
黑龙江省花培育种粳稻品种亲缘系谱及农艺性状分析
2021-07-09 12:31刘宝海高世伟门龙楠刘宇强聂守军常汇琳薛英会王洪彬
中国农学通报 2021年19期
刘宝海,高世伟,唐 铭,门龙楠,刘宇强,聂守军,刘 晴,常汇琳,马 成,薛英会,白 瑞,王洪彬
(黑龙江省农业科学院绥化分院,黑龙江绥化 152052)
0 引言
中国是世界上最大水稻生产国[1],稻米是中国约65%人口的主食[2]。黑龙江省作为国内水稻主产区之一,2017年种植粳稻面积460万hm2,约占全国粳稻种植面积50%以上[3],而新品种选育为黑龙江水稻单产提高、总产持续增加、综合生产能力稳定提升做出了突出贡献[4],对保障国家粮食安全生产和国计民生意义重大[5-6]。在新品种选育方面,花药培养是非常重要和实用的生物技术手段,也是目前获得单倍体植株和进行单倍体育种的主要方法[7]。Niizeki于1968年首次利用水稻花药离体培养分化出再生小植株,拉开了世界利用花药培养技术创新水稻新种质的序幕[8]。1975年粳稻品种‘单丰1号’成功育成,标志着中国水稻花培技术育种取得实效[9],继而全国各地开展了水稻花培育种工作,利用花药培养与杂交、诱变及转基因等育种技术相结合[10]。黑龙江省主要采用接种杂交F1代花药培养与常规系统育种相结合的水稻花培育种技术,利用其育种周期短、选择效率高、实践成果好的特点,在水稻新品种选育等方面已取得显著成效[4],据统计[11-12]1975—2019年全省已通过审定花培育种粳稻品种22个,以此类品种为直接杂交亲本共育成优良品种40个,这些品种在不同时期大面积推广应用,对促进全省水稻种植生产发挥了重要作用。
利用已育成品种系谱信息分析其亲缘关系,有助于育种家了解品种遗传背景,从而制定出更加合理的育种方案[13]。目前品种系谱分析,主要集中于某段时间品种骨干亲本[14-15]、遗传贡献[16]以及某单位[17]或某个[18]、某些性状亲缘关系[19]等方面研究。品种基因型是影响水稻花药培养力的最重要因素[20-21],杂交双亲花药培养力对杂种F1花药培养力有较大正向影响[10],理清现有花培育种粳稻品种系谱关系,可为今后花培育种工作提供有利参考,然而至今关于花培育种粳稻品种亲本系谱的梳理分析却鲜有报道。水稻品种农艺性状研究已有不少报道,中国东北地区水稻品种农艺性状各年代间有交替变化[22],从20世纪60年代以前至21世纪以后水稻生育期延长、株高降低、穗数减少、穗粒数增加、结实率和千粒重变化不大[23],不同年代品种糙米率、整精米率、垩白度、垩白粒率、直链淀粉含量、胶稠度含量、蛋白质含量等品质指标存在显著差异[24],不同时期品种农艺性状间存在相关性变化[25-28]。采用主成分及聚类方法综合评价水稻性状,能够减少单个指标的差异性影响[29],然而至今对不同年代花培育种粳稻品种农艺性状进行系统比较分析的研究报道甚少。本研究运用方差、协方差、相关、多重比较以及聚类等统计方法,重点对1975—2019年黑龙江省花培育种粳稻品种取得成效、亲缘遗传关系以及主要农艺性状进行比较分析,进而全面了解花培育种粳稻品种综合特性,筛选核心亲本种质,梳理主要问题,讨论加快推动花培育种思路方法,以期为今后黑龙江花培育种技术推广及种质资源利用、新品种创新提供理论参考。
1 材料与方法
1.1 材料
供试材料为黑龙江省1975—2019年通过花培育种方法审定的22份粳稻品种(表1)。以品种审定时间划分8个组,其中a组为2015—2019年审定的品种1~5,b组为2010—2011年审定的品种6~8,c组为2008-2009年审定的品种9~11,d组为2000—2004年审定的品种12~14,e组为1998年审定的品种15~16,f组为1992—1993年审定的品种17~18,g组为1988年审定的品种19,h组为1983年审定的品种20,i组为1975年审定的品种21~22。
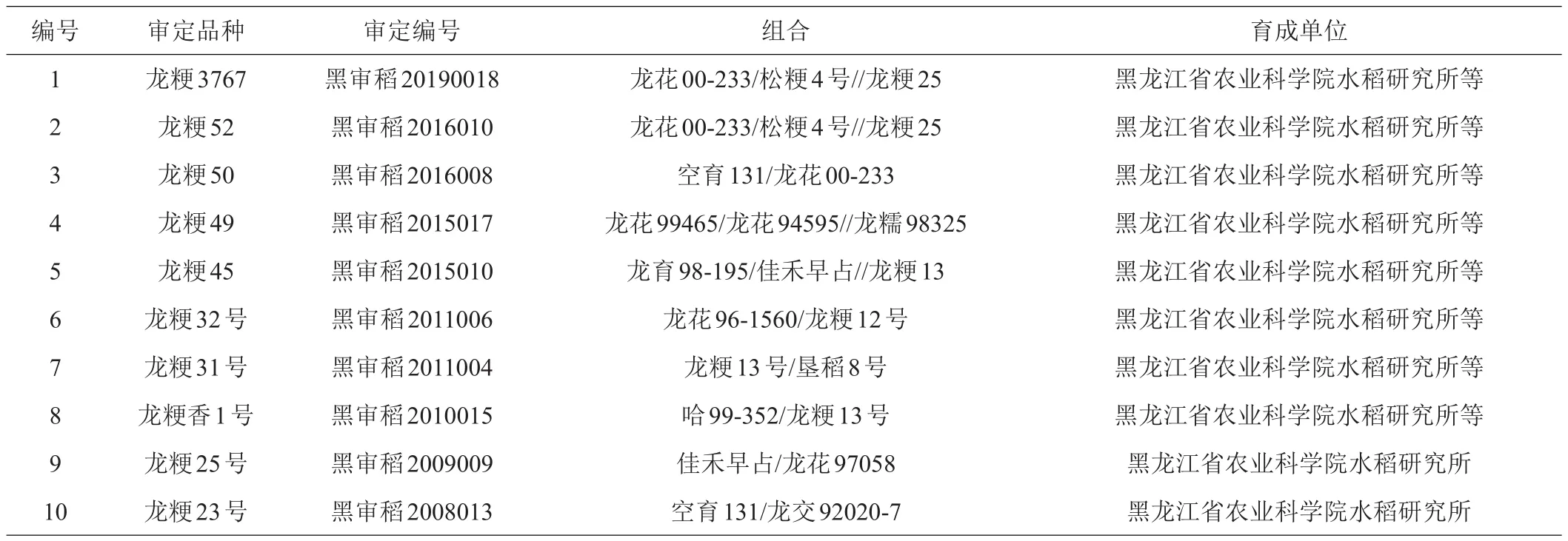
表1 1975—2019年黑龙江省粳稻花培育种品种情况
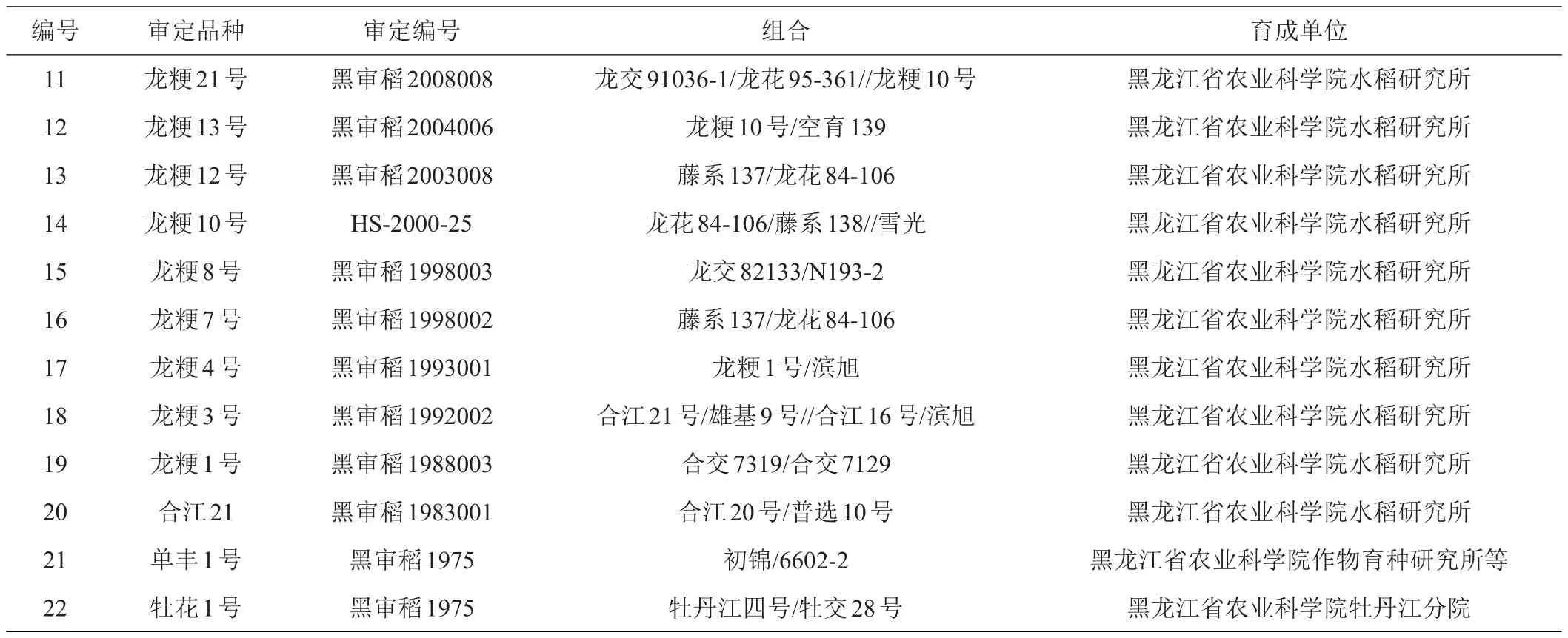
续表1
1.2 数据来源
1.2.1 品种系谱 22份粳稻品种系谱系关系及选育成效信息主要来源于《寒地粳稻育种》[4]与国家水稻数据中心[12]。
1.2.2 品种农艺性状 试验于2018—2019年在黑龙江省农业科学院绥化分院科技园区试验地进行。供试材料种子来源于黑龙江省农业科学院水稻研究所。4月中旬育秧苗床播种19个品种(‘单丰1号’、‘牡花1号’于1975年审定而没有找到种子,‘龙粳49’为糯稻,这3个品种缺失对其他品种间农艺性分析无显著影响),5月下旬单株栽插本田,每个品种4行,行长3 m,行距30 cm,株距13 cm,每穴2~3苗。3次重复。栽培管理与当地稻区一般生产大田相同。成熟期调查每小区中间2行20穗的穗数,按平均穗数取有代表性植株5株调查株高、穗长、穗粒数、千粒重、穗颈瘟级别、空壳率。同时,收获后自然风干、脱粒,测定小区实际产量,稻谷储藏1个月后委托农业部谷物及制品质量监督检验测试中心(哈尔滨)测定垩白度、垩白粒率、糙米率、整精米率、直链淀粉含量、胶稠度含量、蛋白质含量、食味品质等性状。
1.3 统计分析
品种亲缘系数(coefficient of parentage,COP)分析主要参考Emik等[30]亲缘协方差原理与范强等[31]Excel 2000宏程序VBA语言编程方法,并作了部分修改,计算各品种间COP值,多交组合以母本为编号,采用1-COP矩阵表示系谱遗传距离[32]。
笔者分析2018、2019年19个品种的产量、品质等15个农艺性状,试验各指标重复性好且变化趋一致,故主要利用2019年数据进行分析。利用IBM SPSS Statistics 25.0软件,对农艺性状数据进行多因素方差、相关以及SNK法多重比较,采用系统组间联接聚类方法,按照欧式距离测量区间,对亲缘系数矩阵和农艺性状进行聚类分析,运用Excel 2003软件绘图。
2 结果与分析
2.1 花培育种品种亲缘系数分析
22个花培育种粳稻品种亲本系谱关系梳理结果(图1)显示,这些品种的选育是不同亲本优良基因不断杂交重组累加、不良基因不断剔除的传递过程,但由于各亲本应用时期不同,其遗传贡献会有不同表现。在22个品种231个组合(表2)中有11个组合间存在亲缘关系,占组合总数的4.8%,其COP值变化范围在0.00~0.50之间,亲缘系数总和为6.50,平均值为0.31。所有组合中COP值最大为0.50,即由相同亲本育成的姊妹系品种‘龙粳3767’与‘龙粳52’,以及有共同亲本‘龙花84-106’的品种‘龙粳12号’与‘龙粳7号’。其他9个有亲缘关系组合COP值均为0.25,表现为每个品种组合都具有一个共同亲本。22个品种中,平均亲缘系数值小于或等于品种间平均值的品种占总数90.9%,其中品种间COP为0.00没有亲缘关系的6个品种占30.0%,即‘龙粳49’、‘龙粳8号’、‘龙粳4号’、‘龙粳3号’、‘龙粳1号’、‘牡花1号’。22个品种中,品种与其他品种COP平均值最高(0.04)的是‘龙粳13号’、‘龙粳7号’,说明这2个品种与其他品种间遗传亲缘关系相对较近。表明以上品种间亲缘遗传差异非常大,遗传多样性水平非常高。

图1 1975—2019年黑龙江省花培育种品种系谱图

表2 花培育种品种间亲缘系数值

续表2
2.2 花培育种品种亲缘聚类分析
对花培育种品种间的COP值进行聚类分析,在欧氏距离24时可聚类为Ⅰ、Ⅱ、Ⅲ共3个类群(图2)。Ⅰ类群共有2个品种,包括‘龙粳3767’、‘龙粳52’。该类群品种都是由同一个单位主导选育而成,分别在2016、2019年育成审定,类群内2个品种是由相同亲本杂交花培育种选育而成,属于姊妹系,都是具有花培育种材料‘龙花00-233’和‘龙粳25号’的后代,‘龙花00-233’作为中间育种材料具有早熟、丰产、抗病、优质、株型好等优良特性[33]。Ⅱ类群品种数最多,共有17个品种,占品种总数的77.3%,其中有15个品种是由同一个单位主导或独家选育而成,占类群内品种数的88.2%。该类群品种1975—2016年间育成,时间跨度最大,亲本类型也最丰富,以花培育种后代材料为亲本育成品种11个,占品种数的64.7%。根据品种间亲缘系数关系,在欧氏距离22时可划分为5个亚类群,Ⅱ-1亚类群包括‘龙粳49’、‘龙粳8号’、‘龙粳4号’、‘龙粳3号’、‘牡花1号’、‘龙粳1号’6个品种,Ⅱ-2亚类群包括‘单丰1号’、‘合江21’,Ⅱ-3亚类群包括‘龙粳23号’、‘龙粳50’,Ⅱ-4亚类群包括‘龙粳32号’、‘龙粳13号’、‘龙粳21号’,Ⅱ-5亚类群包括‘龙粳31号’、‘龙粳45’、‘龙粳25号’、‘龙粳香1号’4个品种。Ⅲ类群共有3个品种,包括‘龙粳12号’、‘龙粳7号’、‘龙粳10号’。该类群品种由同一个单位独家选育而成,分别在2003、1998、2000年育成审定,且以花培育种后代材料‘龙花84-106’为亲本选育而成。
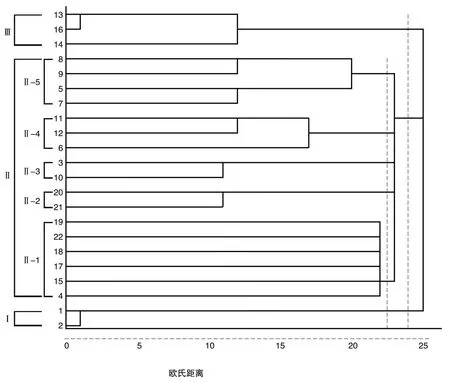
图2 基于1-COP值的花培育种品种聚类
2.3 花培育种品种农艺性状遗传多样性分析
以亲缘系数Ⅰ、Ⅱ、Ⅲ聚类品种划分,对19个品种15个农艺性状(表3)遗传多样性分析结果如表4A,仅穗粒数和蛋白质含量2个性状遗传方差P值为0.045、0.017达差异显著水平,约占全部性状13.3%,表明以亲缘系数聚类划分的品种间农艺性状差异没有充分呈现,有必要做进一步分析。以品种审定时间划分,对8个组19个品种15个农艺性状遗传多样性分析结果如表4B,有6个农艺性状遗传差异达显著水平,约占全部性状40.0%,其中产量、胶稠度含量、垩白度遗传方差P值为0.000、0.006、0.006达极显著差异水平,穗粒数、糙米率、食味品质遗传方差P值为0.046、0.014、0.039达显著差异水平。由偏Eta平方(η2)值大小[34]可知,农艺性状对品种影响效应由大到小为产量(η2=0.870)>胶稠度含量(η2=0.780)>垩白度(η2=0.779)>糙米率(η2=0.740)>食味品质(η2=0.675)>穗粒数(η2=0.664)>穗颈瘟级别(η2=0.638)>直链淀粉含量(η2=0.627)>蛋白质含量(η2=0.617)>穗长(η2=0.569)>垩白粒率(η2=0.506)>空壳率(η2=0.477)>株高(η2=0.345)>整精米率(η2=0.302)>千粒重(η2=0.275)。说明不同时期育成品种的农艺性状间遗传变异程度表现不同,呈现出受产量遗传变异影响最大,其次是胶稠度含量,第三是垩白度,影响最小的性状是千粒重。

表3 花培育种品种农艺性状平均值
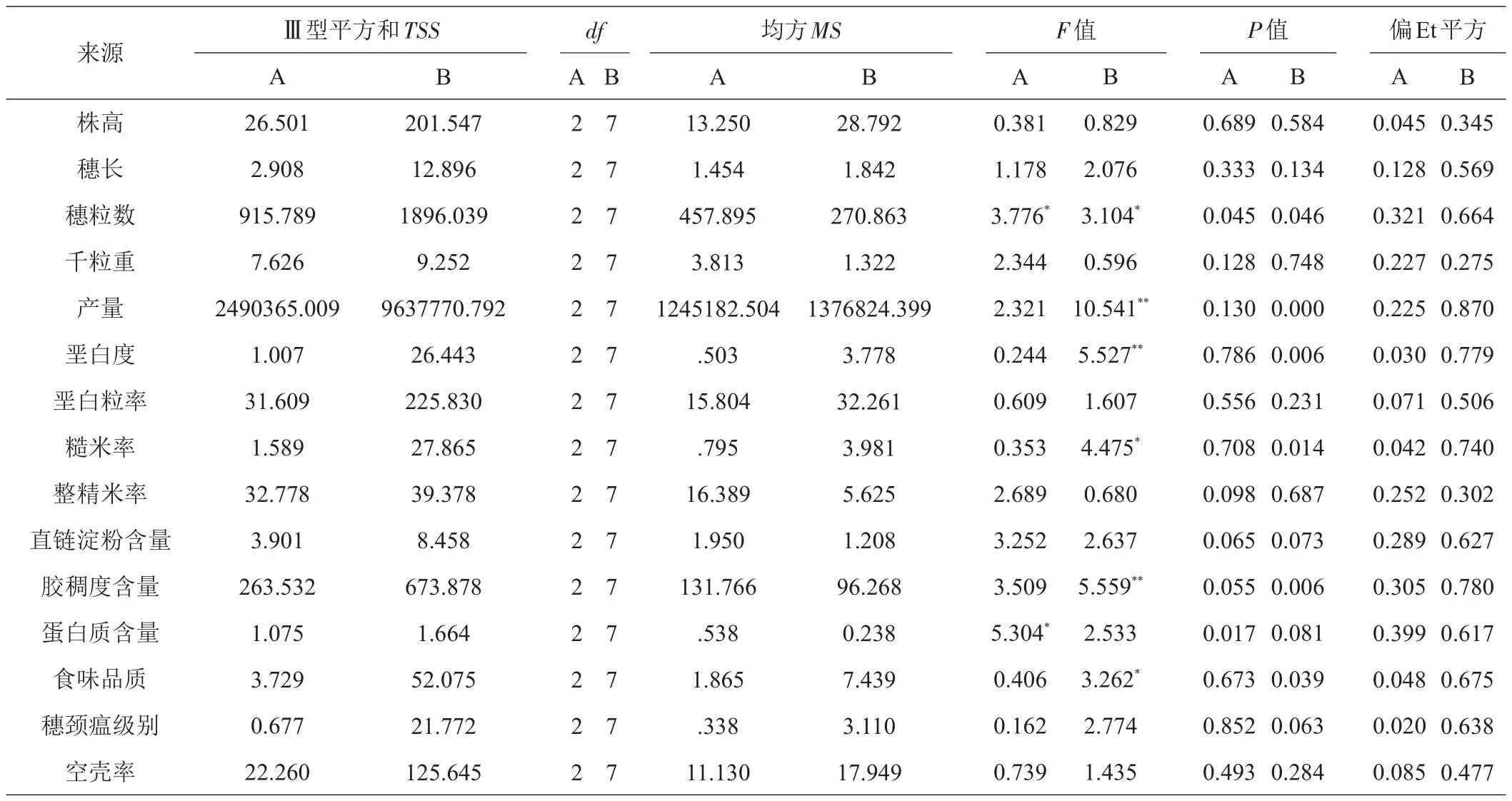
表4 花培育种品种农艺性状遗传多样性
对以品种审定时间划分的遗传变异达显著水平农艺性状进行多重比较分析(图3)表明,产量a组显著高于h、g、f、d、e、b组,但与c组差异不显著;b组显著高于h、g、f组,但与d、e、c组差异不显著;d、e、f、g、h组间差异不显著,表明花培育种品种产量性状由h至a组遗传增益显著,且近10年增益更加明显。穗粒数a组显著多于其他7组;d、e、c、b、f、g组间差异不显著,但显著多于h组;h组最少,但与d、e、c、f差异不显著,表明穗粒数近5年增加显著,而其他时间段里相对较稳定。垩白度b组最小,与d、c组差异不显著,与其他组差异显著;h组最大,与g、f组间差异不显著,但与其他组差异显著;b、d、c组间差异不显著,表明h至a组品种垩白度整体呈下降趋势,但1998年以来波动较大。垩白粒率b组最小,与h组差异不显著,与其他组差异显著;除b组外其他组间不显著,表明花培育种过程中垩白粒率变化幅度相对较小,且无明显规律。胶稠度含量a、b、c组间差异不显著,但显著高于e、f、g、h组;a、b、c、d组间差异不显著,d、e、f、g、h组间差异不显著,表明h至a组品种胶稠度含量呈逐渐上升趋势,d组成为胶稠度含量发生显著变化的过度时期。食味品质a、b组显著高于e、f、g、h组,a、b、c、d组间差异不显著,c、d、e、f、g、h组间差异不显著,表明h至a组品种食味品质呈逐渐增加趋势,c、d组为显著提高的过度时期。由此可知,供试品种农艺性状遗传变异程度不同,η2>0.664且变异显著的6个性状h至a组变化规律有差别,但总体呈现向好趋势,这是优良资源基因累加与科研选择改良共同作用的结果。

2.4 花培育种品种农艺性状相关性分析
对19个品种15个农艺性状的相关性分析(表5)显示,相关系数达显著水平有16个,仅约占105个相关关系的1.1%。遗传差异达显著水平的6个农艺性状与其他9个性状间达到显著相关水平的有产量与蛋白质含量正相关,胶稠度含量与垩白粒率负相关、与蛋白质含量正相关,垩白度与蛋白质含量负相关、与垩白粒率正相关,食味品质与株高、蛋白质含量负相关,蛋白质含量与垩白粒率负相关;9个性状间达到显著相关水平的有整精米率与糙米率正相关、与千粒重负相关,蛋白质含量与株高正相关,穗颈瘟级别与千粒重正相关。同时空壳率、直链淀粉含量、穗长均与其他性状无显著相关性。其中,产量与胶稠度含量、食味品质、蛋白质含量相关系数为0.725、-0.656、0.661,达极显著水平,与穗粒数相关系数为0.547,达显著水平;胶稠度含量与垩白粒率呈极显著负相关,其系数为-0.592;蛋白质含量与垩白度呈极显著负相关,其系数为-0.663,与垩白粒率呈显著负相关,其系数为-0.538,食味品质与胶稠度含量呈极显著负相关,其系数为-0.635,与株高呈显著负相关,其系数为-0.470,表明在一定范围内,产量越高,胶稠度含量、蛋白质含量越大,穗粒数越多,而食味品质会越差,胶稠度含量越大而垩白粒率会越小,蛋白质含量越大而垩白度、垩白粒率会越小,食味品质越高而胶稠度含量会越小。垩白粒率与垩白度相关系数为0.864,其值最大且呈极显著正相关。整精米率与千粒重呈极显著负相关,其系数为-0.612,说明千粒重越大,整精米率越低。整精米率与糙米率相关系数为0.481呈显著正相关。蛋白质与胶稠度含量相关系数为0.7177呈极显著正相关,与食味品质相关系数为-0.590呈显著负相关,与株高相关系数为0.479呈显著正相关,表明蛋白质含量升高食味品质会下降,株高增加蛋白质含量会增加。穗颈瘟级别与千粒重相关系数为0.469呈显著正相关,表明穗颈瘟级别与千粒重具有同向变化趋势。空壳率、直链淀粉含量、穗长均与其他性状无显著相关性,表明这3个遗传性状独立性较强。由此可知,黑龙江花培育种品种农艺性状间相关性达显著水平所占比例较少,性状间相互影响相对较小。
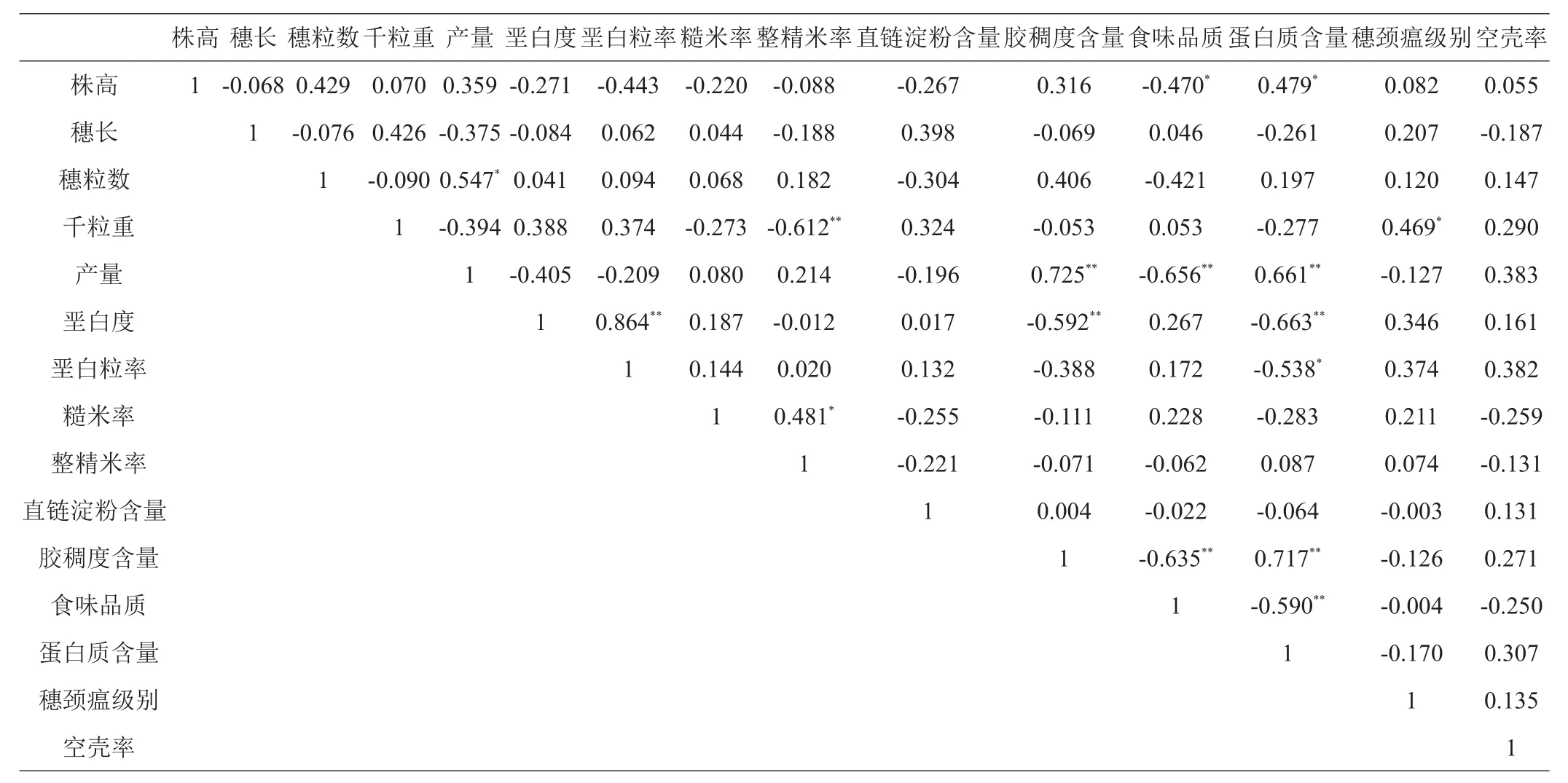
表5 花培育种品种农艺性状相关系数
2.5 花培育种品种农艺性状聚类分析
对19个品种15个农艺性状进行聚类,在遗传距离12时可聚类为Ⅰ、Ⅱ、Ⅲ共3个类群(图4),对各类群品种农艺性状遗传变异分析如表6。

表6 基于农艺性状聚类的花培育种品种遗传变异系数

图4 基于农艺性状值的花培育种品种聚类
Ⅰ类群共有4个品种,包括‘龙粳香1号’、‘龙粳49’、‘龙粳50’、‘龙粳52’。该类群品种于2010—2016年育成,农艺性状变异系数平均值为17.8%,其中垩白度变异系数最大为69.7%,其次是垩白粒率为58.2%,第三位是穗颈瘟级别为54.4%,变异系数最小的3位依次为糙米率(1.1%)、食味品质(1.2%)和整精米率、胶稠度含量(2.4%),说明该类群品种外观品质垩白度、穗颈瘟级别级别变化较大,而加工品质糙米率、整精米率及理化品质胶稠度含量、食味品质变化较小。Ⅱ类群共有4个品种,包括‘龙粳1号’、‘龙粳3号’、‘龙粳4号’、‘龙粳7号’。该类群品种于1988—1998年育成,农艺性状变异系数平均值为9.2%,其中变异系数较大的前3位依次为空壳率(30.5%)、穗颈瘟级别(28.6%)、垩白度(18.4%),变异系数较小的末3位依次为胶稠度含量(0.9%)、糙米率(0.9%)、整精米率(0.9%)和食味品质(1.6%)、蛋白质含量(2.0%),说明该类群品种空壳率、穗颈瘟级别及垩白度遗传变异较大,胶稠度含量、糙米率、整精米率、蛋白质含量和食味品质变化较小。Ⅲ类群包括‘龙粳10号’、‘龙粳21号’、‘龙粳8号’等11个品种,品种数最多,约占总数57.9%,选育时间为1998—2019年,时间跨度也最大。该类群品种农艺性变异系数平均值为23.0%,其中垩白度为126.9%,是3个类群最大值,其次是垩白粒率(64.2%),第三位是穗颈瘟级别(63.6%),变异系数较小的性状依次为整精米率(2.2%)、蛋白质含量(2.2%)和直链淀粉含量(3.5%)、食味品质(4.2%),说明该类群品种垩白及穗颈瘟级别变化幅度大,而食味品质和整精米率、蛋白质含量、直链淀粉含量变化较小。
结合表3~4可知,Ⅰ类群中,产量、胶稠度含量、糙米率、食味品质较高及垩白度、垩白粒率、穗颈瘟级别较低等综合性状优良的前2位品种为‘龙粳香1号’和‘龙粳49’;Ⅱ类群中,产量、胶稠度含量、糙米率、食味品质较高及垩白度、空壳率、穗颈瘟级别较低等综合性状优良的前4位优良品种为‘龙粳45’、‘龙粳32号’、‘龙粳3767’和‘龙粳31号’;Ⅲ类群中,产量、胶稠度含量、糙米率、食味品质较高及垩白度、垩白粒率、穗颈瘟级别较低等综合性状优良前2位优良品种为‘龙粳7号’和‘龙粳4号’。
3 讨论
3.1 花培育种技术的推广与应用
单倍体育种具有加速杂种后代纯合、缩短育种周期、提高选择效率等优点,受到育种专家广泛重视[35]。自1964年印度学者Guha在曼陀罗花药培养中获得单倍体植株以来,迄今为止已在200多种植物中获得成功[36]。以小麦、水稻等作物花药培养为代表的花培育种技术已成为生物技术应用于育种实践的重要组成部分[37]。在本研究中,黑龙江省利用花培育种技术应用成效显著,育成的22个粳稻品种及以其为亲本育成的60个品种在各时期水稻育种与生产过程中发挥了重要作用,如2011年审定的‘龙粳31号’,创造了2012—2016年连续5年种植推广面积全国最大的纪录。但是,除1975年育成‘单丰1号’、‘牡花1号’外的其他20个品种均由一家单位主导育成,这与近10年来30余家科研院所、大学及企业从事并成功审定水稻品种[12]比较,花培育种技术推广与应用任重道远。花培方式育成粳稻品种分别约占1975、1990、2000、2010、2015年以来 419、393、349、247、161个品种总数的 5.3%、4.6%、4.0%、4.0%和2.5%,呈下降趋势,表明该技术的实践应用效果还有待进一步挖掘。同时,针对不同寒地粳稻基因型杂交亲本花药培养力差异大且较低、单倍体加倍率也较低的制约因素,还需在生理生化、分子机制等方面进一步探索。
3.2 花培育种品种资源的筛选与利用
农作物种质资源是品种遗传改良的重要物质基础[38],遗传多样性是水稻种质资源评价和利用的重要研究内容[39],了解作物表型变异遗传基础,筛选、分析和发掘具有优良性状的种质资源,能够帮助人们有效利用现有的遗传多样性资源提高育种效率以及促进作物产量和品质改良[40-42]。在本研究中,80%的花培育种粳稻品种是以花培育种材料为杂交亲本育成,231个品种组合中仅有11个组合存在亲缘关系,约占4.8%,COP小于均值约占90.9%,这些品种间遗传差异非常大,遗传多样性水平非常高,与李红宇等[16]研究黑龙江水稻品种遗传基础相对狭窄结果有所不同,因此明确花培育种资源系谱与亲缘特点对其筛选与利用是非常必要的。亲缘系数和农艺性状值在欧氏距离24与12时,可分别聚类为3个类群,虽然2种聚类结果不同,但可以相互参考、综合评价,按照在不同农艺性状类群中筛选种质资源原则,结合影响品种综合优良特性主要性状因素,本着品种亲缘系数遗传距离聚类较远而利于用作杂交亲本的思路,筛选出‘龙粳香1号’、‘龙粳45’、‘龙粳3767’、‘龙粳7号’和‘龙粳4号’5份种质资源,作为花培育种的核心亲本种质加以利用。同时,针对花培育种粳稻品种分子标记亲缘系谱的研究有待进一步探索。
3.3 花培育种农艺性状的育种目标
高产、优质、多抗是水稻育种的永恒主题[43]。水稻性状之间关系复杂,相互制约与促进并存[44]。如何在育种中协调好这些关系,是需要深入分析和探讨的重要问题[45],也是提高育种效率的必然选择[46]。在本研究中,8组19个品种15个农艺性状遗传变异程度与王海泽等[22]研究结果有所不同,但均呈现遗传改良向好趋势,这应该是多年来对高产、优质、多抗育种目标追求与坚持的结果,其中有6个性状遗传变异达显著水平,且遗传累加增益最大的性状是产量,其次是胶稠度含量,第三位是垩白度,第四位是糙米率,第五位是食味品质,穗颈瘟级别、直链淀粉含量、蛋白质含量、穗长、垩白粒率、空壳率、株高、整精米率、千粒重等9个性状遗传差异达不显著水平,这些性状应该是今后育种要重视和不断改良的目标。同时,农艺性状间相关系数达显著水平仅有16个,包括6个差异显著水平性状与其他9个性状间以及9个性状之间的显著水平相关,约占105个相关关系1.1%,与周健等[23]研究农艺性状间多呈显著正相关结果不同,表明现有花培育种粳稻农艺性状间既有较大遗传差异,也有较强独立性和一定的关联性。今后黑龙江花培育种粳稻目标应该是,在稳定改善产量、胶稠度含量、穗粒数、糙米率、食味品质及垩白度等6个农艺性状基础上,着重提高结实率、整精米率、穗长,降低株高、垩白粒率、蛋白质含量、直链淀粉含量、穗颈瘟级别,稳定千粒重。同时,如何综合考虑种植、加工和消费者等各经济主体对品种农艺性状的需求,采用定性与定量相结合的多育种目标优化设计研究有待进一步探索。
猜你喜欢
粮油食品科技(2022年5期)2022-09-27
今日农业(2021年15期)2021-10-14
纺织高校基础科学学报(2021年2期)2021-07-13
粮油食品科技(2020年6期)2020-12-08
农民致富之友(2020年32期)2020-12-03
红楼梦学刊(2020年2期)2020-02-06
现代装饰(2018年4期)2018-05-22
河南农业(2017年7期)2017-07-25
纺织检测与标准(2016年3期)2016-08-16
浙江农业科学(2016年11期)2016-05-04
