):132-138.
[27]Zhang Y,Jin D C,Zhou Y F,et al. Community composition and diversity of soil oribatid mites(Acari:Oribatida)in karst forests in Shibing,Guizhou,Southwestern China[J]. Acta Entomologica Sinica,2015,58(7):791-799.
[28]贺 祥,熊康宁,陈洪云,等. 喀斯特山区生态治理区石漠化过程的土壤质量特征研究[J]. 云南师范大学学报(自然科学版),2008,28(2):58-64.
[29]彭 熙,黄 英,车家骧,等. 不同石漠化等级条件下土壤性状变化规律研究——以贵州喀斯特中心普定站为例[J]. 中国岩溶,2009,28(4):402-405.
[30]李孝良,陈效民,周炼川,等. 贵州省喀斯特地区石漠化土壤有机碳和全氮变异特征[J]. 南京农业大学学报,2010,33(4):75-80.
猜你喜欢
长江科学院院报(2023年8期)2023-08-29
内蒙古林业调查设计(2021年2期)2021-06-29
中国水土保持(2018年11期)2018-11-06
黑龙江教育学院学报(2016年12期)2017-01-13
科教导刊·电子版(2016年30期)2016-12-26
法制与社会(2016年33期)2016-12-15
企业导报(2016年19期)2016-11-05
学术论坛(2016年5期)2016-05-17
河北渔业(2015年5期)2016-03-01
