
改变碳输入对沂蒙山区典型次生林土壤微生物碳源代谢功能的影响
2021-07-12 01:39李秋梅黎胜杰王欣丽张广娜梅鹤平
生态学报 2021年10期
李秋梅,黎胜杰,王欣丽,刘 波,张广娜,张 弛,高 远,梅鹤平,王 芸,*
1 山东省水土保持与环境保育重点实验室,临沂大学资源环境学院,临沂 276000
2 临沂大学农林科学学院,临沂 276000
3 山东省费县塔山国有林场,临沂 273421
凋落物和根系向土壤中的有机物输入是森林生态系统营养物质循环的两个途径,输入有机物的质量和数量可影响土壤碳汇功能和生产力,也影响土壤微生物的碳源供给。土壤微生物在土壤肥力维持和有机物分解中起重要作用。高质量(高营养、易分解)的有机物可被土壤微生物高效利用[1],而低质量的有机物可能会导致土壤营养匮缺从而抑制微生物代谢[2]。有机物的数量和质量影响土壤微生物活性[3],进而导致不同碳转化过程的差异。
凋落物输入途径和数量对土壤碳代谢功能的影响存在争议。尽管有研究表明,添加凋落物抑制美国Andrews温带针叶林土壤有机碳分解[4],但一般而言,添加地上凋落物刺激土壤呼吸。尽管有研究表明,去除凋落物增加温带阔叶林土壤微生物量碳[5],但大多研究表明去除凋落物降低土壤呼吸[6],降低土壤微生物碳源代谢功能[2]。限制凋落物碳输入(去除凋落物和无输入)显著减少土壤有机碳含量[7],降低土壤呼吸[8]。
根系所产生的根际沉积和根凋落物为土壤微生物的重要碳源[9]。根系输入一方面通过土壤微生物群落更有效地合成与矿物相关的土壤有机碳[9],另一方面根际引发效应加速土壤有机质的分解[10]。去除根系降低土壤有机碳含量[7]。根系对细菌和真菌的影响不同,真菌更喜欢来源于根系的碳,而不是微环境中的碳;细菌只有在无可替代碳源的情况下,才加强对源于根的碳利用[11]。去除根系降低阔叶林和针叶林土壤微生物生物量[12],降低温带针叶林土壤有机碳分解[4]。去除根系降低栓皮栎(Quercusvariabilis)人工林和黑松(Pinusthunbergii)人工林土壤微生物的碳源代谢功能[2]。去除根系降低土壤有机磷含量[13],进而降低微生物碳代谢。总之,去除根系会降低底物可利用性和有机碳的激发效应,导致土壤微生物群落结构和功能变化,但对微生物碳代谢功能的影响还存在争议,并受环境条件的影响。
综上所述,凋落物和根系对土壤微生物碳代谢功能的影响随气候带(如亚热带和温带)、植被类型(如针叶林和阔叶林)改变而变化,对土壤微生物碳源代谢功能的影响还存在争议[2]。鉴于凋落物和根系主要成分为碳,简称改变凋落物和根系输入为改变碳输入。
沂蒙山区位于温带的鲁中南低山丘陵区,其主要森林类型之一为栎类次生林。本文期望揭示改变碳输入途径(地上与地下)、数量(凋落物倍增、凋落物去除与对照)对土壤微生物碳源代谢功能的定量影响,深化对温带天然次生林土壤有机碳代谢过程的认识。
1 材料与方法
1.1 样地概况与样品采集
试验在山东省费县塔山国家森林公园(35°10′—36°00′ N, 117°35′—118°20′ E)开展。当地年均温13℃,平均年降水量700 mm,土壤为棕壤。选取土地利用历史相似、林龄约为40 a 的栎类天然次生林(属针阔混交林),于2014 年11 月开展凋落物添加/去除和根系去除定位控制试验。试验采取随机区组试验设计,因素为改变碳输入,共设置5 个处理,包括对照(不进行任何处理)、凋落物倍增、去除凋落物、去除根系和无输入处理(去除凋落物和根系),5 次重复。随机选取5 个样地,每个样地上选取一个20 m × 30 m 的样方。在每个样方上设置5 个1.5 m × 1.5 m 的小样方,每个小样方随机设置一个处理(表1),所有选取的小样方内均没有乔木。凋落物添加/去除处理每月开展一次。根系去除处理每月开展一次,其中,夏季每月两次。
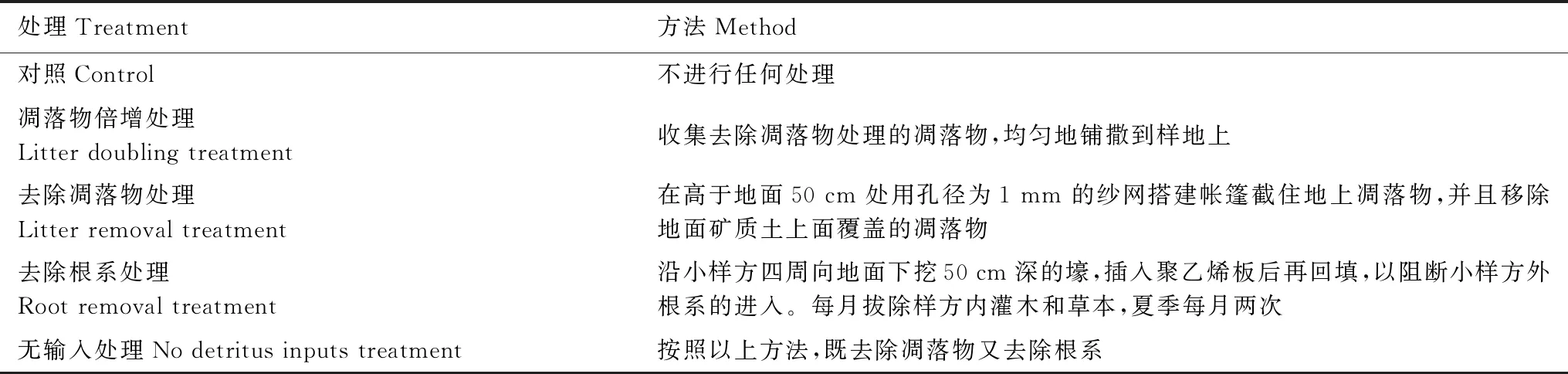
表1 凋落物添加/去除和根系去除试验描述
于定位实验开展21 个月后,即于2016 年8 月13 日取土样,取样深度0—10 cm。在每个小样方取5 个土钻(分布在样方四个角和中心,直径3.5 cm)的土,混合为一个土样。共取25 个土样。土样置于4℃冷藏箱运回实验室。一部分土样用于4℃保存,过2 mm 筛,分析土壤微生物碳代谢功能;其余土样风干,分析理化性质。
1.2 实验室分析
土壤微生物碳代谢功能采用群落水平代谢图谱法表征,用Biolog EcoPlateTM(Biolog Inc., Hayward, CA, USA) 测定[14]。取相当于10 g 干土的新鲜土样,加95 mL无菌的0.85%的NaCl溶液振荡30 min,取稀释1000 倍的150 μL土壤溶液接种至Biolog Eco 微平板中,每个土样接种到一个微孔板上,即每个土样接种3次。然后置于25℃恒温培养箱培养240 h,每12 h于酶标仪(Biotek, Synergy H1)上读数1 次,波长为590 nm。
土壤有机碳含量采用元素分析仪测定(vario MACRO cube, Elementar, Germany)。土壤含水量采用烘干法。pH值采用1∶5 的土水比测试[15]。有效磷含量采用树脂法[16],铵态氮、硝态氮含量采用1∶5 的KCl溶液提取,铵态氮采用水杨酸法测试[15],硝态氮采用双波长紫外分光光度法测试[17]。
1.3 统计分析
1.3.1土壤微生物碳代谢特征
用孔的平均颜色变化率(Average well color development,AWCD)来表征土壤微生物碳代谢特征[18]。AWCD越高,碳代谢能力越高。
(1)
式中,Ci是590 nm 处孔的吸光度。R是对照孔的吸光度。N是碳源的种类数,此处为31。其中负的Ci-r设置为0[19]。
31 孔的AWCD均值随时间t的变化,可用逻辑斯蒂增长方程来拟合[20]。
(2)
式中,K是AWCD 的理论最大值,s是达到最大值一半的时间,R是逻辑斯蒂增长曲线的指数斜率[20]。
由于96 h 是最接近达到AWCD最大值一半时间((93±2) h)的读数时间,所以96 h 的吸光度值用来计算土壤微生物的碳代谢丰富度、多样性和后续的冗余分析(Redundancy analysis, RDA)。
丰富度是在96 h 吸光度大于0.25 的碳源个数[21]。多样性采用Shannon多样性指数[22]。
H′=-∑piln(pi)
(3)
pi=ni/N
(4)
式中,ni为单个碳源吸光度,N为31 种碳源吸光度的和。
根据降解的相对难易程度,把Biolog Eco 微平板中的碳源分为六大类。其中,糖类、羧酸类和氨基酸类碳源归为活性碳;胺类、聚合物类和其它类碳源归为惰性碳。
1.3.2方差分析和冗余分析
采用单因素随机区组方差分析改变碳输入对土壤微生物碳源代谢功能的影响,多重比较采用Duncan法。冗余分析(Redundancy analysis,RDA)用于分析土壤微生物碳源代谢功能与土壤理化性质之间的关系。因变量为31 种碳源在96 h 的吸光度,自变量为土壤理化性质,包含pH、含水量、有机碳含量、全氮含量、铵态氮含量、硝态氮含量、有效磷含量。用Monta-Carlo(999次)检验显著影响土壤微生物碳源代谢的因子。Spearman相关分析用于分析土壤微生物碳源代谢多样性与土壤因子之间的关系。
2 结果与分析
2.1 改变碳输入对栎类天然次生林土壤理化性质的影响
去除根系处理和无输入处理均显著降低了土壤pH(P<0.05,表2)。凋落物倍增处理显著增加了土壤有机碳、铵态氮和硝态氮含量(P<0.05,表2),去除凋落物处理、去除根系处理和无输入处理均显著降低了土壤铵态氮含量(P<0.05);改变碳输入对土壤含水量、全氮、有效磷含量都没有显著影响(P>0.05)。

表2 改变碳输入对沂蒙山区栎类天然次生林土壤理化性质的影响
2.2 改变碳输入对栎类天然次生林土壤微生物碳源代谢功能的影响
与对照相比,凋落物倍增处理增加栎类天然次生林土壤微生物碳源代谢丰富度、碳源代谢多样性和碳源代谢强度(图1),但不显著(P>0.05)。去除凋落物处理显著降低土壤微生物碳源代谢丰富度和碳源代谢强度(P<0.05,图1)。去除根系处理和无输入处理都没有显著影响土壤微生物碳源代谢功能(P>0.05)。去除凋落物处理降低土壤微生物碳源代谢功能的幅度大于去除根系处理。

图1 改变碳输入对土壤微生物碳源代谢多样性的影响
凋落物倍增处理增加了土壤微生物对糖类碳源的代谢丰富度,去除凋落物处理、去除根系处理和无输入处理均降低了糖类的代谢丰富度,都不显著(P>0.05,表3)。无输入处理显著降低了胺类碳源的代谢丰富度(P<0.05)。其余处理对羧酸类、氨基酸类、聚合物类和其它类碳源代谢丰富度影响都不显著(P>0.05)。

表3 改变碳输入对土壤微生物六大类碳源代谢丰富度的影响

表4 改变碳输入对土壤微生物六大类碳源代谢Shannon多样性指数的影响

表5 改变碳输入对土壤微生物六大类碳源代谢强度的影响

图2 改变碳输入处理下微生物碳源代谢多样性与土壤因子之间的冗余分析
2.3 改变碳输入对栎类天然次生林土壤微生物碳源代谢功能的影响途径
土壤有机碳和铵态氮含量显著影响不同凋落物输入途径和数量变化条件下土壤微生物碳源代谢多样性,两者解释了土壤微生物碳源代谢多样性变异的 27.4% (F=5.06,P=0.001,图2),其中轴1 解释了24.0%的变异(F=8.45,P=0.001),轴2解释了3.4%的变异(F=1.79,P=0.086)。土壤微生物的碳源代谢丰富度、多样性、强度与土壤有机碳和铵态氮含量都显著正相关(P<0.05,表6)。凋落物倍增处理在铵态氮和有机碳含量增加的方向。去除凋落物处理、无输入处理都位于铵态氮和有机碳含量减少的方向(图2)。这表明凋落物倍增处理通过增加土壤铵态氮和有机碳含量(表2),增加土壤微生物碳源代谢功能,去除凋落物和无输入处理通过降低土壤铵态氮和有机碳含量(表2),降低土壤微生物碳源代谢功能。

表6 碳源代谢多样性与土壤因子之间的秩相关分析
3 讨论
3.1 改变碳输入对土壤微生物碳源代谢功能的影响
本研究中,凋落物倍增处理轻微增加土壤微生物碳源代谢Shannon多样性。尽管有研究表明凋落物倍增降低针叶林土壤微生物代谢活性[4],但综合本研究和大量国内外相关研究[2],凋落物倍增增加针阔混交林土壤微生物代谢活性。本研究中,凋落物倍增处理增加土壤微生物对糖类和胺类的代谢能力,降低对羧酸类、氨基酸类和聚合物类的代谢能力。凋落物倍增对针叶林土壤活性碳代谢能力(如糖类、淀粉、羧酸类、纤维素等代谢或代谢相关土壤酶活性(β-葡萄糖苷酶、蔗糖酶、纤维二糖酶等)或基因多样性)的影响争议较大[2, 23],但降低对惰性碳的代谢能力(如胺、聚合物、几丁质、角质、木质素等代谢或代谢相关土壤酶活性(芳基硫酸酯、β- 1, 4-N-乙酰葡糖氨糖苷酶、酚氧化酶、过氧化物酶等)或基因多样性)[2]。尽管有研究表明凋落物倍增降低阔叶林土壤微生物活性碳代谢,增加惰性碳代谢[24],但本研究和大多研究[25-26]都表明凋落物倍增处理增加针阔混交林土壤微生物活性碳代谢,降低惰性碳代谢。森林土壤微生物代谢基因会随凋落物输入的质量而变化,当外界输入复杂的有机物,土壤微生物基因更倾向于代谢复杂的有机物[2]。
本研究中,去除凋落物显著降低微生物碳源代谢能力。尽管有研究表明,去除凋落物增加针叶林和阔叶林土壤微生物生物量[5],但更多研究表明,去除凋落物降低针叶林和阔叶林土壤微生物量[27]、碳代谢活性[2,26]。本研究中,去除凋落物处理显著降低土壤微生物的活性碳代谢能力。尽管有研究发现去除凋落物显著增加活性碳代谢[24],但大多研究表明去除凋落物显著降低活性碳代谢,如显著降低栓皮栎人工林土壤微生物的羧酸类、淀粉代谢,显著降低黑松人工林土壤羧酸类的代谢[2],降低阔叶林土壤β-葡萄糖苷酶活性[25-26]。尽管有研究发现,去除凋落物处理显著增加阔叶林土壤惰性碳代谢[24],但大多研究表明去除凋落物显著降低惰性碳代谢[2,28]。本研究和其它大多研究表明,去除凋落物降低针阔混交林土壤微生物碳代谢能力。这可能是去除凋落物减少向土壤中的有机物输入,降低微生物可利用底物。
本研究中,去除根系处理轻微降低土壤微生物碳源代谢能力。尽管有研究表明,去除根系增加美国Andrews针叶林中土壤微生物生物量[27],但大多研究表明去除根系降低土壤微生物碳代谢功能[2,29]。本研究中去除根系处理降低土壤微生物对六大类碳源的代谢能力。大多研究表明,去除根系降低活性碳库代谢[2]。在阔叶林中,去除根系显著降低土壤β-葡萄糖苷酶活性[26-27]。去除根系降低惰性碳库的利用或与惰性碳库相关酶的活性,例如降低黑松人工林中土壤微生物的胺类和聚合物类碳源代谢能力[2]。根系分泌物如糖类、氨基酸、酶、脂肪族和芳香族等化合物,可作为营养物质促进微生物生长[30],而根系分泌物中的酚类物质会抑制土壤微生物活性。根系共生的菌根真菌储存了大量光合固定的碳[3]。本研究和其它大多研究表明,去除根系降低土壤微生物碳源代谢功能。
本研究中,无输入处理降低了土壤微生物碳源代谢能力。尽管在美国Andrews针叶林中无输入处理增加土壤微生物生物量[27],但本研究与其他大多研究表明,无输入处理降低土壤碳代谢功能[2,26]。同时去除凋落物和根系,土壤微生物代谢受可利用底物限制[31],碳代谢功能降低。
本研究中,去除凋落物处理降低微生物碳源代谢功能的幅度大于去除根系处理,表明在控制实验开展21 个月的时间内,凋落物对沂蒙山区次生林土壤微生物碳源代谢功能的影响大于根系。温带落叶阔叶林中,根系对土壤呼吸的影响大于凋落物[32],根系对针叶林土壤有机碳含量的影响大于凋落物[3]。去除根系降低我国亚热带针叶林(侧柏(Platycladusorientalis)人工林)土壤有机碳的程度大于去除凋落物[33]。原因为去除根系降低土壤大团聚体含量及其中的有机碳含量并减少大团聚体的粘合剂-真菌生物量;而去除凋落物未引起上述土壤属性变化[33]。与地上碳输入相比,土壤微生物能更有效的利用地下碳输入形成稳定的土壤有机碳[34]。但在我国亚热带阔叶林(厚夹相思(Acaciacrassicapa)林和尾叶桉(Eucalyptusurophylla)林)中,从生物量和分解所产生的碳来说,凋落物对土壤有机碳含量的影响大于根系[7]。在热带人工林,凋落物对土壤呼吸和微生物生物量的影响大于根系,而在次生林中则相反[35]。凋落物和根系对土壤微生物碳代谢的影响随林型和温度带而变化。
需要指出的是,本研究采用挖壕沟法去除根系,残留根系大部分未降解[36],会增加微生物可利用的碳源,进而增加微生物碳代谢能力。由于残留根系中不同化学成分降解速率不同[37],且不同根系粗细[37]和级别[36]分解速率不同,如细根比粗根养分含量高[38]但分解慢[37],难以对碳源代谢能力的影响定量化。考虑到壕沟里残留根系的贡献,凋落物和根系对土壤微生物碳代谢能力的相对影响可能会发生变化,未来的研究需要考虑到残留根系对土壤微生物碳源代谢的贡献。
3.2 改变碳输入条件下土壤碳源代谢的影响因素
冗余分析表明,土壤有机碳和铵态氮含量显著影响不同碳输入处理条件下土壤微生物碳源代谢多样性。有研究表明,土壤铵态氮、硝态氮含量显著影响黑松人工林中不同碳输入条件下土壤微生物碳源代谢功能多样性[2];而土壤速效磷、含水量显著影响栓皮栎人工林中不同碳输入条件下微生物碳源代谢功能多样性[2]。这与改变碳输入处理下,不同林型下土壤微生物代谢的影响因子不同有关。
冗余分析图中,凋落物倍增处理分布在土壤铵态氮和有机碳含量增加的方向,这表明凋落物倍增处理通过显著增加土壤有机碳和铵态氮含量(表2),改变土壤微生物碳源代谢图谱(图2),并增加微生物碳源代谢功能(表6)。去除凋落物处理、无输入处理都位于土壤铵态氮和有机碳减少的方向(图2),这表明这两个处理通过降低土壤有机碳和铵态氮含量(表2),改变微生物碳源代谢图谱(图2),降低微生物碳源代谢功能(表6)。凋落物倍增为微生物生长活动提供了碳源和氮源,刺激土壤微生物活性[39],如凋落物倍增增强华北落叶松(Larixprincipis-rupprechtii)人工林土壤呼吸[40]。沂蒙山区栎类天然次生林是针阔混交林,叶凋落物碳氮比为40,而低碳氮比有利于提高土壤有机氮净矿化速率[41],增加微生物碳源代谢能力。去除凋落物阻断了地上有机物输入,减少了碳源输入,降低微生物碳源代谢功能。
4 结论
采用Biolog Eco微平板培养法,研究改变碳输入对沂蒙山区栎类次生林土壤微生物碳源代谢功能的影响。结果表明,凋落物倍增处理增加了土壤微生物碳源代谢功能,增加了对糖类和胺类的利用能力。去除凋落物处理、去除根系处理、无输入处理降低了土壤微生物的碳源代谢功能。凋落物对土壤微生物碳源代谢能力的影响大于根系。土壤有机碳含量、铵态氮含量显著影响微生物碳源代谢多样性(P<0.05),并与碳源代谢功能正相关。这表明凋落物倍增处理通过显著增加土壤有机碳含量和铵态氮含量(P<0.05),改变土壤微生物碳源代谢图谱,增加土壤微生物碳源代谢功能。去除凋落物处理、无输入处理通过降低土壤有机碳含量和铵态氮含量,改变微生物碳源代谢图谱,降低微生物碳源代谢功能。结果深化了改变碳输入途径(地上凋落物与地下根系)、数量(凋落物倍增、去除凋落物与对照)对温带栎类天然次生林土壤微生物碳代谢过程的认识。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
土壤学报(2022年2期)2022-06-09
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
现代农业研究(2017年11期)2018-01-12
中国农业文摘-农业工程(2017年2期)2017-01-12
农业与技术(2016年6期)2016-05-14
