
基于两性生命表和年龄-阶段捕食率的南方小花蝽对西花蓟马的控制作用
2021-07-21 10:31胡昌雄范苇张倩陈国华殷红慧徐天养杨进波杨航吴道慧张晓明
中国农业科学 2021年13期
胡昌雄,范苇,张倩,陈国华,殷红慧,徐天养,杨进波,杨航,吴道慧,张晓明
基于两性生命表和年龄-阶段捕食率的南方小花蝽对西花蓟马的控制作用
胡昌雄1,范苇1,张倩1,陈国华1,殷红慧2,徐天养2,杨进波3,杨航3,吴道慧1,张晓明1
1云南农业大学植物保护学院/云南生物资源保护与利用国家重点实验室,昆明 650201;2云南省烟草公司文山州公司,云南文山 663000;3云南格瑞生物科技有限公司,云南弥渡 675602
【】西花蓟马()是我国重要农业入侵害虫,南方小花蝽()是其优势捕食性天敌。论文旨在系统评价南方小花蝽对西花蓟马的控制作用以及同一地区两个种群的生物学差异。采集露地辣椒上的南方小花蝽和西花蓟马在室内条件下饲养2—3代后,用带花的辣椒嫩梢饲喂西花蓟马,用西花蓟马2龄若虫饲喂南方小花蝽进行试验。采用两性种群生命表和特定年龄-阶段捕食率的研究方法,系统测定南方小花蝽和西花蓟马云南种群的生长发育、繁殖力、捕食率以及预测种群数量增长模型。南方小花蝽和西花蓟马的不同虫期之间均存在不同程度的发育阶段重叠现象,且两种昆虫的雌成虫存活率均高于雄成虫。南方小花蝽若虫共5个龄期,而西花蓟马为4个龄期,南方小花蝽具有更长的若虫历期和更短的成虫寿命,且各发育阶段的存活率均低于西花蓟马,尤其是雌成虫的存活率要明显低于西花蓟马。南方小花蝽由卵成功发育成为雌成虫和雄成虫的概率分别为32.67%和20.67%,西花蓟马分别为46.67%和16.67%,西花蓟马具有更高的雌性比例。南方小花蝽和西花蓟马的繁殖参数(x)在整个繁殖期内具有高峰,分别在整个发育时间的22 d和16 d出现。南方小花蝽平均单雌产卵量为42.00粒,低于西花蓟马的平均产卵量59.86粒,其种群的净生殖率(0)、总繁殖率()、内禀增长率()、周限增长率()均低于西花蓟马,而平均世代周期()和种群加倍时间()长于西花蓟马。种群预测结果显示经过90 d后西花蓟马的个体数量达南方小花蝽的9.66倍,其中雌成虫数达到17.15倍。南方小花蝽对西花蓟马2龄若虫的捕食率随龄期逐渐升高,其种群净捕食率(0)为140.81,转化率(p)为9.05。南方小花蝽雌、雄成虫在整个发育阶段对西花蓟马2龄若虫的平均捕食量分别为159.67和86.00头。以西花蓟马为食物的南方小花蝽个体能较好地完成生活史且种群增长稳定;在相同条件下西花蓟马比南方小花蝽具有更强的种群增长潜力。南方小花蝽可作为西花蓟马生物防治的重要天敌,在利用过程中需要注意在西花蓟马暴发前10—20 d释放,并在西花蓟马持续暴发过程中连续释放可达到更好的控制效果。
南方小花蝽;西花蓟马;两性生命表;捕食率;生物防治
0 引言
【研究意义】西花蓟马()属于缨翅目(Thysanoptera)蓟马科(Thripidae),是一种危害严重的全球性害虫[1-2]。西花蓟马是我国的重大农业入侵害虫,在云南、广州、山东等蔬菜花卉种植地区均有发生,除了对植物直接取食危害外,还可传播多种重要植物病毒造成更大危害,这些病毒包括番茄褪绿病毒(tomato chlorotic spot virus,TCSV)和番茄斑萎病毒(tomato spotted wilt virus,TSWV)等[3-4]。同时,西花蓟马发育历期短、繁殖力强也给防治带来挑战[5]。成虫及1、2龄若虫主要危害植物的叶片、花、果实,尤其是2龄若虫有更高的取食量[6]。目前,对西花蓟马的防治方法包括捕食性天敌防治、微生物防治和粘虫板诱集防治等,但化学防治仍然是最有效和普遍的方法[7]。长期大量、不科学地使用化学杀虫剂给环境造成一定影响,并使西花蓟马产生抗药性,其种群逐渐失控[8-9]。因此,探究天敌与猎物间的生物学特性及捕食关系,开展西花蓟马的生物防治愈发重要。【前人研究进展】在西花蓟马的防治中,环境友好型防治方法一直是研究者普遍寻求的方法[10]。其中,生物防治被认为是较有潜力的防治措施[11]。西花蓟马的捕食性天敌包括捕食螨、捕食蝽、瓢虫等[12]。南方小花蝽()属于半翅目(Hemiptera)花蝽科(Anthocorida),是一种重要的捕食性天敌昆虫,对蓟马、蚜虫等多种小型害虫具有较强的捕食能力[13]。在全国范围内广泛分布,是我国南方农田中的优势捕食性天敌类群之一[14]。在田间采用南方小花蝽对朱砂叶螨()和棉花花蓟马()的控制作用为70%—80%[15]。在室内条件下南方小花蝽对花蓟马的捕食量达15.79头/d[16];郅军锐等[17]研究发现,南方小花蝽5龄若虫对蚕豆蚜()成虫的捕食能力强于雌成虫,而对西花蓟马的捕食能力则相反。以上研究结果表明,南方小花蝽在农田系统中对常见小型害虫的防治具有重要的应用价值。【本研究切入点】南方小花蝽作为西花蓟马生物防治资源的开发和利用越来越受到重视,明确南方小花蝽整个生长期对西花蓟马若虫和成虫的捕食能力、测定两种昆虫生命表参数,是系统评价南方小花蝽对西花蓟马控制能力的基础。在天敌对猎物捕食系统研究中,从捕食者生命表、天敌对猎物捕食率、猎物生命表、消耗率以及寄主植物等不同级别递进关系入手,可更好地评价天敌的控害效果[18]。根据Chi[19]的两性生命表理论,年龄-阶段两性种群生命表可以弥补传统生命表以昆虫平均发育历期为计算的不足,能综合考虑雌、雄成虫个体间发育速率和性别比例的差异,从而更加全面和准确地描述种群信息。【拟解决的关键问题】测定南方小花蝽不同龄期若虫和成虫对西花蓟马2龄若虫的捕食能力,并以辣椒和西花蓟马作为唯一的食物来源测定害虫和天敌的寿命、繁殖力、发育时间和南方小花蝽全发育期总捕食量等,为南方小花蝽保护、利用以及西花蓟马的防治提供理论依据。
1 材料与方法
1.1 供试昆虫
于2018年3月至2019年7月开展试验,南方小花蝽和西花蓟马于2018年采自云南省昆明市盘龙区大摆村的辣椒上,在云南农业大学云南生物资源保护与利用国家重点实验室的温室内,利用辣椒()和菜豆()饲养2—3代后进行试验。试验挑选行动敏捷,个体大小一致且虫龄相同的南方小花蝽,接入玻璃罐(d=10 cm,h=15 cm)内,后接入西花蓟马置于人工气候箱内(博迅,BICC-300,上海博迅实业有限公司。温度:(26±1)℃,相对湿度65%—75%,光周期L﹕D=16 h﹕8 h)备用。
1.2 西花蓟马和南方小花蝽生命表测定
1.2.1 西花蓟马生命表测定 发育历期的测定参照胡昌雄等[20]的方法并有所改进,将养虫笼中种植的无蓟马危害的开花辣椒植株移入蓟马养虫笼(1.2 m×1.2 m×1.2 m)中,每笼移入10盆共30株,让蓟马成虫在其花上产卵24 h。移走所有辣椒花上的蓟马成虫,采集带花的辣椒嫩梢转移到10个玻璃培养皿(d=15 cm)中,用塑料保鲜膜封口,在封口膜上用昆虫针扎孔以保持空气流通。后把培养皿放入气候箱内,发现孵化出若虫则记录来估算卵期。分别在每天上午和晚上8点在解剖镜下观察若虫孵化情况。将刚孵化出的1龄若虫转移到小培养皿(d=3 cm)内,后放入带花辣椒嫩梢,并在柄部裹上湿润棉花,培养皿盖子盖好后在周围缠绕上保鲜膜防止其逃逸。每24 h观察并记录蓟马的发育和存活情况,并更换一次辣椒嫩梢。直到所有蓟马死亡,记录蓟马雌雄性比。试验选择50头蓟马,并重复3次,共150头。
西花蓟马繁殖力测定:将上述试验中刚羽化的西花蓟马雌成虫挑出,接入小培养皿(d=3 cm)中,后接入一头同天羽化的雄成虫。同时放入柄部裹有湿润棉花的带花辣椒嫩梢。每12 h观察记录一次并24 h更换一次辣椒嫩梢。更换出的辣椒继续在新的培养皿中保留4 d至卵孵化,并对初孵的1龄若虫进行计数以此估算成虫的产卵能力,直至成虫自然死亡。
1.2.2 南方小花蝽生命表测定 南方小花蝽发育历期测定参照Liu等[21]的方法并有所改进,将温室种植长至10—15 cm高的辣椒苗放入南方小花蝽的饲养笼中,让其产卵24 h后取出辣椒苗,将辣椒苗放置在人工气候箱中进行培养。待卵孵化后,随机选择50只1龄若虫。单头小花蝽放置在小培养皿中,每24 h接入50头西花蓟马2龄若虫进行饲养。每12 h在解剖镜下观察记录其发育情况,记录每只昆虫的存活情况和若虫龄期,待小花蝽羽化后统计雌雄性比。另外,为测定南方小花蝽繁殖力,将同天羽化出的小花蝽雌、雄成虫单对放置在一个饲养瓶(底直径10 cm,高15 cm)中,接入60头西花蓟马并放入一株带盆的辣椒苗植株(苗高10 cm),让其产卵。每天记录存活情况和产卵数,辣椒苗上的卵粒在解剖镜下计数。设置3个重复,共150头。
1.3 南方小花蝽发育期取食量测定
选择在田间数量较大、危害较为严重且在植物上部取食的西花蓟马2龄若虫饲养南方小花蝽。取初孵化的南方小花蝽1龄若虫单头放入玻璃管(d=3 cm,h=10 cm)内。接入50头蓟马2龄若虫,每24 h记录南方小花蝽的生长情况,同时记录蓟马死亡数来判断取食量。空白对照不接入南方小花蝽,仅有50头蓟马。共挑取30头南方小花蝽初孵若虫进行试验。
1.4 数据分析
1.4.1 生命表计算 两种昆虫的生命表数据根据齐心等[22]提出的年龄-龄期两性生命表理论,150头南方小花蝽和150头西花蓟马的发育数据采用计算机程序TWOSEX-MSChart进行处理(时间单位0.5 d)。分析和计算特定年龄-阶段存活率xj(为按龄期划分的单位间距,为特定阶段,下同)、个体从卵发育到年龄的存活率(x)、特定年龄-阶段寿命期望值(xj)、特定年龄-阶段繁殖力(x)、特定年龄繁殖力(x);分别计算其种群参数净生殖率(0)、平均世代周期()、内禀增长率()、周限增长率()、总繁殖率()、种群加倍时间()。两种昆虫种群生命表参数用Bootstrap方法进行估计推断平均值和标准误,其中Bootstrap的设置次数为10 000次[23]。
1.4.2 南方小花蝽和西花蓟马的种群增长预测 采用计算机程序TIMING-MSChart,以最初10粒卵来预测南方小花蝽和西花蓟马90 d的种群增长和种群结构[22]。
1.4.3 南方小花蝽对西花蓟马的捕食率分析 根据年龄-阶段捕食率理论,30头南方小花蝽单位时间(24 h)取食西花蓟马数量的原始数据采用计算机程序CONSUME-MSChart进行分析,参照Xu等[24]计算特定年龄捕食率(x)、特定年龄净捕食率(x)、种群净捕食率(0)、特定年龄-阶段捕食率(xj)、转化率(p)等捕食率相关参数。用Bootstrap方法估计捕食率参数的平均值与标准误,其中Bootstrap次数为10 000次。
两种昆虫的各项试验指标数据用MS Excel 2019进行统计,试验图片绘制均在Origin 2018中进行。
2 结果
2.1 南方小花蝽和西花蓟马的发育历期
南方小花蝽捕食西花蓟马2龄若虫后能完成其发育,但是各龄期的历期不同。南方小花蝽共5个若虫期,其中5龄(3.32 d)最长,2龄(1.95 d)最短;西花蓟马若虫期分为4个龄期,其中1龄(2.96 d)最长,3龄预蛹期(1.38 d)最短。南方小花蝽从卵发育至成虫平均16.25 d,西花蓟马平均仅需要11.34 d。两种昆虫均表现出雌成虫寿命长于雄成虫寿命,且成虫的寿命在两种昆虫中存在明显差异,西花蓟马成虫具有较长的寿命,其雌成虫平均寿命为18.35 d,南方小花蝽的雌成虫平均寿命仅为12.78 d。两种昆虫全发育期寿命基本一致,但西花蓟马具有更短的未成熟虫期和更长的成熟虫期,未成熟虫期时间占整个世代的42.20%,而南方小花蝽未成熟虫期时间占整个世代的55.92%(表1)。

表1 南方小花蝽和西花蓟马的发育历期和成虫寿命
2.2 南方小花蝽和西花蓟马的存活率
两种昆虫因个体间不同虫态的生长发育速率存在差异,南方小花蝽和西花蓟马的不同虫龄之间均存在不同程度的发育阶段重叠现象(图1)。两种昆虫发育阶段的重叠主要发生在相邻虫态,并且由于部分个体在低龄若虫期时发育时间较短,高龄若虫、成虫的发育阶段重叠更明显;南方小花蝽成虫与4—5龄若虫具有更长的重叠时间,为6.5 d,其重叠时间相比于西花蓟马长4 d。两种昆虫的所有虫态中,除卵期外,其余各个发育阶段特定年龄-阶段存活率值(xj)均随发育时间的增加表现出先增加后下降的趋势。南方小花蝽雌成虫最早羽化时间为14 d,雄成虫最早羽化时间为13.5 d。两种昆虫不同发育阶段的存活率中,西花蓟马1龄(最高99.33%)和2龄若虫(最高82.00%)的存活率明显高于南方小花蝽1龄(最高87.33%)和2龄(53.33%)若虫的存活率。在南方小花蝽和西花蓟马成虫羽化后,两种昆虫雌成虫的存活率均明显高于雄成虫,且西花蓟马雌成虫的存活率高于南方小花蝽雌成虫存活率。南方小花蝽由卵成功发育为雌成虫和雄成虫的概率分别是32.67%和20.67%,西花蓟马卵成功发育为雌、雄成虫的概率分别为46.67%和16.67%(图1)。
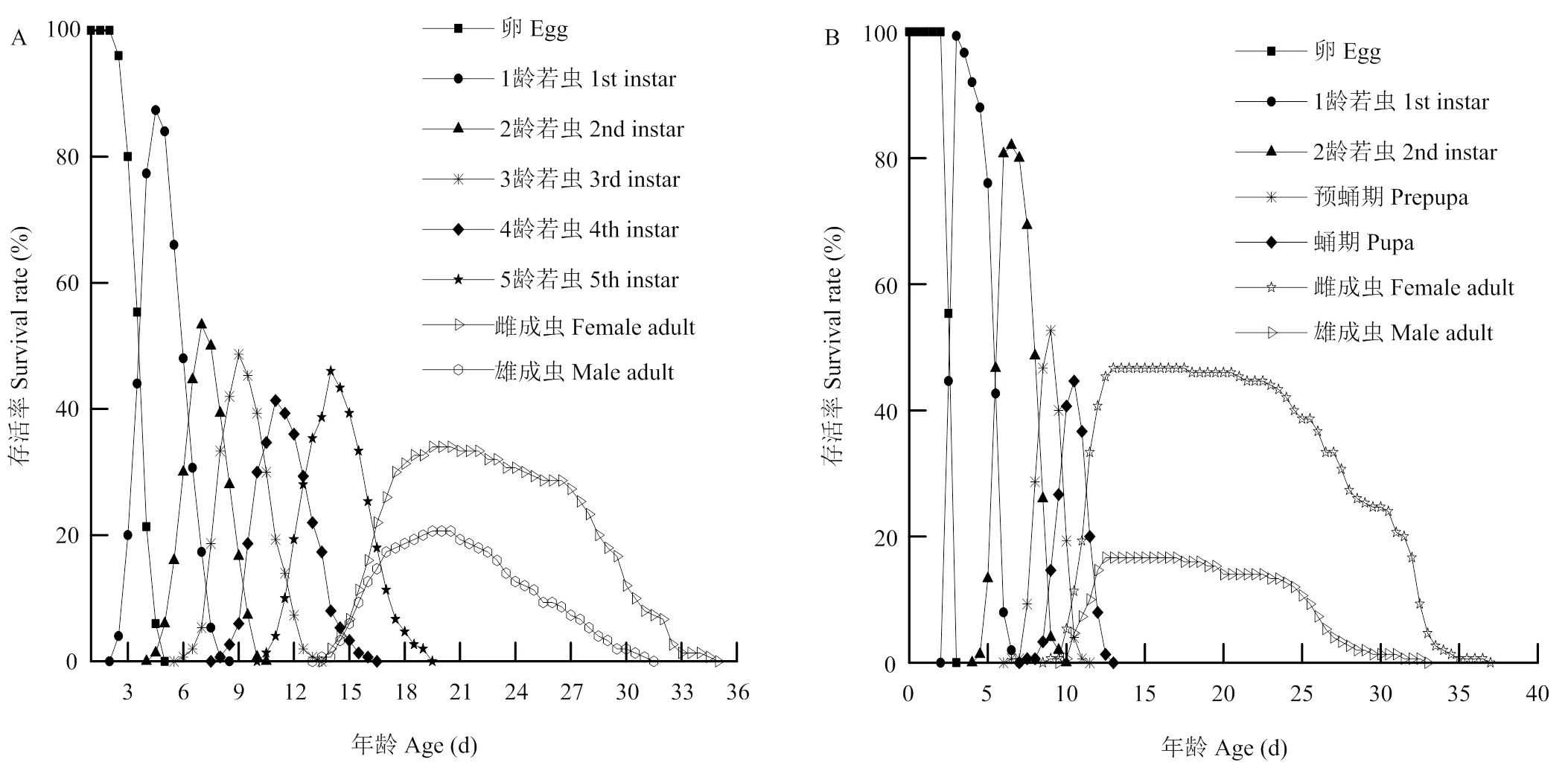
图1 南方小花蝽(A)和西花蓟马(B)的特定年龄-阶段存活率(Sxj)
2.3 南方小花蝽和西花蓟马的繁殖力
两种昆虫的繁殖力参数存在明显差异,其中南方小花蝽的雌成虫寿命、平均产卵期、平均产卵量和性比分别低于西花蓟马5.34 d、5.22 d、17.86粒、1.20。而产卵前期和总产卵前期南方小花蝽所经历的时间明显高于西花蓟马,分别高0.77 d和5.79 d(表2)。
特定年龄-存活率曲线(x)可以反映南方小花蝽取食西花蓟马,以及西花蓟马取食辣椒后从出生至死亡的变化情况(图2)。两个种群的存活率呈现出在若虫阶段其存活率不断下降,南方小花蝽在整个发育阶段的21.0 d后开始下降,西花蓟马为18.0 d,此时两种昆虫均为成虫阶段。南方小花蝽发育为成虫时的累计存活率为54.67%,西花蓟马发育为成虫阶段的累计存活率为63.33%。南方小花蝽在35 d时当代全部死亡,西花蓟马为37 d时当代全部死亡。特定年龄-阶段繁殖力(x)和特定年龄繁殖力(x)能反映两种昆虫从开始产卵到死亡时间段内不同发育阶段和年龄的繁殖情况,其单位为个体在0.5 d内繁殖的平均值。南方小花蝽和西花蓟马的繁殖参数x和x均表现出先升高后下降的趋势,其中南方小花蝽的x在整个发育时间的22 d时达到最大值2.66,西花蓟马为16 d时达到最大值2.87,此时间为两种昆虫的产卵高峰期(图2)。

表2 南方小花蝽和西花蓟马的繁殖力
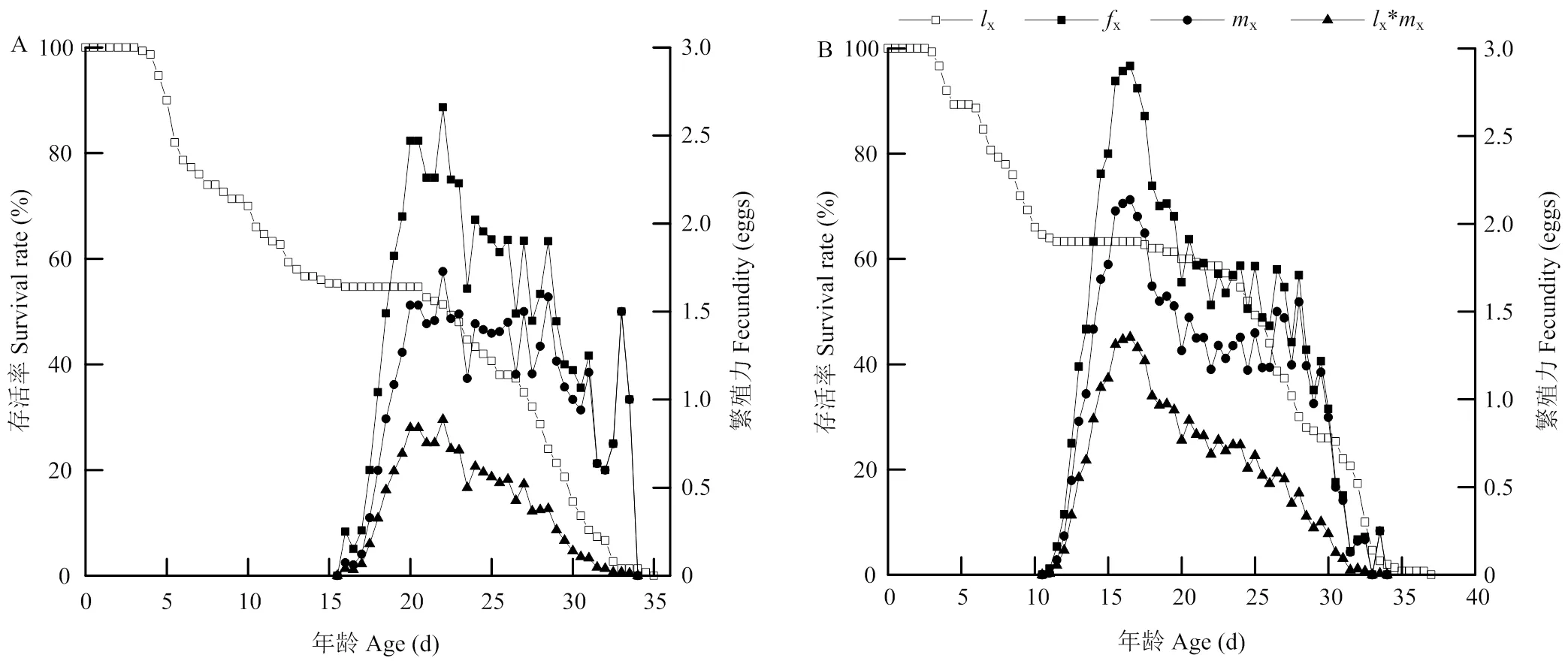
图2 南方小花蝽(A)和西花蓟马(B)的特定年龄存活率(lx)、特定年龄-阶段繁殖力(fx)和特定年龄繁殖力(mx)
2.4 南方小花蝽和西花蓟马的种群参数
南方小花蝽和西花蓟种群参数结果显示出两种昆虫具有不同种群增长潜力(表3)。西花蓟马的净生殖率、总繁殖率、内禀增长率和周限增长率均高于南方小花蝽,对应指数分别高出13.65、13.24、0.06、0.07。西花蓟马平均世代周期和种群加倍时间则短于南方小花蝽,分别少了4.31 d和2.12 d。
2.5 南方小花蝽和西花蓟马的期望寿命
南方小花蝽和西花蓟马的特定年龄-阶段寿命期望值xj结果显示(图3),南方小花蝽的1龄若虫期、西花蓟马的2龄若虫期和预蛹期的寿命期望值先随个体生长逐渐降低,在中期后又表现出增加的趋势。两种昆虫的雌、雄成虫寿命期望值在生长时间轴上总体表现为随着时间增加逐渐下降,并且两种昆虫均表现出雌虫的寿命期望值明显高于雄虫,西花蓟马的雌、雄成虫后期的寿命期望值接近,并且出现一段短暂的平缓变化。西花蓟马的总体生命期望高于南方小花蝽,尤其表现在成虫阶段。
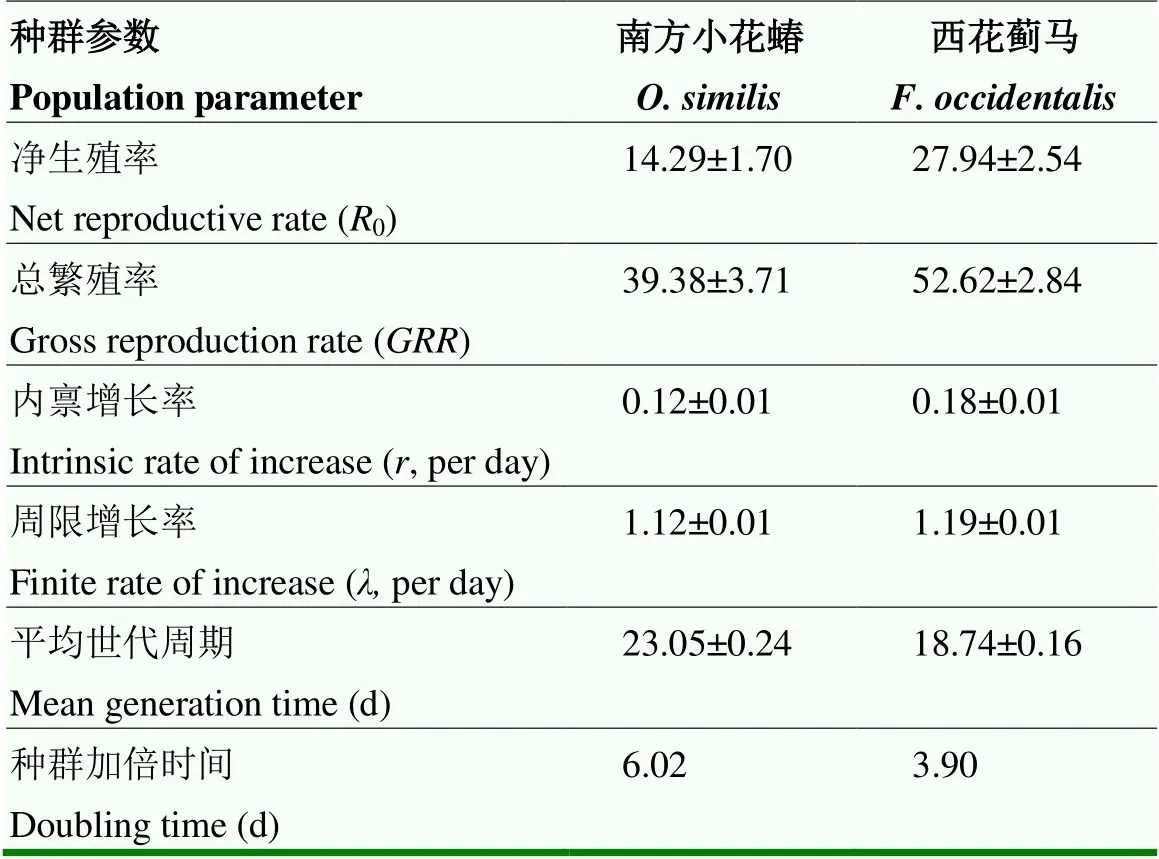
表3 南方小花蝽和西花蓟马的种群参数
2.6 南方小花蝽和西花蓟马的种群数量增长预测
以10粒南方小花蝽或西花蓟马卵为基数,南方小花蝽在90 d时可以繁殖成为768.08个后代,其中卵491.11粒,1—5龄若虫分别为167.30、32.60、6.64、1.47、6.33头,雌、雄成虫分别为41.28和21.33头。西花蓟马在90 d时可以繁殖为7 419.06个个体,其中卵3 259.77粒,1龄若虫至蛹分别为1 507.45、953.45、394.85、358.54头,雌、雄成虫分别为707.94和237.05头。西花蓟马的种群数量为南方小花蝽种群数量的9.66倍。其中作为主要繁殖个体的西花蓟马雌成虫为南方小花蝽的雌成虫的17.15倍。南方小花蝽在以10粒卵为基数经过40—60 d时达到第一个种群高峰,西花蓟马在30—50 d时达到第一种群高峰,南方小花蝽种群第一次高峰时间比西花蓟马晚10—20 d(图4)。
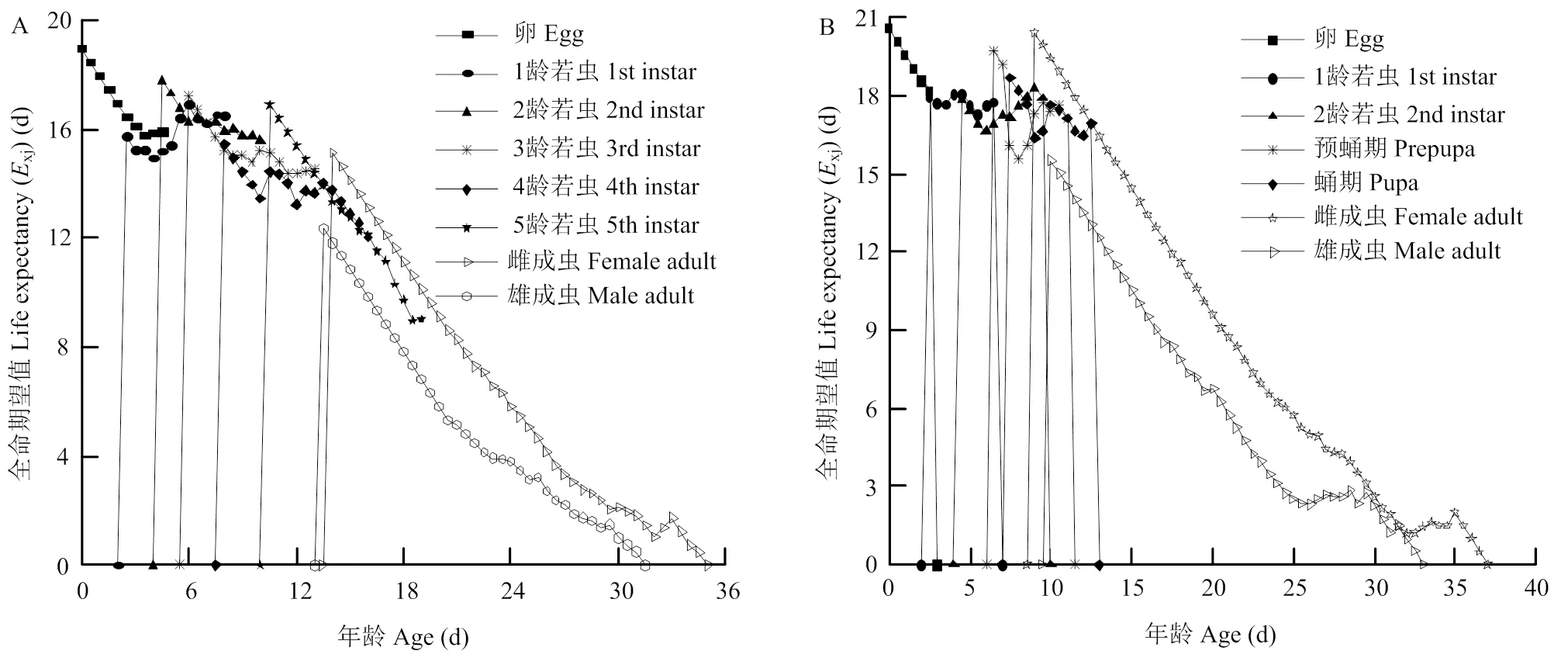
图3 南方小花蝽(A)和西花蓟马(B)的特定年龄-阶段寿命期望值(Exj)
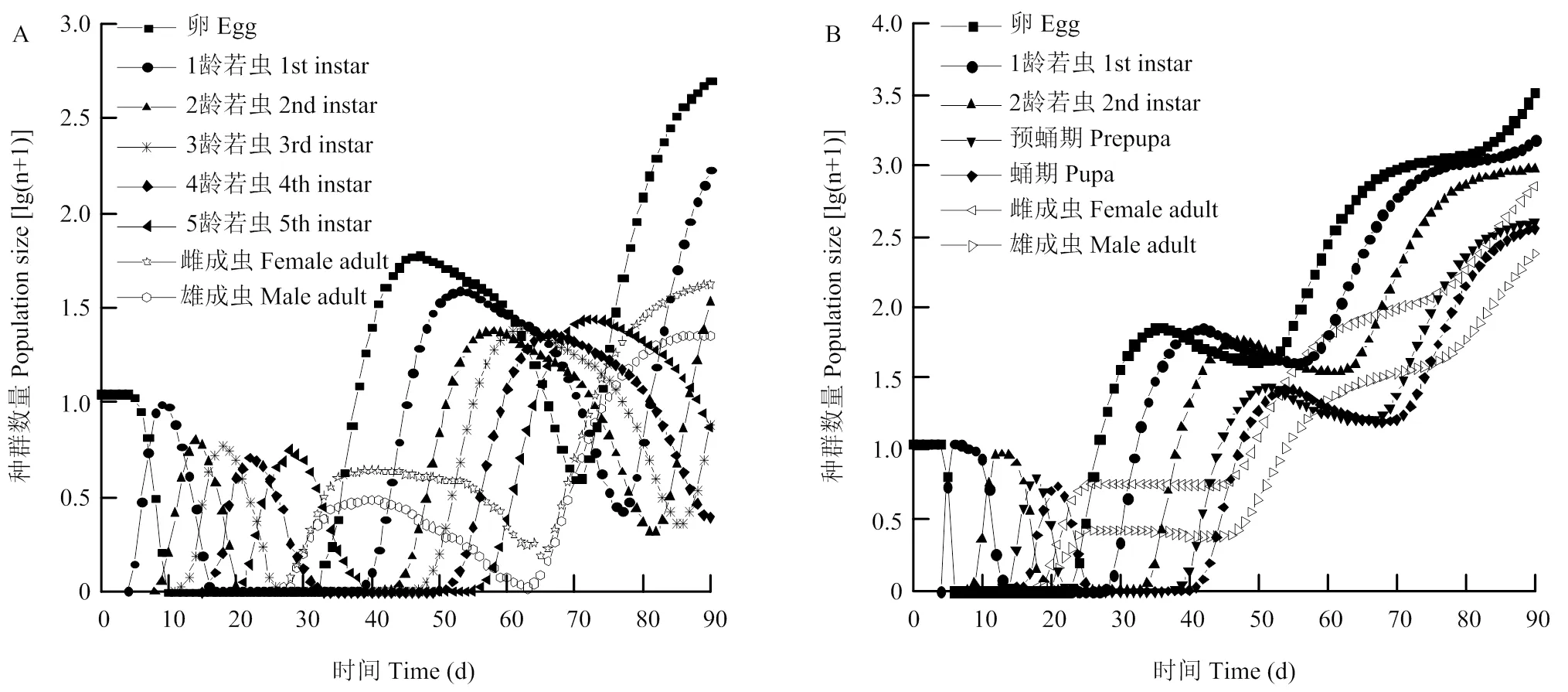
图4 模拟南方小花蝽(A)和西花蓟马(B)在无限制条件下的种群增长
2.7 南方小花蝽全龄期对西花蓟马2龄若虫的捕食量
不同龄期的南方小花蝽对西花蓟马2龄若虫的捕食量随着其龄期增加不断增大(表4),若虫完成一个发育阶段的捕食量为4.86头(1龄)至34.78头(5龄),雌成虫在整个存活期间对西花蓟马2龄若虫的捕食量平均为159.67头,明显高于雄成虫的86.00头。由捕食参数0和p可得,南方小花蝽在整个生命周期对西花蓟马2龄若虫的种群特征净捕食率为140.81头;取食西花蓟马2龄若虫的转化率为9.05头,即南方小花蝽产1粒卵需要消耗9.05头蓟马若虫(表4)。
南方小花蝽对西花蓟马2龄若虫的单日捕食量在1—3龄期时变化较小(图5),每天捕食的蓟马若虫平均分别为1.68、2.83和6.32头。而4—5龄若虫和雌、雄成虫的日均捕食量表现出先升高后下降的趋势,其日均取食量分别为8.82、10.43、12.13和8.90头(表4、图5)。捕食西花蓟马2龄若虫后南方小花蝽特定年龄存活率(x)随着寿命增加不断下降,特定年龄捕食率(x)和特定年龄净捕食率(x)结果表现出先升高后下降的趋势,其在成虫阶段的捕食率最大(图5)。其x的最大值为20 d时的14.06,南方小花蝽在19—24 d时的净捕食率较高,此时间段内为南方小花蝽的产卵高峰(图2、图6)。

表4 南方小花蝽对西花蓟马的捕食量及捕食参数
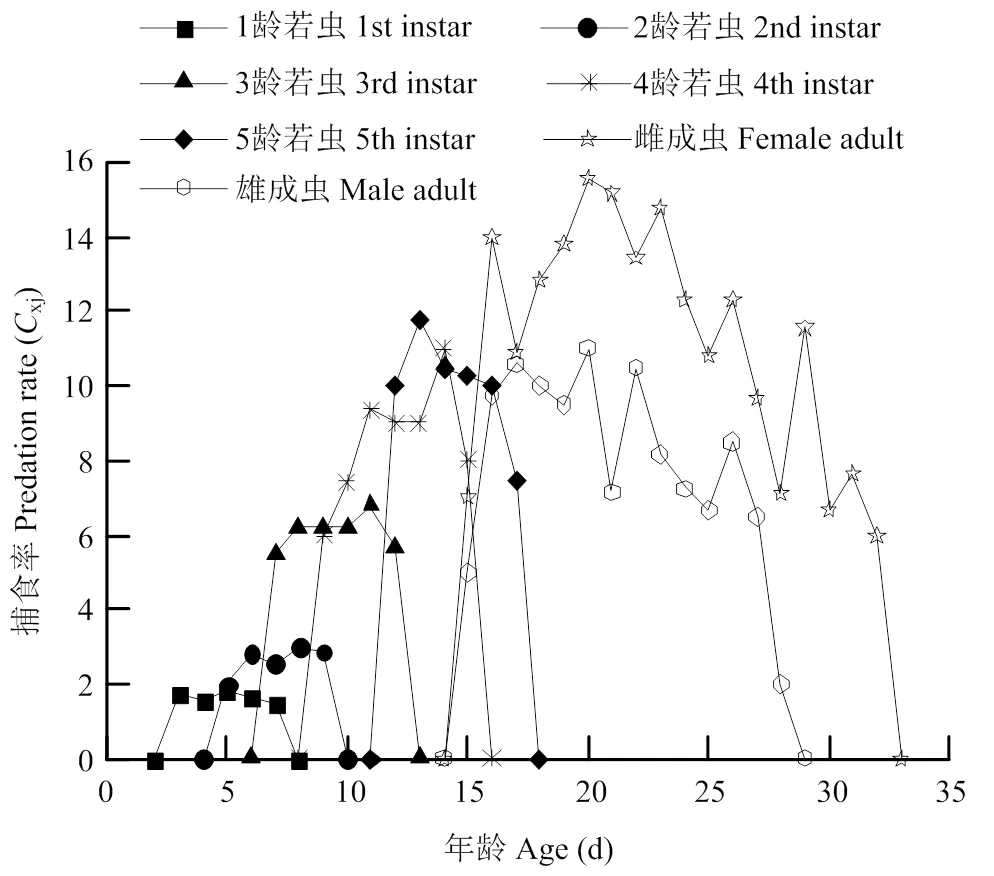
图5 捕食西花蓟马2龄若虫后南方小花蝽特定年龄-阶段捕食率(Cxj)
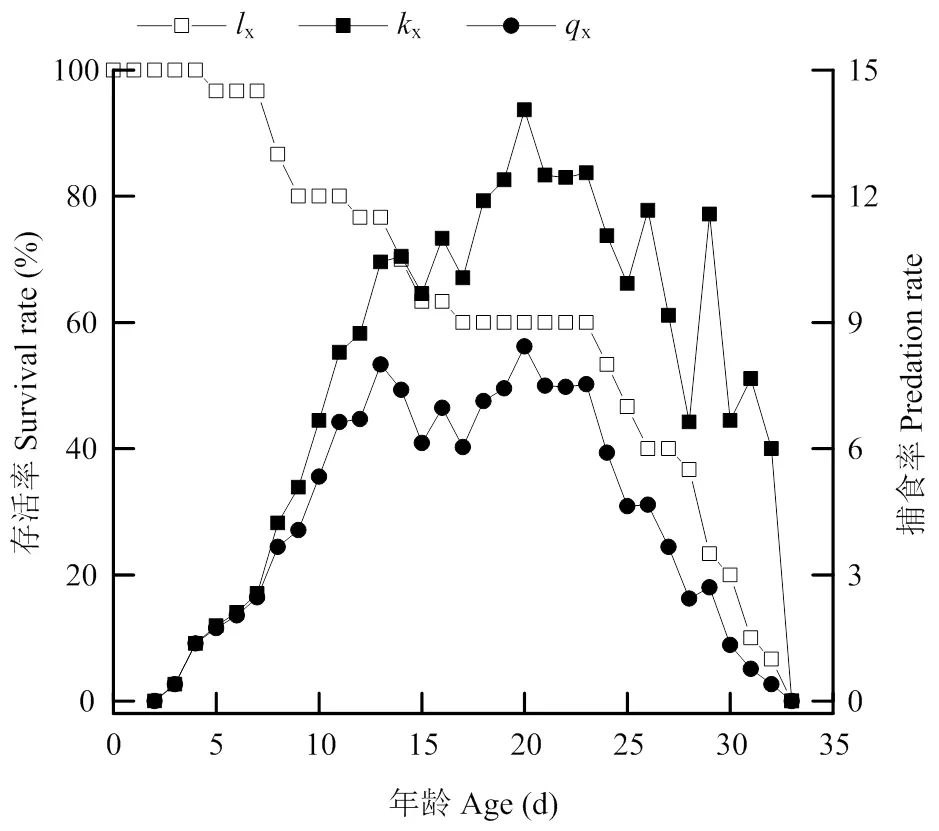
图6 捕食西花蓟马2龄若虫后南方小花蝽特定年龄存活率(lx)、特定年龄捕食率(kx)和特定年龄净捕食率(qx)
3 讨论
昆虫的生命表对于研究昆虫在不同环境中的发育情况、不同阶段的存活和繁殖能力差异以及种间关系探索和利用具有重要意义,昆虫生命表在预测害虫发生、探究昆虫的种群动态等方面起着至关重要的作用[18]。以不同龄期的生存状态构建南方小花蝽和西花蓟马的特定年龄-阶段存活率(xj)和两种昆虫特定年龄存活率(x)可以客观的描述两个种群从卵到成虫死亡的全过程,能更好地反映出天敌和害虫在种内和种间的发育速率和存活差异[18]。本研究围绕对我国农业生产具有重要危害的入侵害虫西花蓟马和南方地区主要的蓟马天敌南方小花蝽,构建了西花蓟马取食辣椒、南方小花蝽取食西花蓟马后两种昆虫的种群两性生命表。结果表明两种昆虫在各自的不同虫期均有较为明显的发育阶段重叠现象,不同发育阶段的存活率不同,同一发育阶段内的存活率均呈现出先升高后下降的变化趋势,且两种昆虫的雌成虫存活率均高于雄成虫。可能由于昆虫雌成虫的抗逆性强于雄性,如研究发现西花蓟马和花蓟马在杀虫剂胁迫下雌性比例升高,且雌成虫对杀虫剂的抗性高于雄性[4,20],这可能是两种昆虫适应寄主的一种机制,需要进一步探究确定。
昆虫的成虫阶段主要完成种群的繁殖,昆虫能否成功发育为成虫以及成虫阶段的寿命长短对种群增长具有重要意义。本研究表明,与西花蓟马相比,南方小花蝽需要更长时间完成若虫期的发育。在同一环境条件下,昆虫成虫阶段对于杀虫剂胁迫、温度胁迫等环境的抗逆性一般要远高于若虫[25-26]。南方小花蝽和西花蓟马相比,西花蓟马在各阶段的存活率更高,并且成虫阶段的存活率明显高于南方小花蝽。此外,在成虫期,南方小花蝽的雌、雄成虫寿命均短于西花蓟马,使得西花蓟马有更多的繁殖时间[27]。两种昆虫的雌成虫在产卵后期存活率不断下降,相似的结果在对加州新小绥螨()取食朱砂叶螨的研究中也有发现,其研究发现雌成螨随产卵期结束存活率下降,这一结果可能与雄成虫完成交配作用降低,雌成虫在大量产卵后消耗其自身能量有关[28]。
净生殖率(0)、总生殖率()、内禀增长率()、周限增长率()、平均世代周期()和种群加倍时间()6个参数作为昆虫种群预测中的重要指标,能反映出昆虫在相同环境中的种群增长潜力[20]。本研究测定了南方小花蝽和西花蓟马雌成虫的繁殖力和种群参数,南方小花蝽平均产卵量为42.00粒,低于西花蓟马的平均产卵量59.86粒,且雌雄性比也低于西花蓟马,此外,两种昆虫均在发育为成虫后4—6 d时达到产卵的高峰时期,但西花蓟马具有更长的产卵期,表明南方小花蝽的繁殖潜力明显低于西花蓟马。本研究对南方小花蝽和西花蓟马种群参数测定结果显示,天敌南方小花蝽的0、、、均低于西花蓟马,而和高于西花蓟马,表明西花蓟马具有更高的种群增长潜力。但本研究结果表明,南方小花蝽在取食西花蓟马后昆明地区的种群参数值低于贵州地区种群[29],而高于南方小花蝽取食豌豆蚜后贵州地区的种群参数[30],说明地理位置、猎物种类以及生存环境的差异均能对南方小花蝽的生长发育和繁殖情况产生较大影响。蒋春先等[28]研究了加州新小绥螨四川种群的发育情况,同样表明不同地理种群间的种群参数值存在较大差别。
基于种群参数,种群预测模型可以模拟昆虫在无限制条件下的种群增长情况[22]。对南方小花蝽和西花蓟马的种群数量预测结果显示,以相同的10粒卵为基数经过90 d后两种昆虫的种群数量明显不同,虽然都表现出较好的个体发育情况和种群增长潜力,但西花蓟马的个体数量达到南方小花蝽9.66倍,其中雌成虫数达到17.15倍,这可能与西花蓟马生物学特征(繁殖能力强)在种群增长中较南方小花蝽贡献更高有关。通常,当有害生物种群达到经济阈值时,生物防治过程中需要释放大量的天敌进行控制,为尽快达到控制效果,确定天敌的最佳释放量和释放时期至关重要,但随着释放量的增加,防治成本逐渐增加,因此,掌握好害虫的种群增长情况,提前进行预测能有效增加防治效果和降低防治成本[31-32]。南方小花蝽种群第一次高峰时间比西花蓟马晚10—20 d,在田间释放防治蓟马时应选择在蓟马密度较低,即种群暴发前10—20 d时进行首次释放,并根据蓟马的发生情况适当增加释放次数,以达到持续控制效果。
年龄-阶段捕食率可以准确描述捕食率随年龄和不同阶段的变化情况,全面评估天敌的生物防治潜力,在进行大规模的饲养和利用天敌昆虫之前,需要明确天敌不同阶段的捕食情况,才能更好地把控生物防治投入和产出之间的平衡点[24,33]。目前有关南方小花蝽捕食能力的报道中,大多以捕食功能反应来评价其捕食能力,且主要关注高龄若虫、成虫对西花蓟马的捕食作用,而未见基于生命表来评价其捕食能力[17,34]。本研究发现南方小花蝽1—3龄若虫的捕食率变化较小,而4—5龄若虫和雌成虫均存在捕食高峰。南方小花蝽雌成虫的捕食率随年龄增长而呈现波动,且其捕食率的波动幅度高于雄成虫。这与雌成虫在产卵阶段可能需要更多猎物来保证其营养供应有关[28]。并且其捕食高峰与产卵高峰时间点接近,在产卵后期雌成虫捕食量不断下降,可能由于产卵后需要营养降低而导致捕食率降低。本研究中南方小花蝽的捕食结果与东亚小花蝽()、无毛小花蝽()等捕食西花蓟马的捕食率变化相似[35-36],但不同小花蝽种在捕食量和龄期间存在一定差异,可能由于不同小花蝽的生物学差异导致。
4 结论
以西花蓟马2龄若虫为猎物时,南方小花蝽具有较好的个体发育、种群增长以及捕食能力。西花蓟马若虫期短、雌成虫寿命长、不同阶段的死亡率低于南方小花蝽,种群增长中有较高的繁殖力、内禀增长率以及较短的种群平均世代周期。西花蓟马种群与南方小花蝽种群相比在相同情况下依然具有更高的增长潜力,南方小花蝽在控制西花蓟马时存在一定局限性,其控制能力会在西花蓟马的种群密度过高时受到影响。此外,在田间释放南方小花蝽防治蓟马时,可根据蓟马种群在寄主植物上的动态变化选择蓟马种群暴发前10—20 d时进行持续释放,以达到更好的控制效果。
致谢:感谢台湾中兴大学齐心教授在本文的软件运用、文献查阅和理论知识补充中给予的帮助。
[1] GAO Y, LEI Z, REITZ S R. Western flower thrips resistance to insecticides: detection, mechanisms and management strategies. Pest Management Science, 2012, 68(8): 1111-1121.
[2] 张晓明, 姚茹瑜, 张宏瑞, 桂富荣, 李正跃. 不同花色菊花品种上西花蓟马种群密度及雌雄性比. 植物保护学报, 2017, 44(5): 737-745.
ZHANG X M, YAO R Y, ZHANG H R, GUI F R, LI Z Y. Population density and sex ratio of western flower thripson chrysanthemum cultivars of different flower colors. Journal of Plant Protection, 2017, 44(5): 737-745. (in Chinese)
[3] ZHANG B, QIAN W, QIAO X, XI Y, WAN F. Invasion biology, ecology, and management ofin China. Archives of insect biochemistry and physiology, 2019, 102(3): e21613.
[4] HE Z, GUO J F, REITZ S R, LEI Z R, WU S Y. A global invasion by the thrip,: Current virus vector status and its management. Insect science, 2020, 27(4): 626-645.
[5] 张晓明, 胡昌雄, 赵浩旭, 张宏瑞, 桂富荣, 李正跃. 吡虫啉胁迫对西花蓟马生长发育及种群性比的影响. 环境昆虫学报, 2017, 39(4): 870-878.
ZHANG X M, HU C X, ZHAO H X, ZHANG H R, GUI F R, LI Z Y. Effects of imidacloprid stress on development and sex ratio ofpopulations. Journal of Environmental Entomology, 2017, 39(4): 870-878. (in Chinese)
[6] GILBERTSON R L, BATUMAN O, WEBSTER C G, ADKINS S. Role of the insect supervectorsandin the emergence and global spread of plant viruses. Annual Review of Virology, 2015, 2(1): 67-93.
[7] 米娜. 两种蓟马趋光规律及不同波长色板田间诱捕效果评价[D]. 北京: 中国农业科学院, 2019.
MI N. The phototaxis behavior and effectiveness evaluation of sticky card trap for two thrips under different wavelengths[D]. Beijing: Chinese Academy of Agricultural Sciences, 2019. (in Chinese)
[8] WANG Z H, GONG Y J, JIN G H, LI B Y, CHEN J C, KANG Z J, ZHU L, GAO Y L, REITZ S, WEI S J. Field-evolved resistance to insecticides in the invasive western flower thrips(Pergande) (Thysanoptera: Thripidae) in China. Pest management science, 2016, 72(7): 1440-1444.
[9] 张晓明, 柳青, 李宜儒, 胡昌雄, 赵浩旭, 陈国华, 张宏瑞, 李正跃. 六种常见杀虫剂对西花蓟马和花蓟马的毒力测定. 环境昆虫学报, 2018, 40(1): 215-223.
ZHANG X M, LIU Q, LI Y R, HU C X, ZHAO H X, CHEN G H, ZHANG H R, LI Z Y. Toxicity of six current commonly used pesticides onand. Journal of Environmental Entomology, 2018, 40(1): 215-223. (in Chinese)
[10] WU S, TANG L, ZHANG X, XING Z, LEI Z, GAO Y. A decade of a thrips invasion in China: lessons learned. Ecotoxicology, 2018, 27(7): 1032-1038.
[11] SAITO T, BROWNBRIDGE M. Compatibility of soil-dwelling predators and microbial agents and their efficacy in controlling soil-dwelling stages of western flower thrips. Biological Control, 2016, 92: 92-100.
[12] 杨真. 云南省烟草蓟马种类及其天敌种类研究[D]. 昆明: 云南农业大学, 2016.
YANG Z. Study on thrips species and its natural enemies on tobacco in Yunnan Province[D]. Kunming: Yunnan Agricultural University, 2016. (in Chinese)
[13] 张昌容,郅军锐. 阶段性组合猎物饲养南方小花蝽的效果评价. 中国生物防治学报, 2017, 33(3): 345-350.
ZHANG C R, ZHI J R. Efficacy of stage-specific combined preys for rearing. Chinese Journal of Biological Control, 2017, 33(3): 345-350. (in Chinese)
[14] 周兴苗. 南方小花蝽大量繁殖关键技术及其田间释放生态学基础研究[D]. 武汉: 华中农业大学, 2007.
ZHOU X M. Key technology of mass rearing ofand its ecological basement of field release[D]. Wuhan: Huazhong Agricultural University, 2007. (in Chinese)
[15] 张士昶. 南方小花蝽人工规模化饲养技术的研究[D]. 武汉: 华中农业大学, 2009.
ZHANG S C. The study of artificial mass rearing technology ofZheng[D]. Wuhan: Huazhong Agricultural University, 2009. (in Chinese)
[16] ZHANG L M, LIU Z C, SUN X Q, LIU L X, CHEN J. Population dynamics ofandon Chinese rose and predatory functional response. Chinese Journal of Biological Control, 2008, 24(Suppl.): 21-27.
[17] 郅军锐, 郑珊珊, 张昌容, 刘丰姣. 南方小花蝽对西花蓟马和蚕豆蚜的捕食作用. 应用昆虫学报, 2011, 48(3): 573-578.
ZHI J R, ZHENG S S, ZHANG C R, LIU F J. The predation oftoand. Chinese Journal of Applied Entomology, 2011, 48(3): 573-578. (in Chinese)
[18] CHI H, YOU M S, ATLIHAN R, SMITH C L, KAVOUSI A, ÖZGÖKÇE M S, GÜNCAN A, TUAN S J, FU J W, XU Y Y,. Age-stage, two-sex life table: an introduction to theory, data analysis, and application. Entomologia Generalis, 2020, 40(2): 103-124.
[19] CHI H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environmental Entomology, 1998, 17(1): 26-34.
[20] 胡昌雄, 李宜儒, 李正跃, 陈国华, 张宏瑞, 桂富荣, 张晓明. 吡虫啉对西花蓟马和花蓟马种间竞争及后代发育的影响. 生态学杂志, 2018, 37(2): 453-461.
HU C X, LI Y R, LI Z Y, CHEN G H, ZHANG H R, GUI F R, ZHANG X M. Effects of imidacloprid on interspecific competition betweenandand their progeny development. Chinese Journal of Ecology, 2018, 37(2): 453-461. (in Chinese)
[21] LIU P, JIA W, ZHENG X, ZHANG L, SANGBARAMOU R, TAN S, LIU Y, SHI W. Predation functional response and life table parameters of(Hemiptera: Anthocoridae) feeding on(Thysanoptera: Thripidae). Florida Entomologist, 2018, 101(2): 254-259.
[22] 齐心, 傅建炜, 尤民生. 年龄-龄期两性生命表及其在种群生态学与害虫综合治理中的应用. 昆虫学报, 2019, 62(2): 255-262.
CHI H, FU J W, YOU M S. Age-stage, two-sex life table and its application in population ecology and integrated pest management. Acta Entomologica Sinica, 2019, 62(2): 255-262. (in Chinese)
[23] TUAN S J, LEE C C, CHI H. Population and damage projection of(F.) on peanuts (L.) under different conditions using the age-stage, two-sex life table. Pest Management Science, 2014, 70(5): 805-813.
[24] XU H Y, YANG N W, CHI H, REN G D, WAN F H. Comparison of demographic fitness and biocontrol effectiveness of two parasitoids,and(Hymenoptera: Aphelinidae), against(Hemiptera: Aleyrodidae). Pest management science, 2018, 74(9): 2116-2124.
[25] JENSEN S E. Insecticide resistance in the western flower thrips,. Integrated Pest Management Reviews, 2000, 5(2): 131-146.
[26] 张治军, 张友军, 徐宝云, 朱国仁, 吕要斌, 吴青君. 不同类型杀虫剂对西花蓟马的室内毒力. 浙江农业科学, 2013(6): 694-697, 699.
Zhang Z J, Zhang Y J, XU B Y, Zhu G R, Lü Y B, Wu Q J. Laboratory toxicity of different insecticides to. Journal of Zhejiang Agricultural Sciences, 2013(6): 694-697, 699. (in Chinese)
[27] 章士美, 杨明旭. 昆虫的成虫期. 江西植保, 1985(1): 22-25.
ZHANG S M, YANG M X. Adult stage of insects. Jiangxi Plant Protection, 1985(1): 22-25. (in Chinese)
[28] 蒋春先, 税玲, 蒲颇, 李庆. 基于两性生命表的以朱砂叶螨为猎物的加州新小绥螨四川种群生长发育、繁殖及捕食率. 中国生物防治学报, 2019, 35(3): 364-373.
JIANG C X, SHUI L, PU P, LI Q. Population growth development, fecundity and predation rate of(McGregor) (Acari: Phytoseiidae) sichuan strain fed onBoisduval (Acari: Tetranychidae) based on two-sex life table. Chinese Journal of Biological Control, 2019, 35(3): 364-373. (in Chinese)
[29] 张昌容, 郅军锐, 莫利锋. 四种猎物对南方小花蝽生长发育和繁殖的影响. 生态学报, 2012, 32(18): 5646-5652.
ZHANG C R, ZHI J R, MO L F. The influence of 4 species of preys on the development and fecundity ofZheng. Acta Ecologica Sinica, 2012, 32(18): 5646-5652. (in Chinese)
[30] 郑珊珊, 郅军锐, 张昌容, 刘丰娇. 南方小花蝽的发育与繁殖研究. 河南农业科学, 2009(12): 88-91.
ZHENG S S, ZHI J R, ZHANG C R, LIU F J. Studies ondevelopment and reproduction with broad bean aphid as prey. Journal of Henan Agricultural Sciences, 2009(12): 88-91. (in Chinese)
[31] XU H Y, YANG N W, WAN F H. Field cage evaluation of interspecific interaction of two aphelinid parasitoids and biocontrol effect on(Hemiptera: Aleyrodidae) Middle East-Asia Minor 1. Entomological Science, 2015, 18(2): 237-244.
[32] YANG N W, WAN F H. Host suitability of different instars ofbiotype B for the parasitoid. Biological Control, 2011, 59(2): 313-317.
[33] FARHADI R, ALLAHYARI H, CHI H. Life table and predation capacity of(Coleoptera: Coccinellidae) feeding on(Hemiptera: Aphididae). Biological Control, 2011, 59(2): 83-89.
[34] 蒋兴川, 桂富荣, 陈斌, 李志华, 穆静娟, 李正跃. 南方小花蝽在不同试验空间对西花蓟马的捕食及搜寻效应. 生物安全学报, 2012, 21(1): 20-26.
JIANG X C, GUI F R, CHEN B, LI Z H, MU J J, LI Z Y. Prey preference and searching efficiency ofonin different experimental space. Journal of Biosafety, 2012, 21(1): 20-26. (in Chinese)
[35] COCUZZA G E, DE CLERCQ P, LIZZIO S, VAN DE VEIRE M, TIRRY L, DEGHEELE D, VACANTE V. Life tables and predation activity ofandat three constant temperatures. Entomologia experimentalis et applicata, 1997, 85(3): 189-198.
[36] WANG S, MICHAUD J P, TAN X L, ZHANG F. Comparative suitability of aphids, thrips and mites as prey for the flower bug(Hemiptera: Anthocoridae). European Journal of Entomology, 2014, 111(2): 221-226.
Control effect ofonbased on the two-sex life table and the age-stage-specific predation rate
HU ChangXiong1, FAN Wei1, ZHANG Qian1, CHEN GuoHua1, YIN HongHui2, XU TianYang2, YANG Jinbo3, YANG Hang3, WU Daohui1, ZHANG XiaoMing1
1College of Plant Protection, Yunnan Agricultural University/State Key Laboratory of Yunnan Biological Resources Protection and Utilization, Kunming 650201;2Wenshan Branch of Yunnan Tobacco Company, Wenshan 663000, Yunnan;3Yunnan Gerui Biological Technology Co., Ltd., Midu 675602, Yunnan
【】is an important agricultural invasive pest, andis the dominant predator species in China. The objective of this study is to systematically evaluate the control effect ofonand the biological differences between this two species in the same area.【】andwere collected from open field, then reared by fresh shoots with flowers offor 2-3 generations under indoor condition, and the 2nd instar nymphs ofwere feed toin the experiment. Growth, reproductive rate, predation rate were tested in Yunnan populations ofandusing the two-sex life table and the age-stage-specific predation rate methods.【】The different insect periods ofandhave different levels of overlap during their developmental stages, and the survival rate of female adults of both insect species was higher than that of male adults.nymph has 5 instars, whilehas 4 instars. Compared with,has a longer nymph duration and shorter adult lifespan, and the survival rate of each developmental period was lower than that of, especially the survival rate of female adults was obviously lower than that of. The probability ofsuccessfully developing from egg to female adult and male adult was 32.67% and 20.67%, respectively, which was lower than that of’s 46.67% and 16.67%.has a higher proportion of females within its population. The reproductive parameters (x) ofandpeaked at 22 d and 16 d during the whole developmental stages, respectively.has an average egg production of 42.00 eggs per female, which was significantly lower than that of the(59.86 eggs). The population net reproduction rate (0), gross reproduction rate (), intrinsic rate of increase () and finite rate of increase () inwere lower than those of, while the mean generation time () and population doubling time () inwere longer than those of. The population prediction results showed that the individuals ofreached 9.66 times than that ofafter 90 days of population growth and development, and the number of female adults reached 17.15 times. The predation rate ofon the 2nd instar nymphs ofgradually increases with age, and the population characteristic net predation rate (0) ofwas 140.81, and the transformation rate (p) was 9.05. The average predation of female and male adultto2nd instar nymphs was 159.67 and 86.00 individuals, respectively.【】population can finish its life history and has stable population growth when fed by.has a stronger population growth potential than that ofunder the same conditions. As an important natural enemy ofbiological control,should be released 10-20 d beforeoutbreak and continuous released duringoutbreak, this can achieve better control effects to.
;; two-sex life table; predation rate; biological control

10.3864/j.issn.0578-1752.2021.13.007
2020-09-04;
2020-10-13
云南省基础研究专项面上项目(202001AT070134)、云南省中青年学术技术带头人后备人才项目(202105AC160071)、“云南省高层次人才培养支持计划”青年拔尖人才项目(云人社通〔2020〕150-09号)、中国烟草总公司云南省公司科技计划(2018530000241015)、云南省教育厅科学研究基金(2019Y0089)
胡昌雄,E-mail:hcxbear@126.com。通信作者张晓明,E-mail:zxmalex@126.com
(责任编辑 岳梅)
猜你喜欢
昆虫学报(2022年9期)2022-10-18
农业工程学报(2022年10期)2022-08-22
昆虫学报(2022年6期)2022-07-13
天津农业科学(2022年5期)2022-05-31
今日农业(2021年11期)2021-11-27
建材发展导向(2021年11期)2021-07-28
南方农业学报(2021年11期)2021-03-05
祝您健康(2020年5期)2020-05-14
江苏农业科学(2017年12期)2017-09-16
新农村(2015年9期)2015-09-24
