
头季稻氮肥运筹对再生稻根际机能及产量的影响
2021-07-19 08:51:56黄锦文吴珈谊陈鸿飞张志兴方长旬邵彩虹林伟伟翁佩莹林文雄
中国水稻科学 2021年4期
黄锦文 吴珈谊 陈鸿飞 张志兴 方长旬 邵彩虹 林伟伟 翁佩莹 林文雄,*
头季稻氮肥运筹对再生稻根际机能及产量的影响
黄锦文1, #吴珈谊1, #陈鸿飞1张志兴1方长旬1邵彩虹2林伟伟1翁佩莹1林文雄1,*
(1作物遗传育种与综合利用教育部重点实验室/福建农林大学农学院/作物生态与分子生理学福建省高校重点实验室, 福州 350002;2江西省农业科学院土壤肥料与资源环境研究所, 南昌 330200;#共同第一作者;*通信联系人,E-mail: wenxiong181@163.com)
【目的】再生稻是头季稻生长的延续。本研究尝试从根际微生态系统的变化来阐明机械化栽培下头季稻氮肥管理对头季-再生季水稻产量形成的影响,为再生稻高产高效栽培提供参考。【方法】以“甬优1540”为材料,通过2年的田间试验,在头季总施氮量(225.00 kg/hm2)不变的前提下,设置头季不同氮肥运筹处理,分析了机械化栽培下再生稻产量、根系伤流强度、根际土壤酶活性及其微生物多样性的变化特点。【结果】头季前氮后移施肥处理(N1、N2)有助于根系保持较高活力,尤其在头季生长中后期N1处理(基肥∶一次分蘖肥∶二次分蘖肥∶孕穗肥=3∶1∶2∶4)根系伤流强度比对照(N0)和当地常规施肥处理(N4)显著提高;N1处理两季总产量最高,2年平均达17 351.23 kg/hm2,比当地常规施肥处理(N4)提高了23.00%。土壤酶活性分析表明,蔗糖酶活性在头季齐穗期表现为N2>N1>N3>N4>N0,之后则表现为N1>N2>N3>N4>N0;过氧化氢酶与硝酸还原酶活性在头季生长期均表现为N1>N2>N3>N4>N0;脲酶活性从头季齐穗期至再生季齐穗始终表现为N1>N2>N3>N4>N0;各生育时期土壤酶活性与根系伤流强度间均呈显著或极显著正相关。细菌 16S rDNA 测序表明,施肥与不施肥处理细菌群落结构多样性差异显著,N1处理细菌 Chao1 指数和香农指数显著高于其他处理。菌群结构分析表明,施肥处理增加了与土壤碳氮循环、有机质含量及抗逆性密切相关的细菌数量,尤其是N1、N2处理硝化螺旋菌属相对丰度较高,有利土壤氮素循环。【结论】机械化栽培下,头季氮肥适当后移有利于再生稻根际微生态系统的改善,从而提高根系活力,促进水稻生长,获得两季水稻的高产。
再生稻;氮肥运筹;根系活力;根际土壤;群落多样性
近年来,强再生力水稻品种的育成和栽培技术的发展,使得再生稻栽培成为中国南方提高复种指数,稳定稻谷总产的一项种植制度[1]。再生稻是头季水稻收割后稻桩上的休眠芽萌发而成的一季短生育期水稻,其根系由头季根系和再生根系两部分组成[2]。研究表明,再生苗成穗率及产量与再生稻的根系机能极显著正相关,健壮的头季稻根系和一定数量新生根系的有效结合是再生稻高产的关键[3-6]。氮素是水稻生长发育进程以及产量形成的关键养分因子。饶鸣钿等[7]研究指出,头季稻氮肥是影响再生季产量的主要因素之一。陈鸿飞等[8]、杨东等[9-10]、王森等[11]和郑景生等[12]从水稻光合生理、氮代谢、干物质积累运转及养分的吸收利用等方面分析了头季稻不同氮肥运筹方式对再生稻产量形成的影响;任天举等[13]、林燕等[14]认为头季稻不同的施肥方式对再生稻的生育进程、根系和再生苗生长具有明显的效应。前人的研究表明,合理的氮肥运筹不仅对提高头季产量,而且对防止头季稻后期根系早衰,稳定再生季产量,实现两季高产意义重大[15]。然而,前人的研究大多是在人工栽培条件下展开的。随着农村劳动力的转移,农业生产机械化已成为当前作物生产的主流。与人工栽培相比,机械化栽培再生稻必须解决两个关键问题:一是头季机插秧移栽秧龄小,生育期短,植伤重[16-19];其次,与人工高留桩栽培不同,机插再生稻必须选用耐碾压品种进行低留桩收割来减轻收割机对稻桩的碾压[1]。可见,机插再生稻的品种选择及相应的栽培管理模式都与人工栽培有所不同。目前,机栽下头季稻氮肥管理方式对再生稻根际机能及产量的影响未见深入的研究报道。此外,近年来研究认为,根系-微生物的直接相互作用对植物的生长、健康和适应性起着至关重要的作用,根际被认为是植物的第二基因组[20]。植物与根际微生物的互作现象及其机制,也是生命科学研究关注的热点和农业微生物利用的关键问题[21,22]。因此,本研究以闽北广泛种植的“甬优1540”为材料,并基于人工栽培水稻农民多施基肥的传统习惯和机栽再生稻生长的特点,设置了头季稻氮肥适当后移的施肥模式,分析头季稻氮肥运筹对再生稻根际微生物群落多样性的影响及其对土壤酶活性和根系活力的影响,尝试从微生态系统变化来阐明机栽再生稻高产形成机理。
1 材料与方法
1.1 试验材料与设计
试验以籼型三系杂交稻“甬优1540”为材料,于2017年3月至11月在福建省南平市建阳区崇雒乡(118°13′E, 27°30′N)进行。试验地为沙壤土,含有机质3.4%,铵态氮20.5 mg/kg,速效磷6.8 mg/kg,速效钾65 mg/kg,pH值为6.3。2017年头季于3月11日播种,采用软盘育秧机插,4月11日插秧,插秧规格30 cm×17 cm,丛插平均2粒谷秧。头季稻施氮量为(折合纯氮)225 kg/hm2,N∶P2O5∶K2O=1∶0.5∶0.8,以尿素作氮肥,以过磷酸钙作P肥,P肥全部作基肥,以氯化钾作K肥,K肥基肥∶孕穗肥= 5∶5,氮肥运筹比例设5种处理,即基肥、一次分蘖肥、二次分蘖肥、孕穗肥比分别为3∶1∶2∶4、3∶2∶1∶4、3∶3∶0∶4、4∶3∶0∶3,依次设为N1、N2、N3、N4处理,以头季及再生季均不施氮肥为对照(N0处理),其中,N4是当地常规施肥方式,具体氮肥施用见表1。每处理种植750 m2,设置3次重复。各处理间设置塑料挡板以防止串肥,设置高30 cm、宽40 cm田埂,工作沟宽30 cm;头季成熟时机收,留桩高度为25 cm。再生季施肥情况:除N0处理外,其他各处理施纯氮172.5 kg/hm2,以尿素作氮肥,按1∶1分促芽肥和促苗肥两次施用,促芽肥于头季齐穗后20 d施用,促苗肥于头季收割后5 d施用。再生稻全程机械化栽培,除施肥处理外,其他按当地常规高产栽培管理。2018年在同一地块上进行重复试验,头季于3月4日播种,4月10日插秧,其他试验设计与2017年相同。2017年进行头季和再生季水稻测产,2018年除产量测定外,还进行了根系伤流量及根际土壤酶活性测定和土壤细菌群落特性分析。
B-头季稻基肥,插秧前2 d施用;T1-第一次分蘖肥,插秧10 d后施用;T2-第二次分蘖肥,第一次分蘖肥施用后10 d施用;RB-孕穗肥,倒2叶期施用。BP-促芽肥,头季稻齐穗后20 d施用;SP-再生季促苗肥,头季收割后5 d施用。
B refers to basal fertilizer of main crop rice, which was applied two days before transplanting; T1, Primary fertilizer application for tillering, which was applied 10 d after transplanting; T2, Secondary fertilizer application for tillering, which was applied 10 d after T1;RB stands for topdressing for panicle initiation, which was applied in the penultimate leaf growth period.BP refers to bud-promoting fertilizer, which was applied 20 d after full heading; SP refers to seedling-promoting fertilizer, which was applied 5 d after harvest.
1.2 测定项目与方法
1.2.1 水稻产量与产量构成因素测定
于头季及再生季水稻成熟期,每个小区根据平均有效穗数取有代表性水稻 5 株,考查每株有效穗数、每穗总粒数及实粒数、千粒重,计算结实率和理论产量并记录。水稻成熟期每处理随机抽取3块有代表性的田块人工收割测产,每块地实割测产面积2 m×2 m。
1.2.2 水稻根系伤流量测定
分别于头季水稻齐穗期、成熟期和再生季水稻的齐穗期、成熟期,采用李合生[23]贴棉吸附法进行根系伤流量测定。每个小区于测定日18:00选取具有代表性的3丛水稻,每丛水稻在离地12 cm处水平割去地上部分,并将已知质量的棉花完全包裹住切口,套上对应的塑料袋,用橡皮筋进行固定,做好防雨水、露水措施;次日6:00回收田间放置的棉花袋,并进行称量,记录其增量。每个处理3次重复。
1.2.3 土壤样品采集
于头季水稻齐穗期、成熟期和再生季水稻齐穗期、成熟期取样,测定土壤相关酶活性;于头季水稻生长后期(齐穗后10 d)取样,分析根际土壤细菌群落特性。
每次每处理采用五点取样法随机选取稻株(连根拔起),用无菌剪刀剥除离根系较远的土壤,露出根际土,用剪刀轻轻剥掉根上大约1~3mm的土壤混匀,分为2份,分别保存于4℃冰箱和−80℃冰箱备用。
1.2.4 根际土壤相关酶活测定
参照关松荫等[24]的方法,根际土壤中脲酶采用苯酚钠-次氯酸钠比色法测定,蔗糖酶采用3,5-二硝基水杨酸比色法测定,过氧化氢酶采用高锰酸钾滴定法测定;硝酸还原酶参照Alef 等[25]的方法采用高锰酸钾滴定法测定。每个小区取3次重复。
1.2.5 根际土壤微生物总 DNA提取及PCR扩增
土壤DNA 提取方法参照 FastDNA SPIN Kit for soil(MP Biomedicals,USA)试剂盒说明书进行。提取得到的DNA样品用1%琼脂糖凝胶电泳检测和分光光度法(260 nm/280 nm光密度比)进行质量检测。检测后,将样品于−20℃下保存以备后续实验使用。微生物多样性检测选取细菌16S rRNA V3~V4区,DNA样本送至北京奥维森基因科技有限公司,利用 Illumina Miseq PE300高通量测序平台进行测序。细菌16S rRNA V3~V4可变区特异引物为338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACNNGGGTATCTAAT-3')。PCR反应体系包括12.5 μL 2×PCR MasterMix,3 μL BSA (2 ng/μL),2 Primer(5 μmol/L),2 μL模板 DNA和5.5 μL ddH2O。反应程序:95 ℃下预变性5 min;95 ℃下变性45 s,55 ℃下退火50 s,72 ℃下延伸45 s,32个循环;72 ℃下延伸10 min。测序原始序列上传至NCBI的SRA数据库。
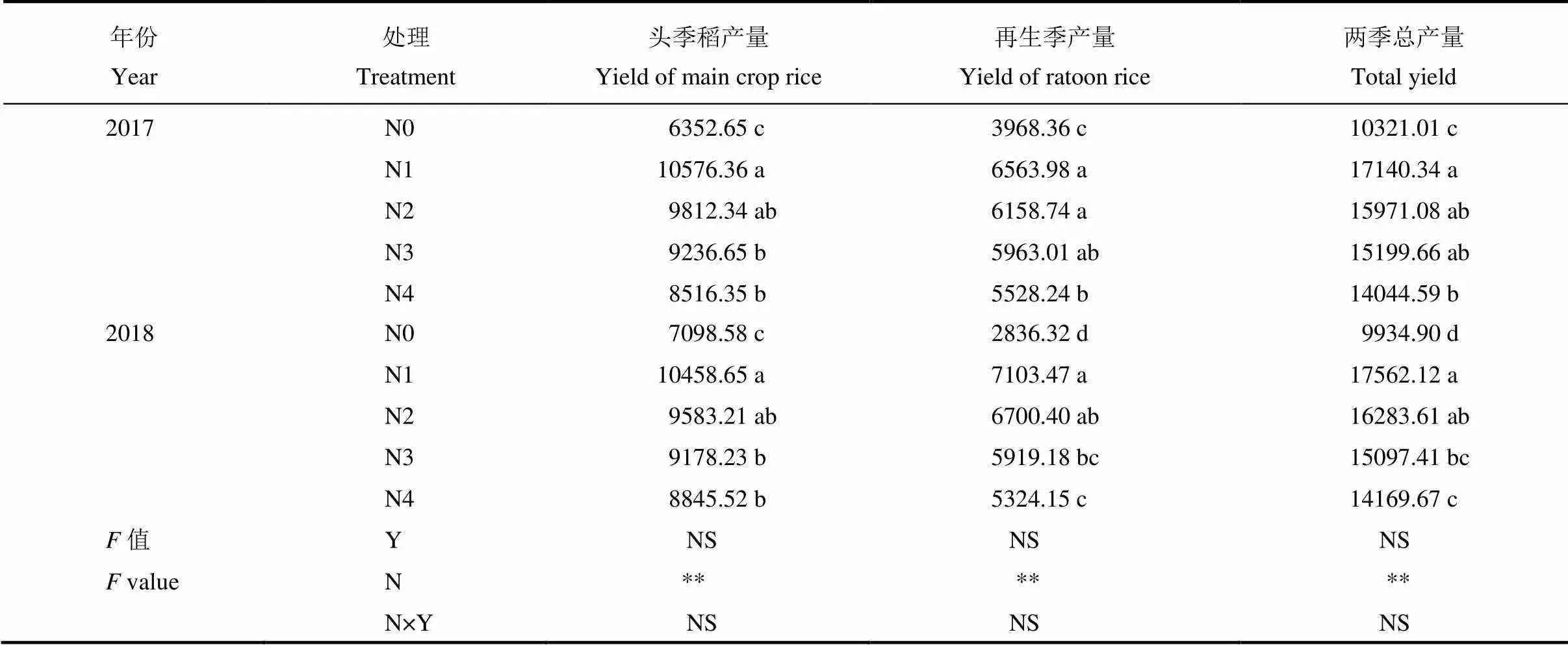
表2 头季稻氮肥运筹对头季-再生季水稻产量的影响
*<0.05; **<0.01; *Significant at< 0.05; **Significant at<0.01.
1.3 数据处理与分析
测序数据分析:原始数据存在一定干扰数据,在去除barcode、primer序列后使用FLASH 软件进行拼接,得到原始Tags 数据(raw_tags),对Tags数据进行过滤,得到高质量Tags数据(clean_tags)。使用usearch软件按照97%相似性序列进行OTU (Operational Taxonomic Unit,可操作分类单元)聚类,获得OTU,统计所测样品每个OTU中的丰度信息。随机抽样法抽取数据,并与所代表的OTU数目构建稀释曲线,使用R语言和Mothur绘制稀释曲线并进行Alpha多样性指数组间差异分析。使用 Qiime 软件计算所测样品Observed-species、Chao1、Shannon、Goods-coverage、PD_whole_tree 指数。对OTU代表序列进行数据库比对分析,得出物种注释总表,并进行不同水平的物种数量统计,获得各分类水平(Phylum, Class, Order, Family, Genus, Species)的物种丰度表,进行属水平OTU丰度微生物功能分析。对OTU丰度进行均一化处理,并且进行Alpha多样性和PCA 统计分析。使用R语言作图。
水稻产量、根系伤流量及酶活性测定结果采用Excel 2016和DPS 7.05统计软件进行整理分析,采用GraphPad Prism 5进行数据图表处理。
2 结果与分析
2.1 头季-再生季水稻产量及产量构成因素
由表2可知,产量年度间差异不显著,施肥处理间差异极显著。2年的产量表现无论是头季还是再生季,施氮处理均极显著高于不施氮(N0)处理,头季及再生季产量均表现为N1>N2>N3>N4>N0,其中N1与N2之间差异不显著。N1两季实际总产量2年平均比不施肥N0、当地常规施肥N4处理分别提高71.32%、23.00%;N2两季实际总产量2年平均比N0、N4处理分别提高59.24%、14.32%。
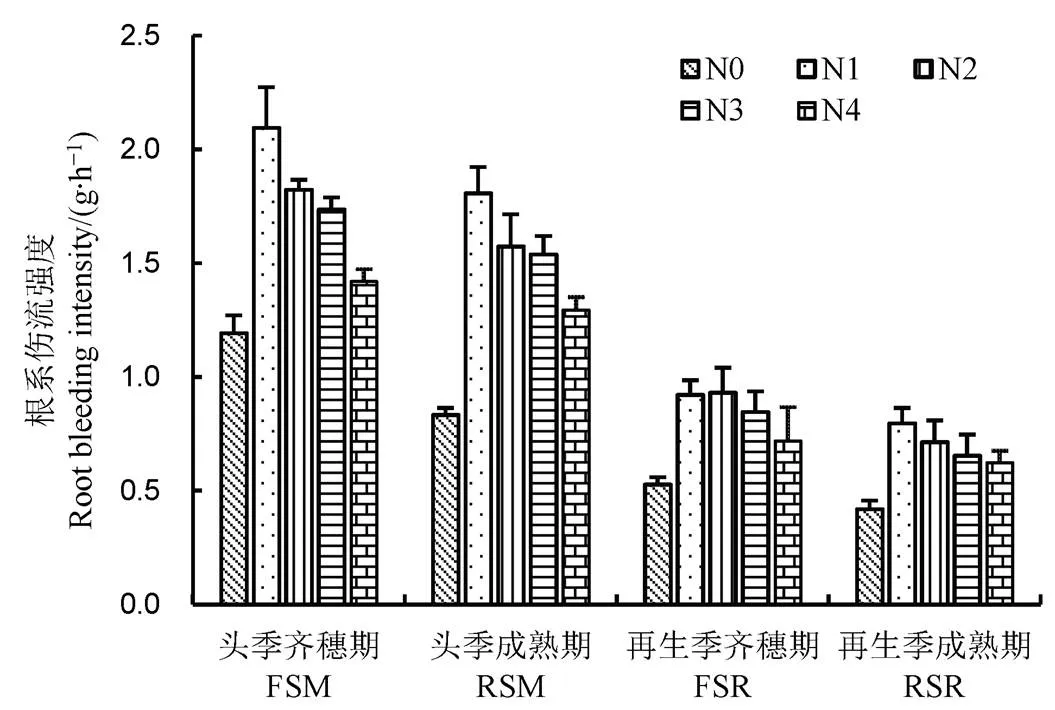
图1 水稻根系伤流强度随生育时期变化
Fig.1.Changes of root bleeding intensity of rice at various growth stages.
由于年度间产量差异不显著,以2年产量平均值作进一步分析(表3),各处理再生季有效穗数均比头季稻多,尤其是施肥处理,再生季平均有效穗数是头季稻的1.27~1.39倍,其中,N1处理不管是头季还是再生季有效穗数均最高,其次是N2处理。每穗粒数方面,再生季比头季少,各处理再生季为头季稻的52.23%~55.24%,头季及再生季各处理间每穗粒数表现为N2>N1>N3>N4>N0,再生季各施肥处理间差异不显著。千粒重不管头季还是再生季各处理间无显著差异;头季结实率平均为93.45%,再生季平均为80.23%,头季高于再生季。
2.2 水稻根系活力及其与再生季产量的相关性
2.2.1 根系伤流强度的变化
根系伤流是不挖掘水稻的情况下能对水稻根系生理活性进行简单判断的有效方法,其流量强度是根系活力的指标之一。由图1可知,从头季齐穗-头季成熟-再生季齐穗-再生季成熟随着生育进程各处理根系伤流强度呈下降趋势,头季齐穗期各处理均达到峰值,N1、N2处理显著高于N3、N4、N0处理,N1处理每株水稻根系伤流强度达2.10 g/h,比N0处理高75.65%,比N4处理高47.89%;N2处理比N0处理高52.94%,比N4处理高28.17%,N2与N3处理差异不显著。头季成熟期各处理根系伤流强度均下降,但依然表现为N1>N2>N3>N4>N0,N1处理分别比N0、N4处理提高了116.88%和39.66%。随着再生稻生长发育进程,再生季水稻根系伤流强度都明显低于头季稻,由于再生季除对照外各处理施肥一致,各处理间(除对照外)差距也明显减小,但依然是N1、N2伤流强度较高。可见,在前氮后移N1、N2处理下水稻整个生育期中根系均能保持较高活力,尤其头季生长后期N1处理根系伤流强度显著高于其他处理。
2.2.2 根系伤流强度与再生季产量的相关性
根系伤流强度与再生季水稻产量及产量构成因素相关分析表明(表4),再生季产量与头季齐穗和成熟及再生季的齐穗与成熟4个时期的根系伤流强度均达到极显著正相关,说明从头季到再生季生长过程根系活力对再生季产量都极为重要,其中与头季成熟期相关度最高。进一步从产量构成因素分析,有效穗数与以上4个时期的根系伤流强度也均为极显著正相关,其中与头季成熟期根系伤流强度相关度最高;每穗粒数与再生季成熟期的根系伤流强度显著相关,但与头季齐穗期、头季成熟期及再生季齐穗期根系伤流量强度相关未达显著水平;其他产量构成因素与根系伤流强度之间相关不显著。由此可见,根系活力主要通过影响有效穗数从而影响再生季产量,且以头季齐穗至头季成熟期根系活力最为关键。
2.3 根际土壤酶活性变化及其根系活力的相关性
2.3.1 土壤酶活性变化
表5表明,水稻根际土壤与营养转化相关的蔗糖酶、过氧化氢酶、脲酶和硝酸还原酶活性,头季和再生季均表现为齐穗期高于成熟期,但不同土壤酶在不同生育时期间变化趋势并不完全一致。

表3 头季-再生季水稻产量及产量构成因素变化(2年平均值)
同一列中同一稻作类型各平均数之后不同小写字母表示差异达5%显著水平。下同。
Different letters indicate statistical significance at the 0.05 probability level within the same column for the same cropping system.
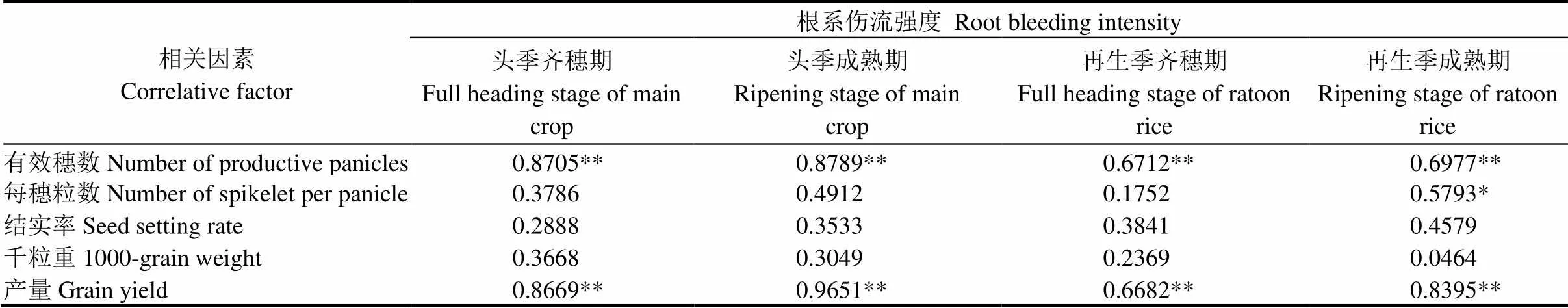
表4 水稻根系伤流强度与再生季产量及其构成因素的相关性
*和**分别表示=0.05和=0.01显著水平;=15,0.05=0.5139;0.01=0.6411。
* indicate significant correlation at the level of 0.05;**indicate extremely significant correlation at the level of 0.01.=15,0.05=0.5139;0.01=0.6411.
蔗糖酶活性总体表现为头季齐穗期最高,头季成熟期下降。再生季齐穗有较大幅度上升,至再生季成熟期又下降。从不同处理看,在头季齐穗期中蔗糖活性最高的为N2处理,比N0和N4分别高出171.12%和136.75%,其次为N1、N3、N4、N0处理,但其他3个时期均是N1处理活性最强,其次是N2处理。

图2 头季水稻生长后期根际细菌群落Shannon-Wiener曲线
Fig.2.Shannon-Wiener curves of bacterial community in rhizosphere soil at the late stage of main crop rice.
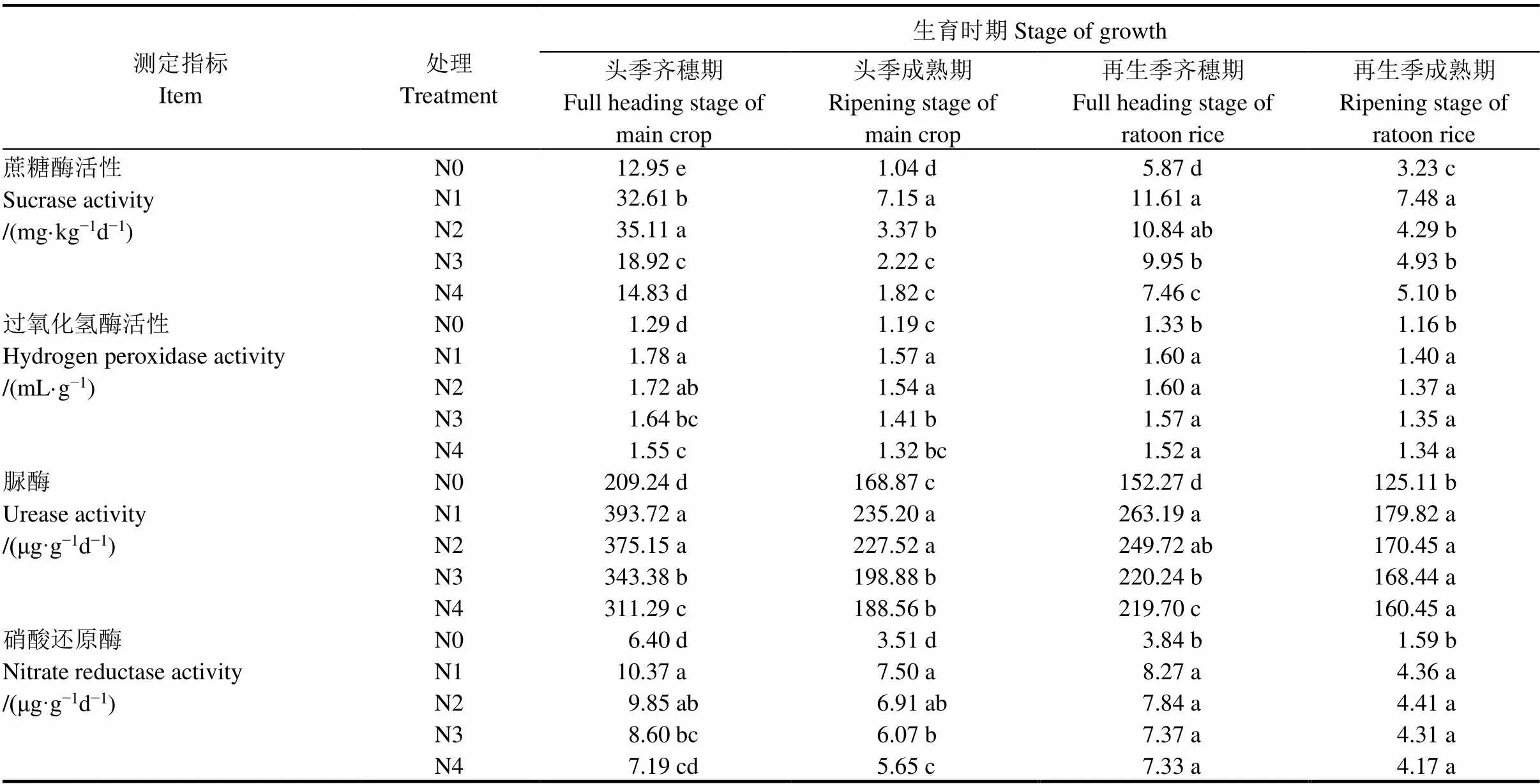
表5 根际土壤酶活性随水稻生育时期变化情况
过氧化氢酶各处理在头季齐穗期和成熟期均表现为前氮后移的N1、N2处理显著高于其他处理,但N1与N2处理间差异不显著,头季齐穗期N1、N2处理分别比N4处理(当地常规施肥)高出14.83%、10.97%,头季成熟期N1、N2处理分别比N4处理高出18.94%、17.42%。但到了再生季齐穗期和成熟期各施肥处理均显著高于不施肥对照,但各施肥处理间差异不显著。
脲酶广泛存在于土壤之中,与尿素氮肥水解密切相关,在一定程度上反映土壤的供氮能力。头季稻齐穗期、成熟期和再生季齐穗期各处理脲酶均表现为N1>N2>N3>N4>N0,但N1与N2处理间差异不显著,到了再生季成熟期各施肥处理间差异不显著,但各施肥处理均显著高于不施肥对照。
在头季稻生长期内,各处理硝酸还原酶活性差异大多显著,在头季稻齐穗期和成熟期均表现为N1>N2>N3>N4>N0,但N1与N2处理间差异不显著,N3与N2处理间差异不显著;到了再生季齐穗期和成熟期各施肥处理间差异不显著,但各施肥处理均显著高于不施肥对照。
2.3.2 根系伤流强度与土壤酶活性的相关性
不同生育时期根系伤流强度与土壤酶活性的相关性分析表明(表6):根系伤流强度与头季齐穗期至再生季成熟期各时期的蔗糖酶、过氧化氢酶、脲酶及硝酸还原酶酶活性均呈极显著或显著正相关。其中,与蔗糖酶活性的相关性不管是头季还是再生季其齐穗期相关性均大于成熟期;与土壤解毒相关的过氧化氢酶及与氮素营养相关的脲酶和硝酸还原酶活性的相关系数则表现为头季大于再生季。可见,头季稻籽粒灌浆过程,即头季齐穗期至头季成熟期土壤酶活性对根系生长的影响更突出。

表6 根系伤流强度与土壤酶活性的相关性分析
*和**分别表示=0.05和=0.01显著水平;=15,r0.05=0.5139; r0.01=0.6411。
* indicate significant correlation at the level of 0.05 level; **indicate extremely significant correlation at the level of 0.01.
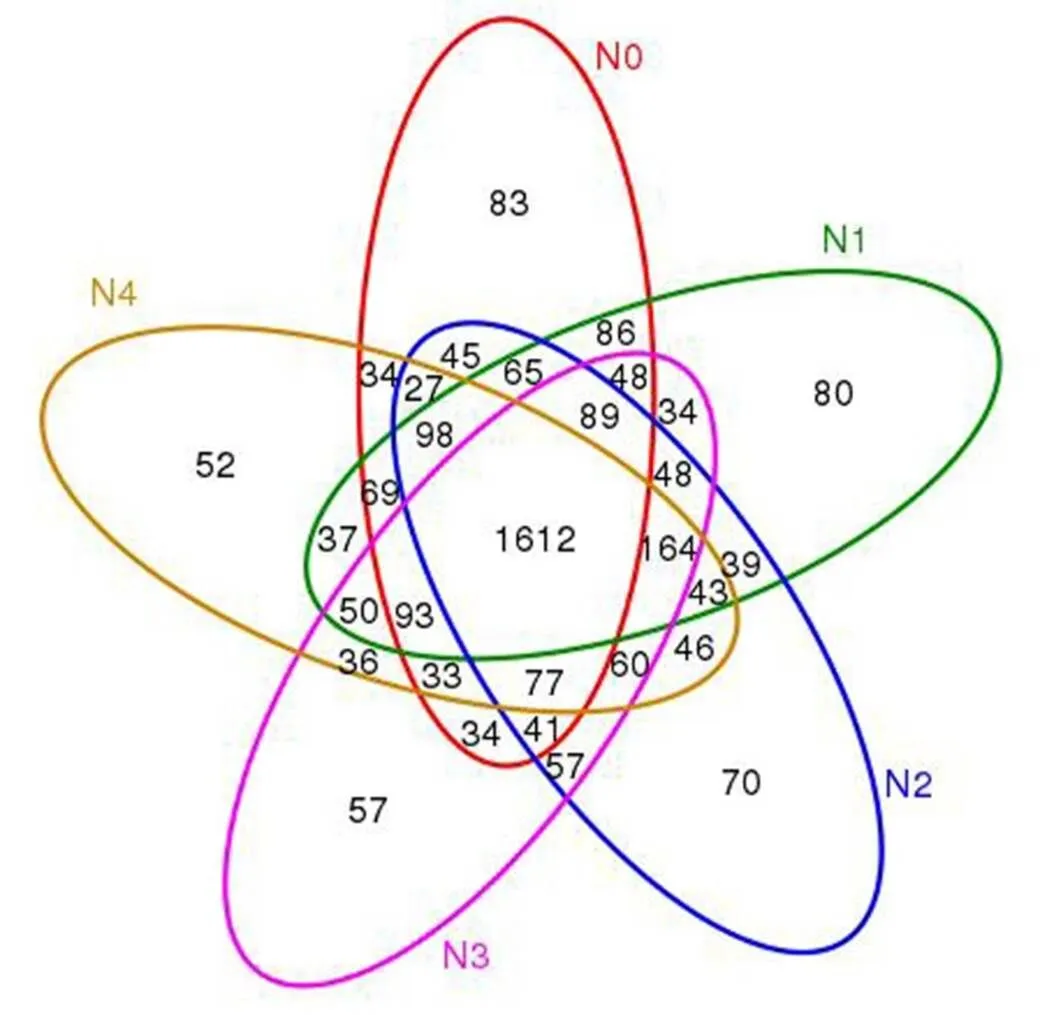
圆圈相交部分表示共有微生物数目,非相交部分表示特有微生物数目
Fig.3.Venn graph of bacterial OTUs in rhizosphere soil at the late stage of main rice.
2.4 根际土壤细菌群落的变化
头季水稻生长后期是决定其收割后稻桩上腋芽萌发好坏的关键时期,腋芽的萌发又与其根系生长好坏有着直接的关系,而土壤微生物参与土壤中的各种生物化学过程,是土壤养分转化、活化的主要动力,是影响根系生长的主要环境因素。
2.4.1 根际土壤细菌群落Alpha多样性分析
Shannon-Wiener曲线是采用97%相似度的 OTU 对所测定土壤样本进行稀释曲线分析,用以反映各样本在不同测序数量时的微生物多样性[26]。通过不同样本的测序量在不同测序深度水平上的微生物多样性指数来构建稀释性曲线,由于深层次测序数量只会产生少量新增的OTU,因此,当曲线趋向平坦时,说明测序数据量足够大,可以反映样品中绝大多数的微生物信息。从图2可以看出,随着测序深度的增加,各处理下的OTU数逐渐增长,随后趋于平坦,说明各处理测序数据量合理,所测根际土壤样本可以反映大多数细菌群落信息。observed_species指数是指随测序深度的增加,实测到 OTU 的数量,Coverage为样本文库覆盖度。表7表明,5个处理的observed_species和Coverage两个指数差异都不显著,且数值均处于合理范围,Coverage各样本测序覆盖度均大于0.95,样本测序结果已经包含样本中绝大部分的细菌物种,可以进行后续分析。
表7显示了头季不同氮肥管理下土壤根际细菌群落Alpha多样性指数。Chao1 指数反映样品中群落的丰富度,与样品中物种丰富性呈正相关。表7表明,所有施肥处理Chao1 指数均高于对照N0处理,N1处理其值最高,其次是N2处理,但N1与N2处理间差异不显著,N3、N4与N0间差异不显著;Shannon 指数反映群落的多样性,5个处理中N1处理其值最高,显著高于不施肥的N0处理和当地常规施肥N4处理,其他4个处理间差异不显著;PD_whole_tree指数反映了样品中物种对进化历史保存的差异。表7表明,N1处理PD_whole_tree指数值最高,N2次之,两者间无显著差异,但显著高于N0和N4处理。

表7 头季水稻生长后期根际土壤细菌群落多样性指数
韦恩图(Venn Graph)可以用来比较样本之间特有及共有的OTU (操作分类单元)数量。图3表明,在头季不同氮肥运筹下,鉴定到头季生长后期N0、N1、N2、N3、N4处理的根际土壤OTU数分别2534、2655、2581、2533、2531个。其中,N1处理所测定的OTU数量最多,达到2655个,其次是N2处理。5个处理共有OTU 1612个,N0、N1、N2、N3和N4处理特有的OTU数分别为83、80、72、57、52个,说明不施肥处理下存在特有的OTU数量最多,而在头季同等施肥量条件下,N1处理特有的OTU数高于其他施肥处理,N2次之,N3、N4处理特有的OTU数量则明显减少。
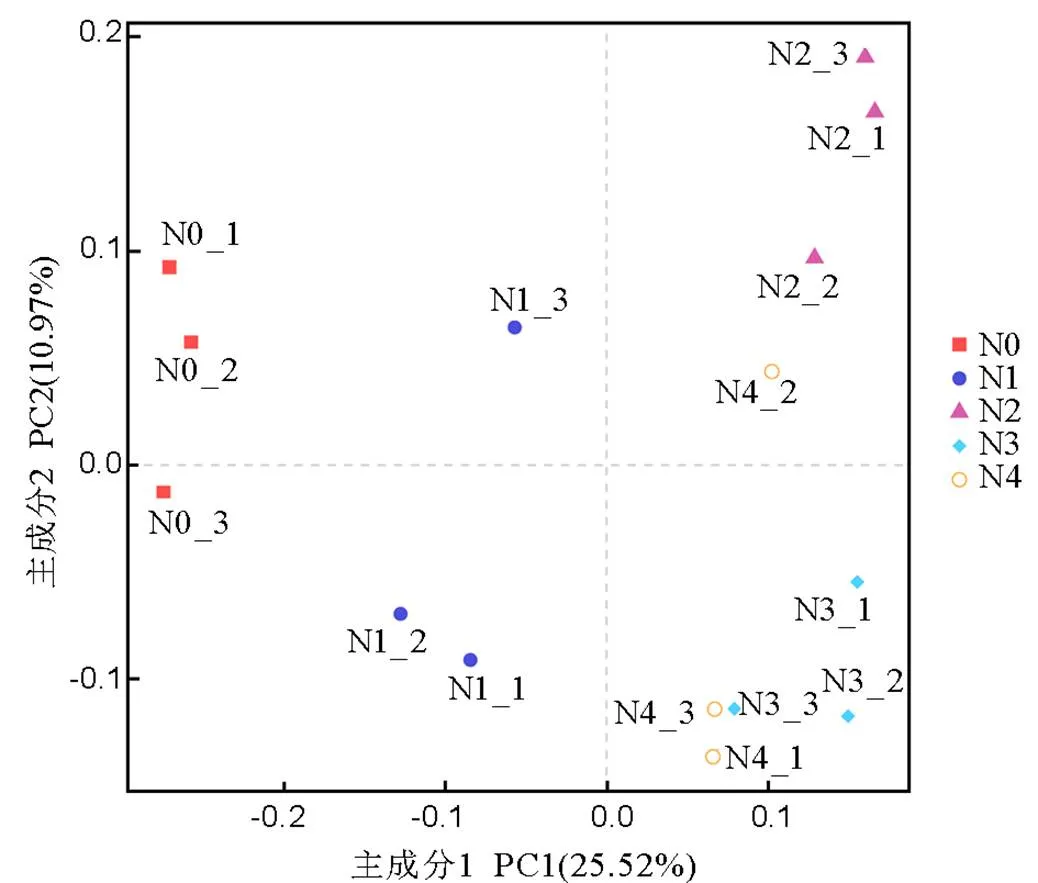
图4 头季稻生长后期根际土壤细菌群落PCA分析
Fig.4.PCA analysis of bacterial community in rhizosphere soil at the late stage of main crop rice.
2.4.2 根际土壤细菌群落PCA分析
PCA分析(Principal Component Analysis),即主成分分析,通过分析不同样本中97%相似性的OTU组成,可以初步反映出各处理样本间可能表现出分散和聚集的分布情况。如图4所示,主成分1和主成分2反映处理间差异的方差累计贡献率分别为25.52%和10.97%。总体来看,头季水稻施肥与不施肥处理引起的差异主要沿着横轴PC1分布,不施肥的N0处理位于主成分1的负端,远离各施肥处理,表明施肥处理与不施肥处理细菌群落差异性较大;N2、N3和N4处理位于主成分1的正端且区分不明显,N1处理与这3个处理有一定的距离,表明N1处理与N2、N3、N4之间细菌群落差异性也比较大。由施肥处理引起的差异主要沿着纵轴PC2分布,N3与N4处理间差异小主要位于PC2的负端,N2处理则位于PC2的正端,N1则位于二者之间。可见,主成分分析基本上能够区分不同施肥处理间根际土壤微生物的群落特征,尤其是施肥与不施肥处理间区分明显。
2.4.3 根际细菌群落构成分析
将优质序列聚类后得到的OTU进行物种注释,选取各分类水平上丰度最高的物种绘制优势物种的相对丰度柱形图。图5-A显示,细菌群落结构中相对丰度大于1%的门类中位于前10位的从大到小依次为酸杆菌门(Acidobacteria)、变形菌门(Proteo bacteria)、绿弯菌门(Chloroflexi)、硝化螺旋菌门(Nitrospirae)、芽单胞菌门(Gemmatimonadetes)、放线菌门(Actinobacteria)、疣微菌门(Verrucomi crobia)、Latescibaoteria、Lgnavibacteriae和拟杆菌门(Bacteroidetes),共占总数的93%以上。各处理均以酸杆菌门相对丰度最高,达24%~40%,其中,N4处理酸杆菌门相对丰度最高,N1处理最低;N1处理变形菌门和绿弯菌门相对丰度均最高,其他各处理间无显著差异;硝化螺旋菌门是一类革兰氏阴性细菌,N1处理相对丰度最高,其次是N0处理,其他各处理间差异不大。芽单胞菌门各施肥处理高于不施肥处理,但施肥处理间差异不显著。
图5-B表明,共鉴定到的细菌纲类有25个。其中根际相对丰度大于1%的纲类(top10)从大到小依次酸杆菌(Acidobacter)、变形菌(Deltaproteobac teria)、α-变形菌-革兰氏阴性菌(Alphaproteobact eria)、硝化螺旋菌门下的Niterospira、SBR2076,酸杆菌门下的Solibacteres、未被鉴定的纲类、β-变形菌纲(Betaproteobacteria)、Subgroup_18和纤线杆菌纲(Ktedonobacteria)。其中,N2、N3、N4处理中酸杆菌纲类、纤线杆菌纲相对丰度比较高;N0和N1处理中变形菌纲类、SBR2076相对丰度比较高;N1、N2处理中β-变形菌纲的相对丰度较高。
图5-C显示,共鉴定到的细菌属类10个,已鉴定的比重最大的细菌属是,属于念珠菌属中分解有机质、利用碳源的细菌。其他依次为地杆菌属()、、氨基酸杆菌属()、。其中N0、N1处理地杆菌属显著高于其他处理。与对照不施肥处理相比较,各施肥处理中氨基酸杆菌属相对丰度较高,该菌属主要功能有利于发酵土壤氨基酸,特别是谷氨酸。

A-门水平差异;B-纲水平差异;C-属水平差异。
Fig.5.Changes in the relative abundance of main bacteria in the rhizosphere soil at the late growth stage of main crop rice.
2.4.4 根际土壤细菌功能分析
如表8所示,相比不施肥(N0)处理,各施肥处理的根际土壤红假单胞菌属、硝化螺旋菌属、罗氏菌属、土壤杆菌属、酸杆菌菌属、黏细菌属微生物相对丰度均有所提升,而厌氧绳菌属、假双头斧形菌属和嘉利翁氏铁柄杆菌属的微生物相对丰度有所下降。其中,红假单胞菌属属于固氮细菌类型,而硝化螺旋菌属则参与地球氮循环的硝化作用,二者均与土壤氮素循环有关[27],各施肥处理红假单胞菌属丰度均比不施肥处理的有所提高,其中N3处理丰度最高,依次为N4、N2、N1处理;各施肥处理下硝化螺旋菌属的微生物丰度明显高于对照,尤其前氮后移的N1和N2处理要高于N3和N4处理,表明施肥尤其头季前氮后移有利头季生长后期根际土壤硝化螺旋菌属数量的增加,从而促进土壤氮循环。土壤杆菌属和酸杆菌菌属功能与改善土壤质地及增加土壤有机质有关[28],各施肥处理均显著高于不施肥处理,促进土壤化能有机营养,从而提高土壤有机质含量。黏细菌属微生物有利提高作物的抗逆性,各施肥处理黏细菌属微生物丰度均比不施肥显著提高,前期施氮较重的N3、N4处理比前氮后移施肥处理的N1、N2处理来得高。此外,厌氧绳菌属、假双头斧形菌属、嘉利翁氏铁柄杆菌属微生物相对丰度则表现为不施肥(N0)处理高于各施肥处理,说明不施肥处理促进土壤碳水化合物降解且土壤微生物耐药能力要强于施肥处理,不施肥处理中嘉利翁氏铁柄杆菌属的增加则会促进土壤中二价亚铁氧化成三价铁,浓缩和积累土壤环境中的铁元素[29]。
3 讨论
再生稻是头季稻收割后稻桩上休眠腋芽萌发生长抽穗再收获的一季水稻,因此,头季稻收割后休眠腋芽能否萌发成苗是再生稻栽培的关键。郑景生等[5]研究指出再生稻再生分蘖的生长发育依赖于头季稻残留的根系。在本研究中,头季齐穗期各处理根系伤流强度均达到峰值,其中N1(基肥∶一次分蘖肥∶二次分蘖肥∶孕穗肥=3∶1∶2∶4)处理最高,其次是N2处理,N1处理分别比N0和N4处理高75.65%、47.89%。进一步的相关分析表明,再生季水稻产量,尤其是产量构成中的有效穗数与各时期(头季齐穗至再生季成熟),特别是头季稻生长后期的根系活力呈极显著正相关(表4)。刘富贵等[30]研究也表明头季稻根系对再生季稻的生长发育起主导作用。可见,前氮后移的N1处理因为根系活力强,尤其头季后期根系不早衰不仅有利头季产量提高,也为后季再生苗分蘖成穗提供了保障,因而两季实际总产最高,比当地常规施肥N4处理提高了23.00%。
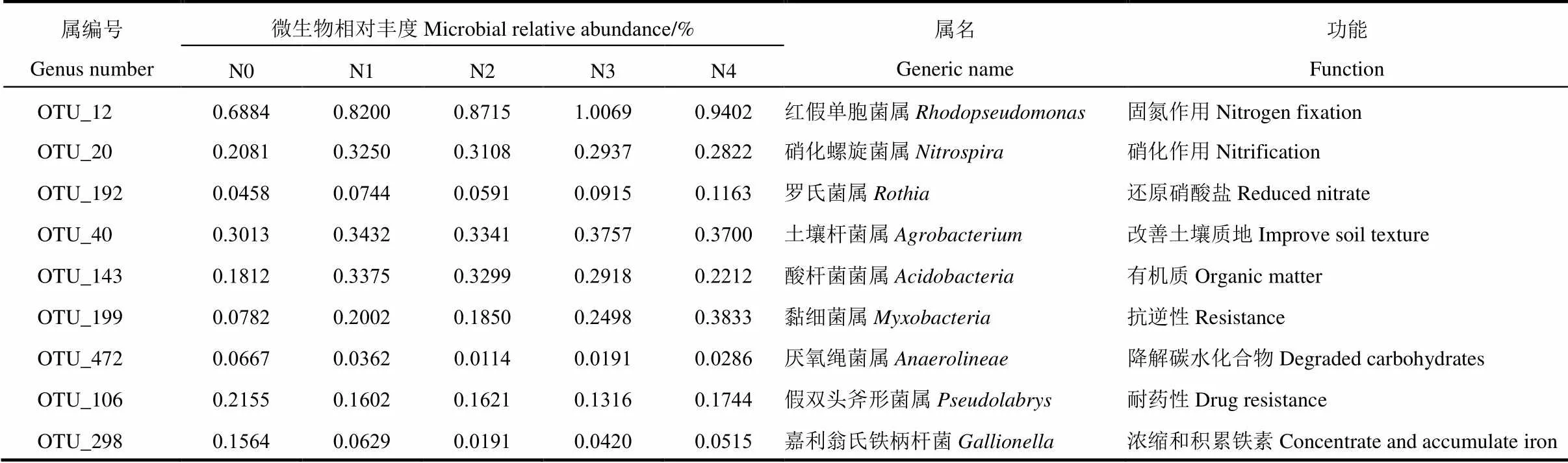
表8 头季水稻生长后期根际土壤中主要功能菌及其相对丰度
作物根系的形态和机能具有品种的固有遗传特性,同时也受到环境的影响[31]。土壤是作物根生长的基础,也是作物高产的基础。土壤微生物种群结构及土壤酶活性是土壤微生态环境中生理活性最强的部分。其中,土壤酶主要来源于土壤微生物和植物根系的分泌物,在物质循环和营养转化过程中起着关键作用[32]。土壤中蔗糖酶对增加土壤中易溶性营养物质起重要作用,脲酶可以表征土壤的氮素供应状况。在本研究中,头季不同氮肥运筹方式下前氮后移的N1、N2处理头季齐穗期和成熟期及再生季齐穗期,这两种酶活性均著高于常规施肥N4处理和对照N0处理;N1、N2处理的过氧化物酶和硝酸还原酶在头季齐穗与成熟期显著高于常规施肥N4处理,在再生季各施肥处理显著高于对照,但施肥处理间不显著。前人研究也表明前氮后移有利于土壤协调发展,提高土壤脲酶、蔗糖酶等养分循环有关的酶活性,提高土壤有机质含量和土壤养分有效性[33-35],这与本研究结果一致。张福锁等[36]认为栽培调控的实质是调控根际环境, 为作物创造良好的根际生态环境从而调控作物、土壤与微生物间的平衡,达到促进作物生长发育的目的。本研究进一步的相关分析表明,根系伤流强度与头季齐穗期至再生季成熟期土壤相关酶活性均达到显著或极显著正相关,尤其是头季齐穗至成熟与土壤解毒相关的过氧化氢酶及与氮素营养相关的脲酶和硝酸还原酶的相关性要高于再生季齐穗至成熟期。由此可见,再生稻生长期间尤其头季稻生长后期土壤养分循环有关的酶活性的协调,有利再生稻根系的生长从而促进植株对营养成分的吸收,实现水稻高产高效的目的。
根际土壤微生物是根际土壤酶的重要来源, 同时也直接参与根际的生物过程。本研究细菌 16S rDNA 高通量测序表明施肥处理与不施肥处理细菌群落结构多样性有显著差异,机械化栽培中,前氮后移的N1处理细菌 Chao1 指数和香农指数显著高于其他处理。菌群群落构成分析表明, 各施肥处理芽单胞菌门高于不施肥处理,各施肥处理中,N1处理下变形菌门、绿弯菌门和硝化螺旋菌门相对丰度均最高。在属水平上,各处理检测到念珠菌属、地杆菌属、氨基酸杆菌属等,与不施肥(N0)处理相比,施肥处理中的氨基酸杆菌属较多,主要有利于发酵土壤氨基酸,提高土壤化能有机营养能力。在属水平下主要细菌属功能分析(表8)表明,各施肥处理中红假细胞菌属、黏细菌属等微生物的提升在很大程度上能够改善土壤质地,有效促进土壤的碳氮循环,提高土壤抗逆性[37],尤其是N1处理硝化螺旋菌属丰度最高,可以利用尿素作为营养来源,在它们的基因组中编码的脲酶可以将尿素分解为土壤所需养分,促进土壤氮素循环[38-41],加之酸杆菌菌属丰度的提高有利增加土壤有机质含量,提高土壤肥力,为水稻的生长发育提供良好的微生物环境。
4 结论
综上所述,再生稻机械化栽培在头季稻总氮量保持不变情况下,氮肥适当后移的运筹方式,可提高再生稻根际土壤微生物多样性,增加根际土壤中促进碳氮循环及提高土壤抗逆性密切相关的细菌数量,有利协调土壤养分循环有关的酶活性,从而活化再生稻尤其是头季生长后期根际功能,促进根系对养分的吸收,达到两季水稻高产。在本研究中,N1处理(基肥∶一次分蘖肥∶二次分蘖肥∶孕穗肥=3∶1∶2∶4)最有利根际机能的改善,两季水稻总产量最高。
[1] 林文雄, 陈鸿飞, 张志兴, 徐倩华, 屠乃美, 方长旬, 任万军.再生稻产量形成的生理生态特性与关键栽培技术的研究与展望[J].中国生态农业学报, 2015, 23(4): 392-401.
Lin W X, Chen H F, Zhang Z X, Xu Q H, Tu N M, Fang C X, Ren W J.Research and prospect on physio-ecological properties of ratoon rice yield formation and its key cultivation technology[J]., 2015, 23(4): 392-401.(in Chinese with English abstract)
[2] 施能浦, 焦世纯.中国再生稻栽培[M].北京: 中国农业出版社, 1999.
Shi N P, Jiao S C.Ratooning rice cultivation in China.Beijin: China Agriculture Press, 1999.(in Chinese)
[3] 林文, 李义珍, 姜照伟, 郑景生.再生稻根系形态和机能的品种间差异及与产量的关联性[J].福建农业学报, 2001, 16(1): 1-4.
Lin W, Li Y Z, Jiang Z W, Zheng J S.Morphologic and functional difference of root systems among ratooning rice varieties and its correlation with yield[J]., 2001, 16(1): 1-4.(in Chinese with English abstract)
[4] 林文, 张上守, 姜照伟, 郑景生, 李义珍.再生稻产量与根系机能的相关性[J].福建稻麦科技, 2001, 19(4): 9-11.
Lin W, Zhang S S, Jiang Z W, Zheng J S, Li Y Z.Correlation between yield and root function of ratooning rice[J]., 2001, 19(4): 9-11.(in Chinese)
[5] 郑景生, 林文, 卓传营, 方宣钧, 林文雄.再生稻根干物质量及根系活力与产量的相关性研究[J].中国生态农业学报, 2004, 12(4): 106-109.
Zheng J S, Lin W, Zhuo C Y, Fang X J, Lin W X.The correlation of dry biomass and activity of root system with grain yield in raton rice(L) [J]., 2004, 12(4): 106-109.(in Chinese with English abstract)
[6] 张林, 郭晓艺, 刘茂, 熊洪, 朱永川, 周兴兵, 徐富贤.杂交中稻再生芽生长的影响因素研究[J].西南农业学报, 2010, 23(2): 309-314.
Zhang L, Guo X Y, Liu M, Xiong H, Zhu Y C, Zhou X B, Xu F X.Factors for promoting rationing bud growth of mid-season hybrid rice[J]., 2010, 23(2): 309-314.(in Chinese with English abstract)
[7] 饶鸣钿, 陈少庭, 郑履端, 廖朝阳.头季稻栽培因素对两优2186再生稻产量的影响[J].河南职业技术师范学院学报, 2002, 30(4): 12-15.
Rao M D, Chen S T, Zheng L D, Liao C Y.The effect of cultural factor of first seasonal rice to yield in ratoon rice of Liangyou 2186 [J]., 2002, 30(4), 12-15.(in Chinese with English abstract)
[8] 陈鸿飞, 杨东, 梁义元, 张志兴, 梁康迳, 林文雄.头季稻氮肥运筹对再生稻干物质积累、产量及氮素利用率的影响[J].中国生态农业学报, 2010, 18(1): 50-56.
Chen H F, Yang D, Liang Y Y, Zhang Z X, Liang K J, Lin W X.Effect of nitrogen application strategy in the first cropping rice on dry matter accumulation, grain yield and nitrogen utilization efficiency of the first cropping rice and its ratoon rice crop[J]., 2010, 18(1), 50-56.(in Chinese with English abstract)
[9] 杨东, 陈鸿飞, 卓传营, 林文雄.头季不同施氮方式对再生稻生理生化的影响[J].中国生态农业学报, 2009, 17(4): 643-646.
Yang D, Chen H F, Zhuo C Y, Lin W X.Effect of different N application modes in the first cropping rice on the physiobiochemistry of the first cropping rice and its ratoon rice[J]., 2009, 17(4), 643-646.(in Chinese with English abstract)
[10] 杨东.再生稻不同施氮方式的增产效应研究[D].福州: 福建农林大学, 2007.
Yang D.Study on the effect of different nitrogen application methods on increasing yield of ratooning rice[D].Fuzhou: Fujian Agriculture and Forestry University, 2007.(in Chinese with English abstract)
[11] 王森, 莫菁华, 汪洋, 游秋香, 任涛, 丛日环, 李小坤.水稻-再生稻体系干物质积累及氮磷钾养分的吸收利用[J].中国水稻科学, 2018, 32(1): 67-77.
Wang S, Mo J H, Wang Y, You Q X, Ren T, Cong R H, Li X K.Dry matter accumulation and N, P, K absorption and utilization in rice-ratoon rice system[J]., 2018, 32(1): 67-77.(in Chinese with English abstract)
[12] 郑景生, 林文雄, 李义珍, 姜照伟, 卓传营.再生稻头季不同施氮水平的双季氮素吸收及产量效应研究[J].中国生态农业学报, 2004, 12(3): 78-82.
Zheng J S, Lin W X, Li Y Z, Jiang Z W, Zhuo C Y.Nitrogen uptake and grain yield effects of double-cropping rice at different nitrogen application rates in the first crop of ratoon rice[J]..2004, 12(3): 78-82.(in Chinese with English abstract)
[13] 任天举, 李经勇, 唐永群, 邹亚兰.15N示踪研究再生稻施用氮肥的吸收分配和效应[J].西南师范大学学报: 自然科学版, 2009, 34(3): 132-136.
Ren T J, Li J Y, Tang Y Q, Zou Y L.15N Tracer study ratooning rice applied nitrogen absorption, assignment and effect [J].:.2009, 34(3): 132-136.(in Chinese with English abstract)
[14] 林燕, 邓学东, 李泽林.促芽肥施用时间与施肥量对再生稻的影响[J].农技服务, 2015, 32(4): 93-94.
Lin Y, Deng X D, Li Z L.Effects of application time and dosage of shoot-promoting fertilizer on regenerated rice[J]., 2015, 32(4): 93-94.(in Chinese).
[15] 王森.水稻—再生稻体系养分需求特性及氮肥合理运筹初探[D].武汉: 华中农业大学, 2017.
Wang S.Characteristics of nutrient requirements and reasonable management of nitrogen fertilizer in rice-ratoon rice system[D].Wuhan: Huazhong Agricultural University, 2017.(in Chinese with English abstract).
[16] 于林惠.对机插水稻生育特点及管理对策的初步探讨[J].中国农机化, 2002 (1): 29-31.
Yu L H.Preliminary discussion on the growth characteristics and management measures of machine- transplanted rice[J]., 2002, 1: 29-31.(in Chinese)
[17] 徐红, 李成, 于凤翠, 唐钧, 徐晓青, 唐玮, 马卉.机插水稻生育特性及高产配套技术[J].安徽农学通报, 2011, 17(23): 150-151.
Xu H, Li C, Yu F C, Tang J, Xu X Q, Tang W, Ma H.Growth characteristics of machine-transplanted rice and its high yield cultivation technique[J]., 2011, 17(23):150-151.(in Chinese)
[18] 茅弼华, 王和平, 王志林.机插水稻的生育特性和有关农艺技术[J].江苏农业科学, 2006, 3: 27-30.
Mao B H, Wang H P, Wang Z L.Growth characteristics and related agronomic techniques of machine- transplanted rice[J]., 2006, 3: 27-30.(in Chinese)
[19] 兰陆寿.机插水稻生育特性及栽培技术[J].现代农业科技, 2011, 18: 56-57.
Lan L S.Growth characteristics of machine-transplanted rice and its cultivation technique[J]., 2011, 18: 56-57.(in Chinese)
[20] Zhang R, Jorge M Vivanco, Shen Q.The unseen rhizosphere root-soil-microbe interactions for crop production[J]., 2017, 37: 8-14.DOI: 10.1016/ j.mib.2017.03.008
[21] 李鸿波, 吴朝晖.水稻根际微生物的影响因素研究进展[J].杂交水稻, 2018, 33(4): 1-6, 54.
Li H B, Wu Z H.Research progress on factors influencing rhizosphere microorganisms of rice[J]., 2018, 33(4): 1-6, 54.(in Chinese with English abstract)
[22] 王孝林, 王二涛.根际微生物促进水稻氮利用的机制[J].植物学报 2019, 54 (3): 285-287.
Wang X L, Wang E T.connects root microbiota and nitrogen use in rice[J]., 2019, 54(3): 285-287.(in Chinese with English abstract)
[23] 李合生.植物生理生化实验原理和技术[M].北京: 高等教育出版社, 2000.
Li H S.Principles and techniques of plant physiology and biochemistry experiments[M].Beijing: Higher Education Press, 2000.
[24] 关松荫, 张德生, 张志明.土壤酶及其研究方法[M].北京: 农业出版社, 1986.
Guan S Y, Zhang D S, Zhang Z M.Soil enzymes and its research methods[M].Beijing: Agricultural Press, 1986.
[25] Alef K, Nannipieri P.Methods in applied soil microbiology and biochemistry[M].New York: Academic Press, 1995.
[26] Wang Y, Sheng, H F, He Y, Wu J Y,Jiang Y X, Tam N F, Zhou H W.Comparison of the levels of bacterial diversity in fresh water, intertidal wetland, and marine sediments by using millions of illumina tags., 2012, 78(23): 8264.DOI: 10.1128/AEM.01821-12.
[27] 张晶, 林先贵, 尹睿.参与土壤氮素循环的微生物功能基因多样性研究进展[J].中国农业生态学报: 2009, 17(5), 1029-1034.
Zhang J, Lin X G, Yin R.Advances in functional gene diversity of microorganism in relation to soil nitrogen cycling[J]., 2009, 17(5), 1029-1034.(in Chinese with English abstract)
[28] 中国科学院微生物研究所《伯杰细菌鉴定手册》翻译组译.伯杰细菌鉴定手册(第八版)[M].北京: 科学出版社, 1984.
Translation team of Bergey’s manual of systematic bacteriology, Institute of Microbiology, Chinese Academy of Sciences.Bergey’s manual of systematic bacteriology (Eighth edition)[M].Beijing: Science Press, 1984.
[29] 林超峰, 龚骏.嗜中性微好氧铁氧化菌研究进展[J].生态学报, 2012, 32(18): 5889-5899.
Lin C F, Gong J.Recent progress in research on neutrophilic, microaerophilic iron(II) -oxidizing bacteria [J]., 2012, 32(18): 5889-5899.(in Chinese with English abstract)
[30] 刘富贵, 王学栋, 吴跃进.再生稻根系栽培生理初探[J].安徽农业科学, 1990(2): 105-109.
Liu F G, Wang X D, Wu Y J.A preliminary study on the root culture physiology of ratoon rice [J]., 1990(2): 105-109.(in Chinese)
[31] 潘晓华, 王永锐, 傅家瑞.水稻根系生长生理的研究进展[J].植物学通报, 1996, 13(2): 13-20.
Pan X H, Wang Y R, Fu J R.Advance in the study on the growth-physiology in rice of root system() [J]., 1996, 13(2): 13-20.(in Chinese with English abstract)
[32] 邱莉萍, 刘军, 王益权, 孙慧敏, 和文祥.土壤酶活性与土壤肥力的关系研究[J].植物营养与肥料学报, 2004, 10(3): 277-280.
Qiu L P, Liu J, Wang Y Q, Sun H M, He W X.Research on relationship between soil enzyme activities and soil fertility[J]., 2004, 10(3): 277-280.(in Chinese with English abstract)
[33] 陈军, 黄珊瑜, 刘冰, 吴林坤, 林文雄.不同氮肥运筹对水稻根际土壤理化性质及代谢物质的影响[J].福建农业学报, 2015, 30(11): 1082-1089.
Chen J, Huang S Y, Liu B, Wu L K, Lin W X.Effects of different nitrogen regimes on soil physico-chemical properties and metabolites in rice rhizosphere [J]., 2015, 30(11): 1082-1089.(in Chinese with English abstract) .
[34] 陈龙怀.氮肥运筹调控下水稻根际土壤生物学特性分析[D].福州:福建农林大学, 2012.
Chen L H.Biological characteristics of rice rhizosphere soil under the control of nitrogen fertilizer application[D].Fuzhou: Fujian Agriculture and Forestry University, 2012.(in Chinese with English abstract).
[35] 张玉, 秦华东, 黄敏, 江立庚, 徐世宏.氮肥运筹对免耕水稻根系生长、根际土壤特性及产量的影响[J].广西植物, 2014, 34(5): 681-685.
Zhang Y, Qin H D, Huang M, Jiang L G, Xu S H.Effect of different nitrogen application modes on root growth,rhizosphere soil characteristics and rice yield under no-tillage[J]., 2014, 34(5): 681-685.(in Chinese with English abstract)
[36] 张福锁, 王激清, 张卫峰, 崔振岭, 马文奇, 陈新平, 江荣风.中国主要粮食作物肥料利用率现状与提高途径[J].土壤学报, 2008, 45(5): 915-924.
Zhang F S, Wang J Q, Zhang W F, Cui Z L, Ma W Q, Chen X P, Jiang R F.Nutrient use efficiencies of major cereal crops in China and measures for improvement[J].2008, 45(5): 915-924.(in Chinese with English abstract)
[37] Men S, Ding F, Zhang H, Ke C, Zhang S, Huang Z.Effects of Nano-carbon humic acid water-retaining fertilizer on citrus growth and the soil bacterial community in citrus field[J]., 2018, 9(6): 84-89.
[38] Zhang Q, Li Y, He Y, Liu H, Dumont M G, Brookes P, Xu J.Nitrosospira cluster 3-like bacterial ammonia oxidizers and Nitrospira-like nitrite oxidizers dominate nitrification activity in acidic terrace paddy soils [J]., 2019.DOI: https://doi.org/10.1016/j.soilbio.2019.01.006
[39] Han S, Zeng L, Luo X, Xiong X, Wen S, Wang B, Chen W, Huang Q.Shifts in Nitrobacter- and Nitrospira-like nitrite-oxidizing bacterial communities under long-term fertilization practices[J]., 2018, 124: 118-125.
[40] Jin S, Chen Z, Li Y, Xia T, Ren Z, Yin L.Study on endophytic bacteria diversity in the root of peperomia dindygulensis Miq.by applying clone library construction method[J]., 2017, 8(6): 13-16.
[41] Li H, Chi Z, Yan B.Long-term impacts of graphene oxide and Ag nanoparticles on anammox process:Performance, microbial community and toxic mechanism., 2019, 79(5): 239-247.
Nitrogen Fertilizer Management for Main Crop Rice and Its Carrying-over Effect on Rhizosphere Function and Yield of Ratoon Rice
HUANG Jinwen1,#, WU Jiayi1, #, CHEN Hongfei1, ZHANG Zhixing1, FANG Changxun1, SHAO Caihong2, LIN Weiwei1, WENG Peiying1, LIN Wenxiong1, *
(Key Laboratory for Genetics Breeding and Multiple Utilization of Crops, Ministry of Education/ College of Agriculture, Fujian Agriculture and Forestry University/Key Laboratory of Crop Ecology and Molecular Physiology, Fuzhou 350002, China; Institute of Soil Fertilizer and Resources Environment, Jiangxi Academy of Agricultural Sciences, Nanchang 330200, China; )
【Objective】Ratoon rice is the continuation of the main crop.We attempt to elucidate the effect of mechanical transplanting of main crop together with nitrogen fertilizer management on the yield of the main crop and ratoon rice from the perspective of rhizosphere microecosystem, which may provide a reference for the high-yielding and efficient cultivation of ratoon rice.【Method】The rice genotype of Yongyou 1540 was used as the material.At a total nitrogen application level of 225.00 kg/hm2in the main crop, different nitrogen fertilizer treatments were designed to analyze the effects of mechanical cultivation on the yield, root bleeding intensity, enzyme activities and microbial diversity in rhizosphere soil in two consecutive years in paddy field.【Result】In N1 and N2 treatments, the root system maintained higher vigor, especially in the late growing period of the main season in the N1 treatment (basal fertilizer : primary fertilizer for tillering : secondary fertilizer for tillering : fertilizer for panicle initiation= 3 : 1 : 2 : 4) showed a significant increase in root bleeding intensity as compared with the control N0 and the local conventional treatment of N4.The total yield of the N1 treatment was the highest in the two growing seasons, reaching an average of 17 351.23 kg/hm2, 23.00% higher than that of N4.Soil enzyme activity analysis showed that the activities of sucrase followed the trend of N2>N1>N3>N4>N0 in the full heading stage during the main season, but N1>N2> N3>N3>N4>N0 for ratoon rice; the activities of catalase and nitrate reductase both followed a trend of N1>N2> N3>N4>N0 in the full heading stage during the main season; the activities of urease, N1>N2> N3>N4>N0 from the heading stage of the main crop to the heading stage of ratoon rice.Highly significant positive correlations were observed between soil enzyme activities and root bleeding intensity at all growing stages.Bacterial 16S rDNA sequencing showed that the structural diversity of bacterial communities differed significantly between the fertilized and non-fertilized treatments, and the Chao1 index and Shannon index of N1 treatment were significantly higher than those of other treatments.Function analysis of rhizospheric bacterial community showed that fertilization increased the number of bacteria closely related to soil carbon and nitrogen cycle, soil organic matter content and soil stress resistance, especially the N1 and N2 treatment with higher relative abundance of, which were favorable to soil nitrogen cycle.【Conclusion】Under mechanical cultivation, the treatment with properly postponed nitrogen fertilizer application in the main crop is beneficial to the improvement of the rhizosphere microecosystem of ratoon rice, thus improving root vigor, promoting rice growth, and obtaining high yield of rice in both growing seasons.
ratoon rice; nitrogen management; root activity; rhizosphere; community diversity
10.16819/j.1001-7216.2021.200603
2020-06-04;
2020-08-30。
国家重点研发计划资助项目(2016YFD0300508,2017YFD0301602,2018YFD0301105)。
猜你喜欢
农村百事通(2023年7期)2023-08-05 09:18:56
农业科技通讯(2023年1期)2023-02-12 07:09:56
落叶果树(2022年1期)2022-11-16 17:02:29
农业工程学报(2022年14期)2022-10-19 01:15:54
福建稻麦科技(2019年2期)2019-07-11 06:55:48
农家之友(2018年12期)2018-03-12 00:50:28
西北园艺(果树)(2018年4期)2018-02-12 02:31:57
农村百事通(2015年24期)2015-12-28 11:01:03
河南林业科技(2015年2期)2015-06-26 01:44:20
四川农业科技(2015年3期)2015-03-21 11:13:34
