
作物的从头驯化:原理、进展及挑战
——以野生植物沙米的驯化为例
2021-07-24 07:15马小飞钱朝菊朱新宇
南通大学学报(自然科学版) 2021年2期
马小飞,燕 霞,钱朝菊,朱新宇
(1.南通大学 生命科学学院,江苏 南通 226019;2.中国科学院西北生态环境资源研究院 甘肃省寒区旱区逆境生理与生态重点实验室,甘肃 兰州 730000)
植物的引种驯化历史可以追溯至新石器时代,人类逐渐学会生产食物代替搜集食物,慢慢使得农业出现并发展,并促进了多元文化的兴起。植物的引种驯化不仅与古文明史的诞生和发展息息相关,还对人类社会精神文明和物质文明的进步及人类的发展有着不可替代而深远的影响。现存已知的40多万种植物中,被成功驯化的作物尚不超过100种,而且仅有15 种成为主要的粮食作物,其中水稻、小麦和玉米等三大作物占比高达50%。有研究表明,受气候变化的影响,全球的玉米、水稻、小麦和大豆等主要作物的总产量每年约减产三分之一[1]。传统作物的随机性驯化使得其遗传多样性远低于野生祖先种[2],导致作物对气候变化的适应能力较差。自21 世纪以来,世界人口急剧增加,气候变化剧烈,对粮食及其多样性的需求也与日俱增。为了改善这种现状,联合国粮食及农业组织在《应对气候变化:粮食和农业遗传资源的作用》中指出,养活世界日益增长的人口需要适应力更强的粮食原料,要筛选和驯化能够适应气候变化的小作物和野生物种,同时要大力保护遗传多样性,从而增强农作物对气候变化的适应性,进一步完善植物遗传资源基因库[3]。
1 作物的驯化理论发展
我国是植物引种驯化理论研究最早的国家之一。早在春秋时期,就出现“土宜论”,认为不同的农作物生长需要不同的土壤条件;在此理论基础之上,西汉后期的《氾胜之书》加入了适时播种、适时施肥、适时灌溉、适时收获等内容,进而发展成“风土论”。这两个理论虽然只考虑了环境因子对作物驯化的影响而忽略了植物本身对外界环境因子的适应性潜能和适应性进化,但是对我国古代的作物驯化依然起到了积极有效的指导和推进作用。在北魏和明代时期,人们开始意识到“风土论”的局限性,并在《农政全书》中强调了人为因素的重要性,即通过人为努力改造植物对环境的适应性,该理论逐渐发展成“排风论”。这个时期是我国古代作物驯化理论研究的一个高峰期[4]。
国际上对作物驯化一直存在不同的见解和看法。直到19 世纪,英国人达尔文在《物种起源》中提出,物种变异的普遍性和遗传性是驯化的基础。在此基础上,人们根据喜好去劣存优,使其按照人类预期的进化方向发展。该理论为后期的驯化实践提供了不可或缺的理论依据[5]。20 世纪以来,一些学者又陆续提出“气候相似论”“风土驯化学说”“植物地理学差率法”“栽培植物起源中心论”“生态因子季节节律同步论”“因素论”“生态综合分析法”等相关的作物驯化理论,极大地促进和推动了植物的引种驯化实践以及农业文明的发展[4,6-12]。随着科学技术的不断发展、驯化理论研究的不断深入和完善,人们普遍认为驯化主要是以进化论、遗传学及生态学等基础学科为理论依据,辅以植物学、育种学、植物生理学、植物地理学及植物栽培学等其他分支学科[13]。
所谓植物的从头驯化(de novo domestication),就是根据人类的自身需求,通过对优良性状关键基因的驯化,在较短时间内将自然界中的野生植物物种转变为家养品种的过程[14]。与传统驯化相比,从头驯化在时间上大大缩短了植物的驯化历程,传统驯化与从头驯化的比较见表1。通常,野生植物物种是否驯化成功都会依赖于人工种植收获过程中的某类或几类关键性状指标,例如在小麦、大麦和水稻等谷类作物中的不落粒性和种子休眠等[15-23]。在全球气候变化和人口快速增长的背景下,对作物的气候适应性及营养多样性的需求逐渐加大,特别是最近几十年随着传统作物驯化综合征基因的定位,基因组测序技术的飞速发展,以及进化理论的完善,植物遗传学家对野生植物的从头驯化热情受到了极大鼓舞。

表1 传统驯化与从头驯化的比较Tab.1 Comparison of traditional domestication and de novo domestication
2 作物从头驯化的进展及挑战
尽管驯化通常被描述为人类意图明确的选育结果,是人类在自然选择的基础上强化了定向驯化的能力[7-8],但是驯化是人类、环境、植物和动物之间连续不断地相互作用的复杂过程的结果。该过程通常受到生态、生物本身、生物多样性以及人类文化等多方面因素的驱动,是一个长期的过程[9-10]。传统的引种驯化效率常常会受到基因与环境间的相互作用(gene-by-environment)以及基因与基因间的相互作用(gene-by-gene)影响而降低[11]。此外,早期的选种驯化仅发生在较小的群体范围内,性状的选择范围有限,由于受基因流、遗传漂变等遗传因素以及种植技术和气候变化等因素的影响,植物驯化的不确定性和驯化所需的时间大大增加。例如,仅一个谷类作物关键驯化指标——不落粒性,在小麦、大麦和水稻中的驯化历史长达2 000~4 000 年[12]。这种漫长的驯化周期远远不能满足现代日益增长的人口和营养多样性对驯化作物的需求。另外,在人类长期的定向选择作用下,虽然高产优质等性状的有益变异在基因组中被固定下来,但是与野生种相比,由于受基因间共表达关系等效应的影响,驯化作物的遗传多样性和对环境变化的适应性大量丢失,严重阻碍了植物资源的多样性保护和可持续发展[15]。
近几十年,随着植物基因组的深入挖掘,人类陆续鉴定出作物驯化过程中一些重要的驯化基因,例如水稻、大豆和番茄的种子休眠基因Sdr4、OsG、G、SolyG[17-18],番茄果实大小基因SUN、fasciated、fw2.2[19-21]等。这些被挖掘出的驯化基因,通过基因工程技术手段,在野生种或是半驯化作物种中经过人工修饰用以调控关键的农艺性状,这样,在短短20代内就可以获得抗逆性强、产量高、品质好、性状稳定的作物新种质。这就是基于基因编辑技术的从头驯化。目前该方法已成功运用于野生番茄等野生植物的从头驯化和作物的遗传改良工作中,逐渐成为作物驯化和分子辅助遗传育种的主流方式[22-25]。虽然该方法大大提高了野生植物到栽培作物的驯化时间,见效快,但是只能基于个体或是较小群体水平。在新品种大面积推广时,往往会受局域适应性等因素影响使得农艺性状无法达到预期水平。而且在种植时,基于基因编辑技术改造的作物品种(暂归于转基因作物)还存在不确定性农业生态系统风险[26]。目前,转基因作物及其加工食品尚不能被大众广泛接受[27]。
如何解决传统人工选育的高耗时、低适应性以及基因编辑技术所带来的转基因恐慌等问题,是从头驯化过程中的难点和关键问题。本课题组通过广泛的群体基因型收集并通过同质园实验在大的有效群体中进行规范化种植、品种的选育、优良性状基因型间的杂交、分子标记技术确定品种的稳定性,总结出一套新的适合野生植物从头驯化的技术方法。该方法不仅比传统的人工选择育种大大节省时间,而且有效规避了转基因作物市场推广的问题。目前通过该方法,已经选育出数个适宜沙米驯化的品种(如图1 所示)。

图1 沙米从头驯化策略示意图Fig.1 Schematic diagram of A.squarrosum′s de novo domestication strategy
3 基于群体遗传学的沙米从头驯化
在我国,西北温带荒漠生态系统面积约占国土面积的六分之一[28-29],虽然生长在荒漠中的植被种类稀少且密度稀疏,但是在荒漠化土地防治和温带荒漠生态系统的稳定性维持中起着关键作用。并且,由于长期适应干旱、高温、营养贫瘠等极端恶劣的荒漠生境,荒漠植物基因组中积累了大量宝贵的抗逆遗传资源[30],改造或驯化能够适应这些极端生境的粮食和能源作物,已然成为生态农业建设的有效途径之一[31-32]。此外,西北地区由于其得天独厚的地理和气候优势而孕育了丰富的药食同源植物,例如沙米等,但是对于沙漠药食同源植物的开发和应用远不能满足现代社会发展的需求。西北温带沙漠拥有广袤的种植基地,是开展药食同源植物有机种植的最佳土地资源。同时,沙漠地区也是我国生态建设的主战场之一,开展荒漠植物的从头驯化工作和合理开发应用,不仅可以促进我国西北荒漠区的生态恢复,还可以带动西北荒漠区的绿色经济发展。
3.1 沙米简介
沙米学名沙蓬(Agriophyllum squarrosum(L.)Moq.),又名吉刺儿[蒙语]、蒺藜梗,是广泛分布于我国及中亚干旱-半干旱区流动沙丘的一年生先锋植物。作为沙蓬属(Agriophyllum)中分布最为广泛的一个种(http://www.eflora.cn/sp/Agriophyllum),沙米的生态适应性范围极广,在我国北方年降雨量为50~400 mm 和年平均温度为-1~10 ℃的各大沙漠以及沙地的贫瘠土壤上很常见[33],具有卓越的耐热和抗干旱能力,对全球气候变暖所带来的水热变化具有较强的适应能力[34],是防沙治沙的先锋物种和沙漠化逆转过程中的首要建群植物[35-37],更是沙漠贫瘠土壤中最重要的碳、氮等营养流的来源[38]。此外,沙米还具有丰富的地下种子库,在5—6 月期间遇适当降雨后便可迅速发芽,生长周期较短,通常10 月份即可成熟,并且其枯萎的枝叶依然残留在地表2~3 个生长季。单株立枯沙米至少可降低90%风速,能够有效阻挡风沙流形成沙舌,具有明显的防风固沙能力[35-36]。
除此之外,沙米还具有极高的营养和药用价值。沙米种子至少有1 300 年的悠久食用历史。早在唐代就成为了军队的供给粮,后期一度成为救荒植物[39]。沙米种子营养丰富,约含蛋白质23%、脂肪酸10%、碳水化合物45%,籽粒营养丰富全面,可以与“太空粮食”藜麦媲美[40]。相对于水稻和小麦等传统主要粮食作物,沙米种子中碳水化合物含量低(仅45%)不饱和脂肪酸含量高(9%),富含蛋白质(23%)和纤维素(8%),而且所含人体必需的8 种氨基酸比例与联合国粮食及农业组织推荐比例几乎完美吻合,是一种天然的高级营养食品[41],具有作为绿色食物开发的巨大潜力。中医认为沙米种子可作为减肥人群的主食选择,是糖尿病、心脑血管疾病、肾脏功能减退病人的理想食品[42-44]。目前在西北地区的美食中,沙米凉粉、沙米馍馍、沙米珍子面、沙米撒饭等深受人们的喜爱。另一方面,沙米植株的干燥地上部分可以入药,是蒙药的传统常用药材。作为特色蒙药资源,沙米植株具有祛疫、清热、解毒、利尿之功能[43]。用其煮散剂或入丸散可治疗瘟疫、头痛、赤目、黄疸、肾热、尿道灼痛、胃“赫依”、口舌生疮、齿龈溃烂等症[44]。同时沙米还含有丰富的抗氧化、抗肿瘤、抗炎、抗菌、降血压、降血脂和保护心脑血管等功能化合物[42]。研究表明,沙米粗寡糖对糖尿病,肝、肾损伤有良好的抑制作用或延缓病情恶化的作用,且能降低血糖、调节血脂和增加胰岛素敏感性[45-46]。沙米全草含有较多药理活性强的物质[47],如:生物碱类化合物可扑热息痛,具有抗炎、解热和镇痛作用;紫罗兰酮具有抗炎、抗氧化和化学防癌的作用;水杨苷具有解热、镇痛以及昆虫据食性作用;黄酮类物质及其苷类化合物具有较强的降血糖作用[48]。虽然沙米被列入《中国药用植物》名录,但是由于沙米独特的地理分布使得人们对沙米的关注度和整体认识较低,制约了沙米作为药食兼用植物在现代生态农业中的进一步发展以及育种专家对其的认知,沙米的驯化工作目前也仅限于西北地区相关的科研工作者之间。
3.2 沙米的自然分布区
分布区调查结果显示沙米的生境范围很广,在塔克拉玛干沙漠、古尔班通古特沙漠、库姆塔格沙漠、巴丹吉林沙漠、腾格里沙漠、柴达木盆地、雅江河谷、乌兰布和沙漠、毛乌素沙地、库布齐沙地、科尔沁沙地和浑善达克沙地等全国各大沙漠均有分布。自2013 年起,根据中国各标本馆、地方植物志的标本记录以及野外实地考察标记的我国沙漠/沙地沙米的自然分布区记录,本课题组进行了沙米野外资源调查以及种质资源的采集。目前本课题组收集的沙米基因型达1 000 多个,囊括我国西北部各大沙漠和沙地群体,为沙米的优良性状选择提供了充足的样本。
3.3 沙米的表型多样性及驯化性状的确定
植物的表型变异是遗传变异与环境异质性共同作用的结果,也是作物驯化过程中性状筛选的首要依据之一。通过同质园实验(common garden experiment)将原先在不同生境居群的个体引种并栽植在同一园地,让其保持生境一致,可以剔除环境不均一性造成的表型变异中的居群内个体差异或居群间差异(环境饰变作用),从而可以单独考察遗传因素对表型的效应。通过多年的摸索,我们已经制定出一套适宜沙地的沙米规模化种植技术体系,不仅可以用于同质园实验,更适用于沙田大范围种植。
前期,通过对采集自不同种源的26 个沙米群体的表型特征和生殖策略的调查,统计分析了包括植株高度、茎直径、基部分枝长、地上生物量、收获种子重量和种子粒径等重要生态农艺性状的变异模式[49],结果如图2 所示。研究发现,南部群体的地上生物量较高,适宜于沙米作为牧草等的驯化,而东北的沙米群体其籽粒粒径和千粒重均显著大于南部的群体,但是生物量则较低,适宜于沙米作为食用作物的驯化;另一方面,与东北的沙米群体相比,南部群体的表型多样性更高,具有较高的种源异质性,为沙米的从头驯化提供了多样的表型性状,可根据相应的驯化目的选择适宜的驯化类群[49]。
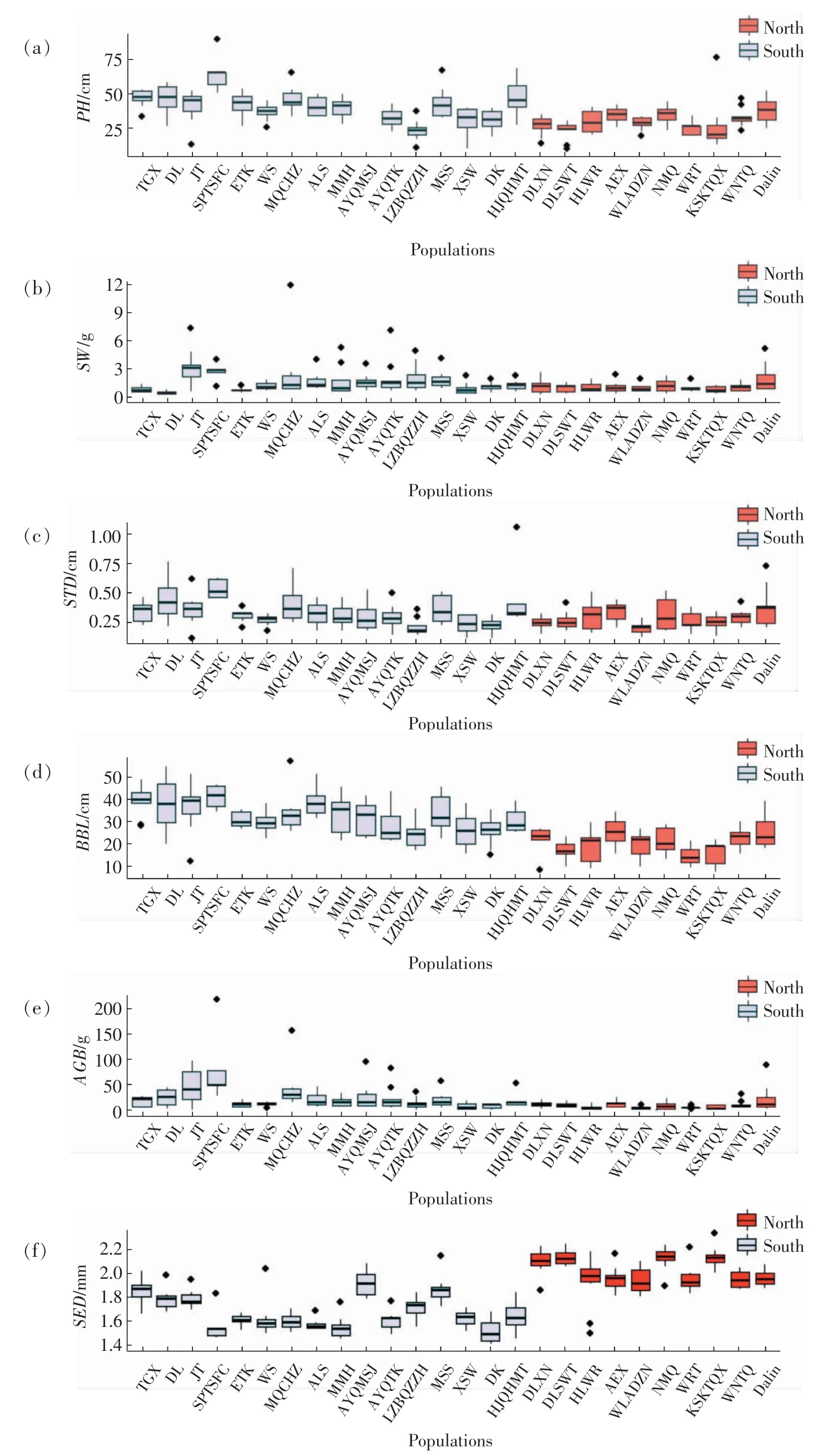
图2 沙米种群表型多态性分析[49]Fig.2 Analysis of phenotypic polymorphism of A.squarrosum population[49]
为了更好地筛选和利用沙米的优良农艺性状,本课题组利用Canoco 软件将沙米的表型变异与种源气候因子进行关联分析。结果显示,沙米表型性状随纬度变化明显,具有显著的纬度地带性。同时,随纬度升高,种子粒径逐渐增大,而其他表型性状则相反(图3)。通常,纬度变异主要伴随着温度的变化,而经度变异则主要伴随着降雨量的改变。因此,沙米的表型多样性总体上与温度相关性最大,其次是降水量。年均温变化和温度季节性变化越大则越有利于单粒种子的繁殖生长;降雨量和降雨量变异系数越小越有利于沙米营养生长[49]。通过计算比较,发现温度和降雨的变异不能完全解释沙米的表型变异。将沙米的表型变异与风速进行相关性分析发现,种子粒径与秋冬季(10 月—次年1 月)最大风速显著正相关,而基部枝长则与夏季生长季(7、8 月)的最大风速呈正相关。这说明季节性风力大小影响了沙米的表型分化[50],这为选育不同风沙区的沙米种质资源提供了重要参考。
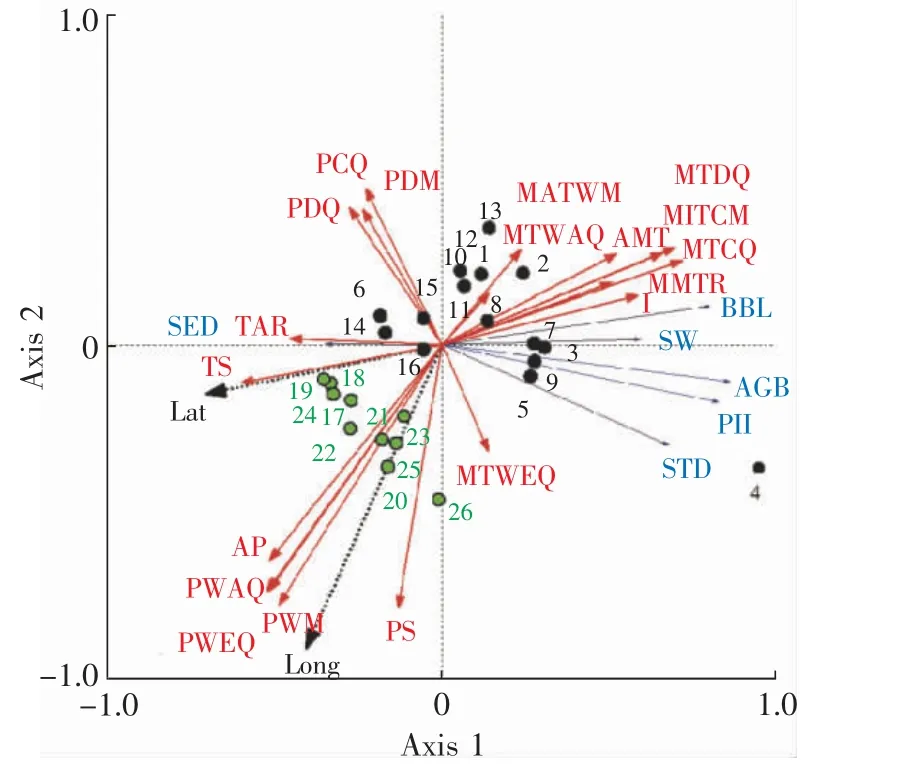
图3 基于26 个沙米居群的RDA 双标图[49]Fig.3 RDA double plot based on 26 A.squarrosum populations[49]
同质园表型监测结果表明,武威、铁盖乡和阿尔乡群体的农艺性状明显优于其他种源的群体,例如:武威群体具有稳定而较大的地上生物量,非常适合驯化成为牧草品种(图4(a));铁盖乡群体的株型整齐且穗长,易于农田种植和管理(图4(b));阿尔乡群体种子粒径大且多,适宜驯化为产种的优良品种(图4(c))。经过多年的大田表型观测和优良性状品系人工选育,这三个群体中优良性状品系经过人工选育后的品种各项性状比野生型均有显著的改良且性状能够稳定遗传,其中单株最大种子产量约100 g。目前,已筛选出生物量大(WW10)、种子粒径大(AEX3)和穗长(TGX9)3 个优良性状的地方品种(图4)。

图4 沙米人工选育品种Fig.4 Artificial selection of A.squarrosum varieties
3.4 沙米的谱系地理和遗传多样性
由于种群动态历史等会对现有物种的遗传多样性的模式和水平、基因组内的结构变异以及基因间的连锁关系等产生一定的影响,会直接或间接影响野生植物的从头驯化过程等[51],因此,在进行植物的从头驯化时,需要对其谱系地理和种群动态历史进行研究从而更好地指导驯化工作。Qian 等[52]人通过对广泛分布于我国北方干旱、半干旱区沙丘上的一年生先锋植物沙米的现存分布区的全面调查,采集了包括塔克拉玛干沙漠、古尔班通古特沙漠、库姆塔格沙漠、巴丹吉林沙漠、腾格里沙漠、柴达木盆地、雅江河谷、乌兰布和沙漠、毛乌素沙地、库布齐沙地、科尔沁沙地和浑善达克沙地等典型沙化土地区域的46 个群体共188 个沙米样本,并利用5 个母系遗传的叶绿体基因(cpDNA)和一个双亲遗传的核糖体内部转录间隔区片段(nrITS)的序列变异式样,构建了沙米的谱系结构,并模拟其群体动态历史和不同历史时期的分布区变化。结果表明:沙米大约在160 万年前起源于古尔班通古特沙漠,随后在几次冰期期间,随着亚洲内部干旱化的加剧,沙米通过3 次大的定植过程扩散至其他沙漠及沙地区域,分布范围广阔,资源丰富。与其他区域的种群相比,中部沙米群组(包括库姆塔格沙漠、巴丹吉林沙漠、腾格里沙漠、柴达木盆地、雅江河谷、乌兰布和沙漠、毛乌素沙地、库布齐沙地)的遗传多样性最高(图5);并且处于季风区的沙米有效群体的快速增长(图6),印证了全球气候变化对不同荒漠区域稳定性的影响不尽相同[53],为沙米的种质资源的筛选和收集提供了理论指导。此外,分子谱系揭示青藏高原的沙漠化可能始于110 万年前左右,早于先前的风尘记录。在全球变暖背景下,沙米的潜在分布区将向高亚洲区域扩张,青藏高原也存在沙漠化的巨大风险(图7),这也暗示着沙米作为未来粮食作物的巨大潜力和前景。
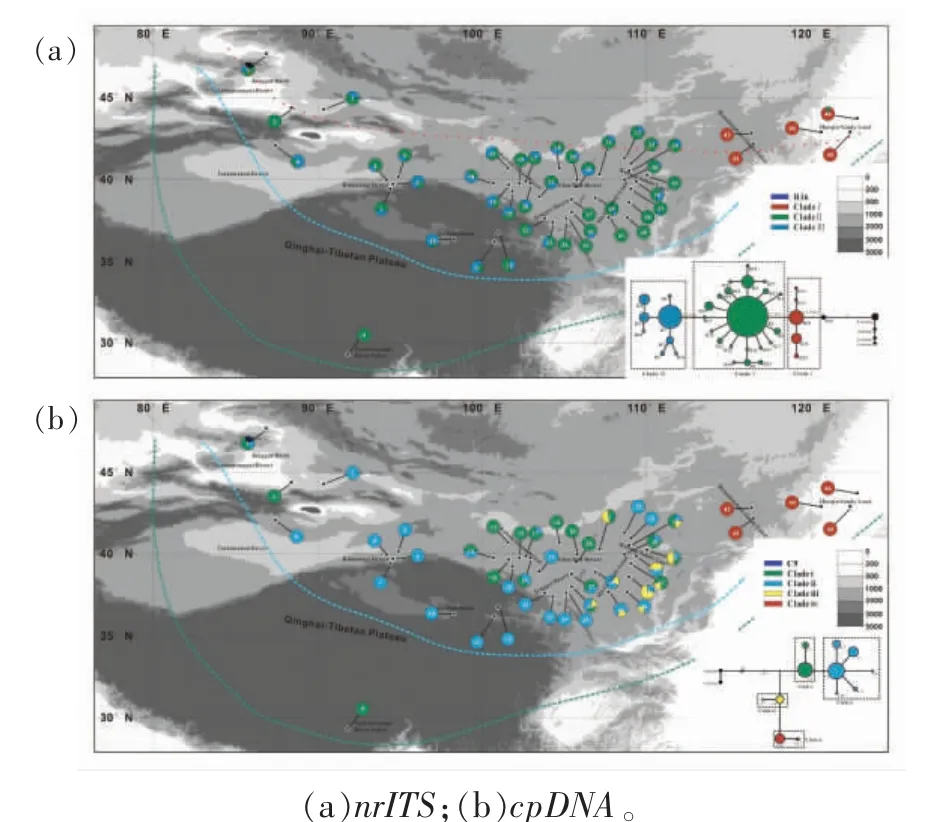
图5 沙米群体采样位置、nrITS 和cpDNA 单倍型分布及拓扑结构[52]Fig.5 Distribution of haplotypes in the geological locations and Median-joining networks of the haplotypes in the 46 A.squarrosum populations examined in this study[52]
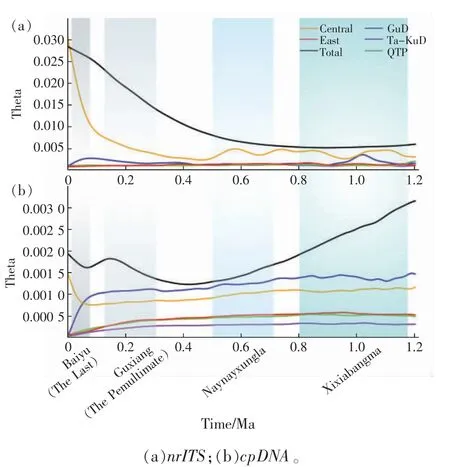
图6 基于nrITS 和cpDNA 数据模拟沙米有效群体大小动态历史变化[52]Fig.6 Skyline plot estimated by Migrate-n for each group of A.squarrosum based on nrITS and cpDNA data[52]
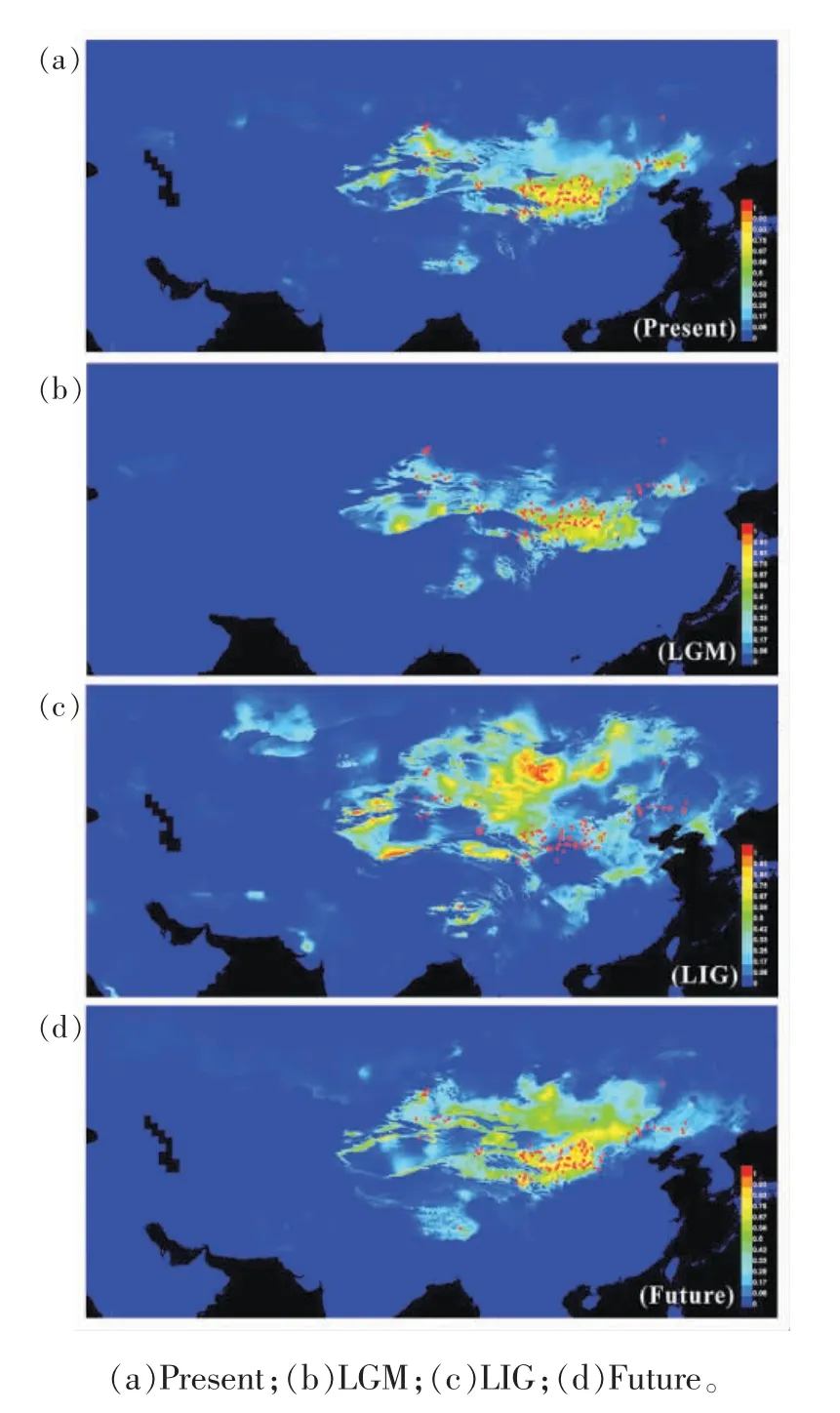
图7 沙米不同时期的分布区模拟[52]Fig.7 Predicted distributions of A.squarrosum[52]
该谱系地理工作通过对草原生态系统与沙漠生态系统转化过程中的典型先锋植物的分子谱系调查,不仅为理解荒漠生态系统对季风气候的响应提供了新的分子数据支持和解析角度,还为选育应对全球气候变暖的“未来作物”——沙米提供了生态适应性潜力和种质资源遗传多样性评估。
3.5 分子辅助育种指导沙米的从头驯化
植物的表型性状通常是由环境和基因共同作用的结果。特定的分子标记可以快速筛选出抗逆性强、性状优良的基因型和品种。分子标记辅助育种通过分析与目标基因紧密连锁的分子标记的基因型,借助分子标记对目标性状基因型进行选择,从而可以快速准确地筛选出目的性状,提高育种效率,缩短育种年限。在大田人工选育优良性状的基础上,借助分子标记可精确进行性状筛选,鉴别亲本亲缘关系,回交育种中数量性状和隐性性状的转移、杂种后代的选择、杂种优势的预测及品种纯度鉴定等,同时又避免了市场对转基因育种的不信任与恐慌。因此,为了进一步快速筛选和培育更加优良的沙米品种,本课题组正在通过群体遗传学方法从多方面深入挖掘沙米基因组中与抗逆性、茎秆成药性、落粒性以及粒径大小等农艺性状相关的基因或基因调控途径,并在此基础上确定相应的分子标记用于沙米的从头驯化,具体如何实现沙米的从头驯化可以分为以下几个方面:
首先,沙米优良作物性状的筛选及农艺性状相关基因精确定位。全基因组关联分析(genome-wide association study,GWAS)[54]是通过应用基因组中数以百万计的单核苷酸多态性(single nucleotide polymorphism,SNP)为分子遗传标记,进行全基因组水平上的对照分析和性状等相关性分析,从而定位影响复杂性状的基因变异的方法,现已广泛应用于人类疾病研究及动植物分子辅助育种。为了能够更好地指导沙米的从头驯化,现在正在进行沙米的基因组测序及群体基因组重测序,并期望通过GWAS 分析以及QTL(quantitative trait locus)定位等分子手段,精细挖掘沙米基因组中的农艺性状相关基因,勾画沙米目标驯化性状的遗传调控机制,从而精确辅助沙米优良品种的筛选和杂交培育。例如,植物的落粒性是谷类作物是否驯化成功的一个显著性标志。通过大范围的筛选和表型监测,本课题组已发现两个落粒性较差的沙米基因型,为沙米的从头驯化研究提供了可靠的材料。
其次,沙米的抗逆性筛选以及抗逆品系选育。沙米广泛分布于我国北方年降雨量为50~400 mm的各大沙漠和沙地[33],具有卓越的抗旱能力。为了深入挖掘沙米基因组中的干旱适应性调控网络及关键干旱胁迫响应基因,基于对不同干旱程度生境来源的两种生态型沙米(AEX:年降水量约400 mm;WW:年降水量约100 mm)进行干旱胁迫对照下的表型监测以及比较转录组学分析。表型监测结果显示:与AEX 生态型相比,WW 生态型的抗旱性更强。转录组数据分析表明:干旱胁迫下AEX 生态型和WW 生态型中分别检测出23 656 和24 994 个干旱响应基因。GO(gene ontology)富集分析表明:在严重干旱胁迫下,两种生态型均诱导了与细胞内部分、基因表达和核糖核蛋白复合物生物发生过程有关的基因显著上调,同时抑制了与光合作用和催化活性过程相关基因的活性。通过进一步的共调控网络分析表明,重度干旱下诱导上调的基因能够显著促进涉及光合作用和蛋白质合成相关过程调控网络的形成。比较转录组分析进一步表明,有98 对直系同源物的Ka/Ks值显著大于1.5,且大多数基因都是参与生物及非生物胁迫压力响应的基因,后期将通过GWAS 对沙米干旱胁迫响应方式及其调控方式进行深入的挖掘和验证,为沙米的从头驯化提供抗逆遗传资源(数据尚未发表),为定向基因编辑以及杂交育种等分子手段驯化高抗逆沙米品种提供基础数据。
最后,沙米的药用价值筛选及药用品系改良驯化。前期通过比较代谢组学,已经确定沙米茎秆中含有大量的药用活性成分[55],尤其是异鼠李素含量非常高;通过抑菌实验进一步验证沙米地上组织中的黄酮粗提物对尖孢镰刀菌(Fusarium oxysporum)、腐皮镰孢(Fusarium solani)和链格孢菌(Alternaria alternata)等植物致病菌具有显著的抑制作用(数据尚未发表)。通过群体代谢物差异分析和比较转录组分析,现已确定生长于高海拔的某些基因型地上部分的药用活性成分尤其是黄酮类物质含量显著高于其他基因型,并初步勾画出沙米道地性形成的分子代谢基础,为药用型沙米的从头驯化提供了优势类群和品种。后期将进一步通过代谢组学全基因组关联分析(metabolome genome-wide association study,mGWAS),深入探讨沙米的成药道地性形成的分子代谢基础,并挖掘沙米药用活性成分相关调控基因或基因网络,通过分子辅助育种和杂交育种等方式筛选和培育出药用活性成分高的沙米品种。
4 总结与展望
植物的引种驯化与人类文明发展及人类粮食安全息息相关,植物的成功引种驯化能够显著促进社会文明的进步。随着测序技术的发展及分析技术的日益精进,基于基因编辑技术的植物从头驯化方法不仅可以在短短20 几代就能获得性状优良、抗逆性高的作物,而且还可以避免传统人工选择进行植物从头驯化过程中作物的遗传多样性丢失和基因组中有害突变积累等问题。但是,随着粮食安全问题日益突出,转基因技术改造的作物品种尚存在不确定性农业生态系统风险,需要更加安全地进行植物的从头驯化。对植物的谱系地理、表型多样性、遗传多样性、基因组结构变异特征以及遗传—表型—环境关联性等问题进行全面了解和剖析,能够更好地指导该物种的从头驯化。此外,新型粮食和药用植物的探寻和筛选也是有效解决粮食安全问题的途径之一。通过沙米广泛的群体种质资源的收集、同质园实验进行沙米的规范化种植、品种的选育、优良性状基因型间的杂交等步骤进行沙米的从头驯化,不仅避免了传统驯化耗时费力,还解决了市场对转基因作物的不认可。这一从头驯化策略将为野生植物的从头驯化提供新的研究思路和方法。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
四川蚕业(2022年2期)2022-11-19
中华医学图书情报杂志(2022年1期)2022-11-18
广东农业科学(2022年7期)2022-09-14
中国现代医生(2022年21期)2022-08-22
农村科学实验(2022年2期)2022-03-12
华人时刊(2021年21期)2021-03-09
三农资讯半月报(2020年2期)2020-03-09
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
