
水生生态系统食物网复杂性与多样性的关系
2021-07-12 00:06李晓晓崔保山邵冬冬
生态学报 2021年10期
李晓晓,杨 薇,2,*孙 涛,2, 崔保山,2,邵冬冬,2
1 北京师范大学环境学院,水环境模拟国家重点实验室, 北京 100875
2 黄河口湿地生态系统教育部野外科学观测研究站, 东营 257500
食物网是生态系统中以多种生物作为节点、其间捕食关系作为营养链接形成的网络,认识食物网的复杂结构是生态学的中心问题之一[1- 5]。食物网的节点数量称为食物网多样性,用于反映食物网的大小[4- 5]。食物网的链接密度和连通度可系统反映食物网内部各个节点联系的紧密程度,体现食物网的复杂性[4- 5]。
食物网的多样性(节点数)和复杂性(链接数、链接密度、连通度)均是食物网研究中最重要的概念之一,渗透到食物网结构、能流、稳定等多领域研究中。例如May[6]提出食物网的稳定性与食物网多样性和复杂性均有直接的关系,且二者的乘积越小食物网越稳定;目前广泛使用的生态位模型[7]则仅需要输入节点数和连通度建立食物网的拓扑结构。这些研究均要求对于自然生态系统食物网多样性和复杂性之间的关系有科学论述[8]。在过去的几十年中,生态学家致力于探索食物网复杂性与多样性之间的普适关系。早期研究普遍认为存在链接-物种缩放法则[9- 10](link-species scaling law),即多样性不同的食物网具有相等的链接密度。后有研究提出了连通度常数假设[11](constant connectance),指食物网随着多样性的增加,其连通度不变。考虑到两种假设的不一致性,仍需进一步探究食物网的复杂性和多样性间的关系。
本研究基于一个水生生态系统食物网数据集Tromodels[12-13],并补充了课题组在黄河口海草床建立的食物网(待发表),共涉及河口、湖泊、海洋和河流四种生态系统类型,探究水生生态系统的食物网复杂性(链接数、链接密度和连通度)与其多样性(节点数)的关系,验证早期提出的链接-物种缩放法则和连通度常数法则,探究是否存在不同水生生态系统类型均适用的普适性规律,以期为了解自然生态系统多样性和复杂性间的关系提供理论支撑。
1 材料与方法
1.1 水生生态系统食物网案例
本研究共选取48个水生食物网案例(图1),其中47个食物网案例源于R软件中enaR安装包的一个实际食物网数据集TroModels[12- 13],食物网均采用生态网络分析的方法构建。另外1个为本课题组在黄河口海草床构建的食物网(待发表),采用文献调研结合稳定同位素方法构建。

图1 48个水生生态系统食物网案例的分布示意图
根据美国湿地分类标准[14],将48个水生食物网案例分为河口生态系统(14个)、湖泊生态系统(5个)、海洋生态系统(25个)和河流生态系统(4个)(表1)。研究的食物网案例多由牧食食物链和碎屑食物链耦合而成,即浮游植物或水生植物、碎屑等初级生产者共同驱动食物网的能量循环[15- 16]。仅包含碎屑食物链的食物网案例为湖泊生态系统Lake Findley, Mirror Lake, Marion Lake和美国南卡罗来纳州的牡蛎礁Oyster Reef(表1),这与食物网采样时间和对浮游植物和碎屑的区分方法有关。多数河口和海洋生态系统的食物网案例含有微食物环,即水中的溶解有机碳被细菌利用,细菌被原生动物(鞭毛虫、纤毛虫)摄食,原生动物进而被浮游动物摄食,浮游动物又产生溶解有机碳,形成了碳循环的环[17- 18]。总体上,研究的食物网案例数据质量较高,已得到广泛研究[19- 20]。且该48个食物网案例在拓扑特性上差异较大,拓扑特征尺度由节点数少、营养链接关系稀疏至节点丰富、营养链接紧密。均有助于本文在较大尺度上探究食物网复杂性与多样性间的普适性规律,进而验证早期提出的链接-物种缩放法则和连通度常数法则,案例在拓扑特性上差异较大,具有研究的代表性。

表1 48个水生生态系统食物网案例的特征
1.2 食物网多样性和复杂性指标
以食物网的节点数表征食物网的多样性[4- 5]。值得注意的是,这里的节点指食物网中具有相同捕食者和被捕食者的营养群(Trophic groups)和碎屑(如水体悬浮颗粒物、溶解性有机物等)[4,5,8,13]。
食物网的复杂性指标包括食物网的链接数、链接密度和连通度。食物网的链接数(Trophic links,L)即食物网的捕食关系数,这里本文不考虑TroModels数据集里食物网案例的物种迁入迁出等带来的碳流入流出,仅考虑食物网内部的营养关系:(1)对于营养群,考虑其之间的捕食或自食关系;(2)对于碎屑者,本文考虑由碎屑者流出至捕食者的关系,而对于由捕食者流入至碎屑者的关系不考虑在内[21]。链接密度(Linkage density,LD)以食物网中的链接数与物种数的比值表征,LD=L/S。连通度(Connectance, C)表示一个群落中物种之间相互作用联系的紧密强度,C=L/S2。
对不同水生生态系统食物网采用单因素分析法(one-way ANOVA)分析其物种数、链接数、链接密度、连通度是否有显著差异(P<0.05),并利用Turkey法对不同水生生态系统食物网案例进行两两比较。上述分析均通过SPSS 20.0操作。
1.3 营养缩尺模型
缩尺定律(scaling law)常用来表示物理学或化学研究中不同变量之间存在的普遍联系[9]。在食物网研究中,生态学家试图通过营养缩尺模型(Trophic scaling model)描述食物网的复杂性特征随食物网多样性的普遍规律[11],表现为复杂性指标与多样性之间的缩尺关系:
L=a1Sb1
(1)
LD=a2Sb2
(2)
C=a3Sb3
(3)
式中,L为链接数,LD为链接密度,C为连通度,S为食物网节点数,a1、a2、a3、b1、b2、b3为常数。利用Origin 2018进行模型数据拟合。
2 结果
2.1 食物网多样性和复杂性的变化特征
48个水生生态系统食物网节点数的分布范围为4—124(图2),不同的水生生态系统在食物网节点数上不存在显著性差异(ANOVA,F3,47=1.71,P=0.18)。海洋生态系统具有较高的节点数均值(46.16±32.46,平均值±标准差),其次为河口生态系统案例(33.14±27.23)和湖泊生态系统案例(32.80±38.53),河流生态系统案例的节点最少(12.25±7.76)。同时,海洋生态系统食物网案例的节点数具有最高的变异系数(1.17),河口、湖泊和河流生态系统食物网案例的节点数的变异系数近似(0.70—0.83)。
对于食物网链接数(分布范围为3—1830,图2),不同水生生态系统在食物网链接数上也不存在显著性差异(F3,47=1.17,P=0.33)。湖泊食物网案例具有较高的链接数均值(470.40±638.37),其次是海洋生态系统案例(391.67±540.41)和河口生态系统案例(238.00±417.72),河流生态系统食物网案例最低(27.5±29.01)。不同生态系统食物网案例的链接数均具有较高的变异系数(1.05—1.76)。
食物网链接密度的分布范围为0.75—15.71(图2),不同水生生态系统在食物网链接数上也不存在显著性差异(F3,47=0.91,P=0.44)。不同生态系统食物网案例的链接密度的差异特征与其链接数的差异特征近似,湖泊生态系统食物网案例的链接密度均值较高(6.79±8.04),其次为海洋生态系统案例(5.69±4.68)和河口生态系统案例(4.35±3.77),河流生态系统食物网案例最低(1.68±1.12)。湖泊生态系统食物网案例的链接密度的变异系数最高(1.18),河口、湖泊和河流生态系统食物网案例的物种丰富度的变异系数近似(0.67—0.87)。
食物网连通度的分布范围为0.06—0.25(图2),且不同水生生态系统的连通度存在显著性差异(F3,47=4.27,P=0.01)。湖泊生态系统食物网案例的连通度(0.20±0.01)显著高于河口(0.13±0.04,P=0.026)和海洋生态系统的食物网案例(0.13±0.05,P=0.013),但与河流生态系统食物网案例的连通度无显著性差别(0.17±0.06,P=0.797)。此外,湖泊生态系统食物网案例的变异系数最低,为0.04。河口、湖泊和河流生态系统食物网案例的节点数变异系数较高(0.27—0.36)。总体上,不同水生生态系统食物网案例的连通度的变异性明显低于节点数、链接数和链接密度。

图2 不同水生生态系统(河口、湖泊、海洋、河流)的食物网特征
2.2 食物网节点数与链接数的关系
基于营养缩尺模型拟合了48个实际食物网案例中节点数和链接数的相关关系(图3),具体拟合参数见表2。可以看出,整体上食物网链接数随节点数的增加呈现规律性增加趋势(R2=0.92,P<0.001)。不同水生生态系统食物网呈现相似规律,对于河口、湖泊、海洋生态系统,R2=0.92—0.99,P<0.001;对于河流生态系统,R2=0.87,P=0.039。营养缩尺模型的指数项b1为1.81—2.11。基于湖泊生态系统食物网案例的拟合的营养缩尺模型的常数a1最高,为0.46(表2)。

图3 水生生态系统食物网节点数和链接数的关系

表2 水生食物网节点数和链接数的营养缩尺模型拟合关系
2.3 食物网节点数与链接密度的关系
进一步拟合了48个食物网案例整体上及不同水生生态系统类型的节点数与链接密度关系(图4和表3)。可以看到,48个食物网案例的链接密度随食物网节点数的增加而逐渐增加(R2=0.82,P<0.001)。不同水生生态系统食物网案例也呈现类似关系,对于河口、湖泊、海洋生态系统,R2=0.77—0.99,P<0.001;对于河流生态系统,R2=0.78,P=0.036。指数b2为0.80—1.07,因此,食物网节点数和链接密度之间的关系较接近线性模型(表3)。

图4 水生食物网节点数和链接密度的关系
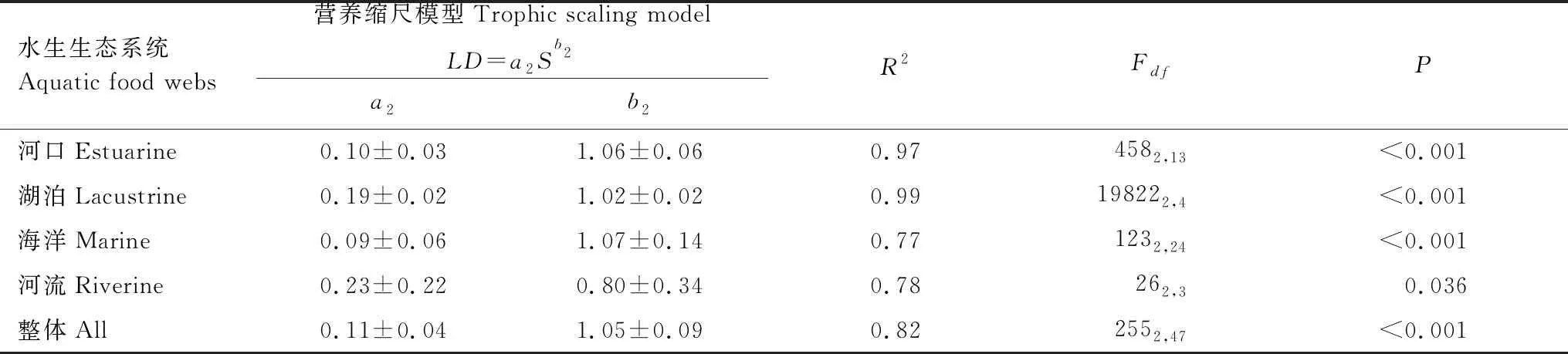
表3 水生食物网节点数和链接密度的营养缩尺模型拟合关系
2.4 食物网节点数与连通度的关系
拟合了水生生态系统食物网的节点数与连通度的关系(图5和表4)。从整体上,48个食物网案例的连通度随食物网节点数的增加而逐渐降低(R2=0.23,P<0.001)。河口、海洋、河流生态系统食物网案例呈现相似的关系,R2=0.06—0.41,P<0.001,其拟合的指数b3均为负值,为-0.32—0.12。湖泊生态系统食物网案例中连通度随节点数的变化不明显,其拟合指数b3为0.02和常数a3为0.19(表4)。

图5 水生食物网节点数和连通度的关系
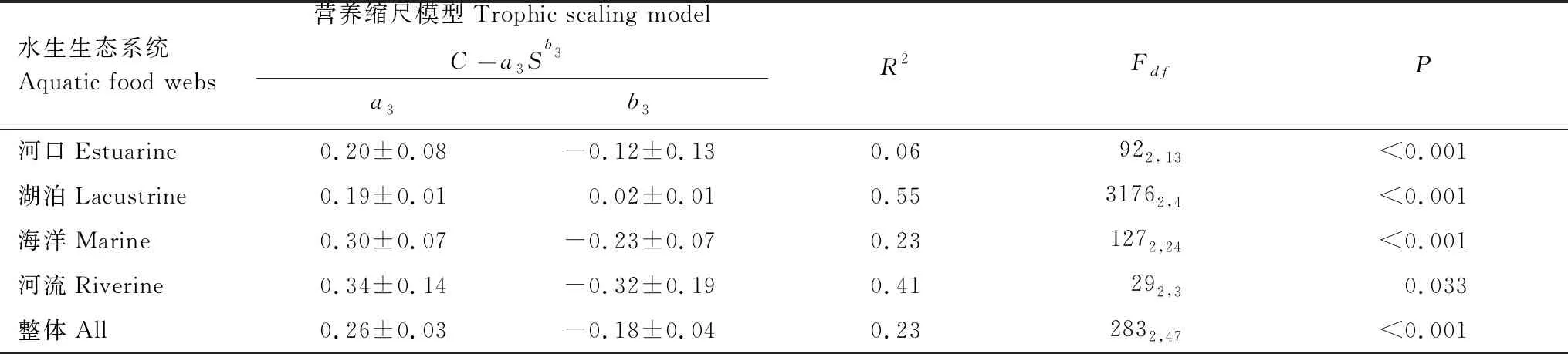
表4 水生食物网节点数和连通度的营养缩尺模型拟合关系
3 讨论
3.1 水生生态系统食物网多样性和复杂性关系分析
由于食物网是特定系统中物种间摄食关系的反映,受不同采样时间、采样地点、研究区域的大小等因素,48个食物网的节点数和链接数差异很大。如湖泊生态系统中,Lake Oneida食物网案例的物种数和链接数明显高于其他湖泊生态系统食物网案例,主要因为Lake Oneida研究区面积207 km2,而其他湖泊食物网案例的研究区范围均< 2 km2[22- 23]。相对于湖泊等系统,海洋生态系统的食物网变化动态特征更为明显,如海草床在不同季节,其水温、初级生产力、支撑草种和生物种类的组成上存在较大的差异。例如,位于佛罗里达州的墨西哥海岸的St. Marks海草床共有6个食物网案例,分布在3个相近的采样地,分别间隔100 m,涵盖两个采样季节(1月份和2月份)[24]。于2月采样的3个采样地食物网案例中节点数和链接数均高于于1月采样构建的食物网。同样地,对于河口生态系统,Neuse Estuary[25]在夏季末期构建的食物网节点数和链接数高于在夏季早期构建的食物网。具有最小节点数的食物网案例为湖泊Lake Findley,主要因为其位于华盛顿Cascade山脉针叶林中的贫营养亚高山湖,海拔1200米,生长季较短[23]。具有最多节点数的为基于干湿季节的佛罗里达最南部的海湾Florida Bay的两个案例,为热带海湾,面积广为2200 km2,具有丰富的海草和红树林,初级生产力高,为多种底栖动物和鱼类的育苗场[20]。
然而由于本研究中的食物网案例湖泊和河流生态系统的案例数较少,且每一生态系统类型的食物网案例的复杂性和多样性差异较大,分布不均匀,因此本研究中该48个食物网案例在不同水生生态系统类型间不存在节点数、链接数和链接密度的显著性差异。
尽管涵盖4种生态系统类型的48个食物网案例的复杂性和多样性差异较大,本研究致力于探究适应于不同食物网大小的复杂性和多样性的普适性关系。发现食物网多样性(节点数)与各复杂性指标(物种数、链接密度、连通度)之间的相互关系存在显著差异。其中,节点数与链接数之间的关系与节点数和链接密度之间的关系近似,即链接数、链接密度均随节点数的增加而增加,且在不同的生态系统中无显著差别。然而,节点数和连通度的关系与之相反,除湖泊生态系统外,连通度随节点数的增加而降低,湖泊生态系统的连通度随节点数的变化不明显。因此,本研究未能证明以往研究中提出的链接-物种缩放法则和连通度常数假设的普遍性,唯有湖泊生态系统食物网案例的连通度与节点数的关系近似于连通度常数假设[10]。总体上,发现的食物网节点数与复杂性之间的关系与Riede等[11]发现的现象近似, 即食物网拓扑特征与节点数之间的存在显著幂函数关系,但本研究进一步发现连通度与节点数之间幂函数关系在不同水生生态系统之间有差异。
值得考虑的是食物网的多样性是以食物网中的节点数来表征。自从Cohen发布了一个包括30个食物网的数据集后[20],节点在食物网中多是指具有相同捕食者和被捕食者的营养群或是非生态形态的碎屑物质。因此,当一个食物网具有较高的节点数时,意味着该生态系统的生物具有较高的营养多样性(Trophic diversity)[26-27],即消费者营养群的生态位较宽或不同消费者之间的生态位重叠较小[28-30],其食源具有较高的多样性。
3.2 对复杂性-稳定性悖论的调和作用
关于复杂性和稳定性之间的关系一直是生态学学者争论的焦点。1955年MacArthur提出食物网稳定性随着能量通路的增加而提高的论点[31],之后,Elton提出了生态系统越简单就越不稳定的观点[32],Odum提出食物网较大的物种多样性意味着较长的食物链、更多的共生和负反馈控制的更大可能性,更大程度上减少了波动并因此而提高了系统的稳定性[33]。然而20世纪70年代,以May为代表的理论生态学家挑战了这一观点,从数学上证明复杂性导致不稳定性[6]。近期,Jacquet等对海洋、淡水和陆地生态系统研究发现食物网的复杂性和稳定性之间没有显著性关系[34]。从反映食物网复杂性的指标来看,早期以MacArthur, Elton, Odum等为代表的生态学家多以链接数、链接密度来考虑食物网的复杂性;而May和Jacquet的研究中以连通度反映食物网的复杂性。本研究发现的随着节点数的增加,连通度呈现的与链接数和链接密度相反的关系可为调和这一悖论提供一定支撑。May和Jacquet等基于相同的复杂性指标得到的不同结论主要是由于May研究中基于随机的食物网结构,而Jacquet基于实际的食物网结构,随机的食物网结构显著降低食物网的稳定性[35-37]。
3.3 对水生生态系统保护和修复的启示
目前全球生物多样性灭绝下降的背景下,基于拓扑模拟研究发现具有较高物种丰富度或较高连通度的食物网对物种灭绝的抵抗性更强,产生更少的次生灭绝,因此该食物网具有较高的稳健性[5]。而基于动力学模拟的研究发现节点数更多的食物网对物种灭绝的抵抗能力更弱,而连通度更高的食物网对于物种灭绝的抵抗能力更强[13]。对于退化生态系统,摸清其现存食物网的多样性和潜在的食源关系,在现有基础上恢复节点之间潜在的营养链接关系,并且优先恢复邻居数(与该节点有直接链接关系的邻居数量)较高的节点,可促使其食物网的连通度增加,进而提高退化生态系统的抵抗力和稳健性。另一方面,借助基于多样性和物种相互作用之间关系的研究[38-40],恢复退化生态系统的多样性,在恢复物种多样性的同时仍需注重消费者的营养多样性,可以降低系统内物种之间的相互作用强度,促进其稳定性。
4 结论
(1)48个水生生态系统食物网案例的多样性(节点数)、复杂性(链接数、链接密度、连通度)分布特征:节点数为4—124,链接数为3—1830,链接密度为0.75—15.71,连通度为0.06—0.25。不同生态系统(河口、湖泊、海洋、河流)的节点数、链接数、链接密度均无显著性差异,但连通度存在显著差异。湖泊生态系统的连通度最高。
(2)四种水生生态系统食物网案例的链接数、链接密度均随节点数的增加而增加。
(3)河口,海洋和河流生态系统食物网案例的连通度随节点数的增加而降低,而湖泊生态系统的食物网连通度随节点数的变化不明显。
猜你喜欢
中华养生保健(2020年2期)2020-11-16
现代装饰(2020年10期)2020-10-28
科学(2020年1期)2020-08-24
短篇小说(原创版)(2017年6期)2017-08-29
小小说月刊(2017年8期)2017-08-14
短篇小说(2017年6期)2017-07-31
小学阅读指南·低年级版(2017年2期)2017-03-23
中国卫生(2016年9期)2016-11-12
肿瘤影像学(2015年3期)2015-12-09
椰城(2013年4期)2013-11-16
