
湘西两种花色野生巨紫荆光合作用日变化规律
2021-07-09 03:09和红晓
经济林研究 2021年2期
和红晓,杨 艳,黄 斌
(1.湘西土家族苗族自治州林业局,湖南 吉首 416000;2.湖南省林业科学院,湖南 长沙 414000)
湘西野生巨紫荆Cercis gigantea为豆科紫荆属落叶大乔木,是我国特有的珍稀乡土树种。该树种在我国的分布较少,其高大乔木及野生群落更是罕见。其树体高大,生长速度快,适应能力强,初春先花后叶,无论枝与干都布满紫红花,花簇繁茂,嫣红夺目,艳丽可爱,叶片心形,圆润而有光泽,光影相互掩映,颇为动人,是优良的观赏、材用、蜜源兼用树种;巨紫荆对氯气有一定的抗性,滞留尘埃的能力强,耐旱、耐寒、耐水渍、耐贫瘠,具有豆科植物所特有的根瘤菌,能提高土壤地力,改善生境条件,是生态修复先锋树种;巨紫荆根、叶、皮均可入药,具有活血、化淤、消肿的功效,有很高的药用价值,是理想的经济药用植物。综合而言,巨紫荆具有重要的观赏、经济、生态和独特的开发利用及研究价值,是当前乡村振兴及生态文明建设不可多得的特色植物资源,具有广阔的开发利用前景[1-7]。
光合作用是植物的重要生理活动,是影响果实品质及产量的重要因素。在自然条件下,植物光合作用的日变化曲线有两种典型的类型:一种是单峰型;另一种是双峰型。光合作用的强度受植物种类、光照、温度、环境中CO2浓度、水、矿质元素等内外因素的影响[8-10]。柯世省等[11]指出,直接影响紫荆净光合速率日变化规律的主要因子分别有光合有效辐射、空气相对湿度和空气CO2浓度,其影响程度的大小顺序为:空气CO2浓度<空气相对湿度<光合有效辐射。陈模舜等[12]对紫荆连体叶片的光合日变化及光响应情况进行了研究,他们指出,其净光合速率(Pn)的日变化曲线表现为双峰型,其胞间CO2浓度(Ci)的日変化趋势与其净光合速率的日変化趋势基本相反,中午其Ci增高,说明此时净光合速率的降低主要受到非气孔限制因素的影响。而紫荆的暗呼吸速率日变化曲线则表现为单峰型,中午其测定值最高,表明光合“午休”现象产生的重要原因是长时间的强光照射造成的光抑制作用。目前,我国学者针对巨紫荆的相关研究工作虽已启动,但相关研究多集中在巨紫荆种子特性和栽培管理及其在园林绿化中的应用等方面,而关于野生巨紫荆光合作用日変化规律的研究未见报道。为给野生巨紫荆的引种驯化、苗木培育和高效栽培提供科学理论依据,本研究对自然条件下巨紫荆光合作用及其环境因子的日变化情况进行了观测,分析了红花和粉白花巨紫荆的光合特性的日变化规律及其与生理和环境因子的关系。
1 试验地概况
供试的野生巨紫荆生长于湖南省湘西土家族苗族自治州永顺县高坪乡西米村三支箭山,位于永顺县羊峰山东南40 km 处,地处北中纬度,最高海拔为923.9 m。属中亚热带山地季风性湿润气候,光热充足,四季分明,平均气温16.2 ℃,平均日照时数1 306 h,年均降水量1 327 mm,无霜期216~265 d,年均冰冻期在20 d 以上。土壤为石灰质黄壤,岩石多,土层薄,其pH 值为5.5~6.1,呈酸性[1]。
2 试验材料和试验方法
2.1 试验材料
在试验样地中,选择生长于自然条件下、胸径均为16 cm 的野生红花巨紫荆、粉白花巨紫荆树冠中部枝条上生长一致、无病虫害、向阳面的叶片作为试验材料。
2.2 试验方法
2020年9月,选择晴朗天气的8:00—18:00 时,采用Li-6400 便携式光合作用测定仪(美国LICOR 公司生产),在自然条件下,选择试验样地中平均木的冠中部向阳面枝条上的叶片进行光合作用的测定,每隔2 h 测定1 次,每株测定5 片叶子,每个叶片各测定1 次,取其平均值。
测定参数分别为净光合速率(Net photosynthesis rate,μmol·m2s-1)、气孔导度(Stomatal conductivity,μmol·m2s-1)、蒸腾速率(Transpiration rate,mmol·m2s-1)、胞间CO2浓度(Intercellular CO2concentration,μmol·mol-1),同时记录光照强度(Photosynthetically active radiation,μmol·m2s-1)、气温(Air temperature,℃)、叶片温度(Temperature of leaf,℃)、环境中的CO2浓度(Air CO2concentration,μmol·mol-1)、空气相对湿度(Relatively humidity of air,%)等参数;按照以下公式计算水分利用率(Water use rate,μmol·mmol-1)和气孔限制值(Stomatal limitation value,μmol·mol-1):
水分利用率=净光合速率/蒸腾速率;
气孔限制值=(环境中的CO2浓度-胞间CO2浓度) / 环境中的CO2浓度。
2.3 数据处理
运用Excel 2010 软件进行数据统计和制图,采用SPSS 23.0 软件进行相关性分析及双侧显著性检验。
3 结果与分析
3.1 光合特性日变化
3.1.1 净光合速率的日变化规律
两种花色野生巨紫荆叶片净光合速率的日变化曲线如图1所示。由图1可知,不同花色野生巨紫荆叶片净光合速率的日变化曲线均为双峰曲线,其首个峰值均大于其次峰值,净光合速率最大值出现在10:00 时前后,可能因为此时的光照、空气温湿度等环境因子比较适宜;“午休”现象比较明显,出现在12:00 时左右。红花和粉白花巨紫荆叶片净光合速率的差别较大,红花巨紫荆的净光合速率最大值为7.81 μmol·m2s-1,粉白花巨紫荆的净光合速率最大值为5.17 μmol·m2s-1,后者仅为前者的66.1%;当其“午休”时,红花巨紫荆的净光合速率为4.64 μmol·m2s-1,粉白花巨紫荆的净光合速率为1.28 μmol·m2s-1,后者仅为前者的27.5%。红花巨紫荆的净光合速率日均值为4.47 μmol·m2s-1,粉白花巨紫荆的净光合速率日均值为2.59 μmol·m2s-1,前者是后者的1.72 倍。

图1 净光合速率的日变化曲线Fig.1 Diurnal change of net photosynthesis rate
3.1.2 气孔导度的日変化规律
两种花色野生巨紫荆气孔导度的日变化曲线如图2所示。由图2可知,两种花色巨紫荆气孔导度的日变化曲线也都为双峰曲线,其“午休”现象明显,其首峰值均大于其次峰值,其日最大峰值均出现在10:00 时左右,“午休”现象均出现在12:00 时,但红花巨紫荆的气孔导度明显高于粉白花巨紫荆的气孔导度。气孔导度第1 个峰值,红花巨紫荆的为0.22 μmol·m-2s-1,粉白花巨紫荆的为0.13 μmol·m-2s-1,后者仅为前者的56.6%;“午休”时的气孔导度,红花巨紫荆的为0.15 μmol·m-2s-1,粉白花巨紫荆的为0.06 μmol·m-2s-1,后者仅为前者的40.4%。气孔导度的日变化曲线和净光合速率与光合速率日变化曲线均基本一致,说明其相关性均较密切。
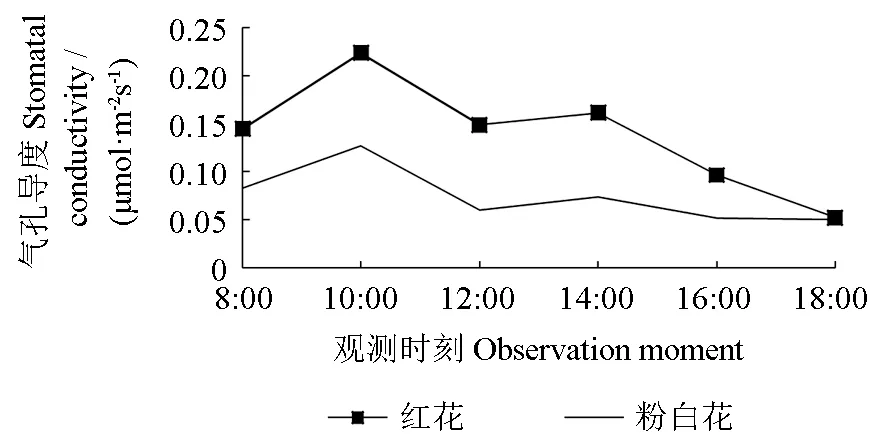
图2 气孔导度的日变化曲线Fig.2 Diurnal change of stomatal conductivity
3.1.3 胞间CO2 浓度的日変化规律
两种花色野生巨紫荆胞间CO2浓度的日变化曲线如图3所示。由图3可知,巨紫荆的胞间CO2浓度,8:00—10:00 时逐步下降,12:00 时后略有回升,之后又逐步下降,16:00 时后又开始逐步回升。早上巨紫荆的胞间CO2浓度最大,红花的为356.57 μmol·mol-1,粉白花的为369.78 μmol·mol-1;其原因可能是,随着光合作用的增强,胞间CO2浓度逐渐下降,而早上环境中的CO2浓度较高。至12:00 时,巨紫荆的胞间CO2浓度缓慢回升,14:00 时降为最低值,红花与粉白花的分别为299.17 和282.15 μmol·mol-1;之后,随着光照强度的逐渐减弱,胞间CO2浓度逐渐上升,至18:00 时,红花与粉白花的胞间CO2浓度分别升至336.40 和329.51 μmol·mol-1,均较接近于早上水平。红花巨紫荆和粉白花巨紫荆胞间CO2浓度的差别不太明显。

图3 胞间CO2 浓度的日变化曲线Fig.3 Diurnal change of intercellular CO2 concentration
3.1.4 蒸腾速率的日変化规律
两种花色野生巨紫荆蒸腾速率的日変化曲线如图4所示。由图4可知,两者均为双峰曲线,且与其净光合速率的变化趋势均基本一致。红花和粉白花巨紫荆的蒸腾速率,从早上开始均逐渐上升,10:00 时分别升至3.56 和1.78 mmol·m-2s-1,均出现了第1 个峰值;然后随着净光合速率的下降,其蒸腾速率也均随之下降,12:00 时后均又开始上升,14:00 时均出现了第2 个峰值,且均达到了全天的最大值,红花和粉白花的分别为4.44 和2.65 mmol·m-2s-1,与其第1 个峰值相比,分别大0.88和0.87 mmol·m-2s-1;之后随着光照强度的降低而逐渐下降,18:00 时分别降为 0.9 和0.8 mmol·m-2s-1。巨紫荆的蒸腾速率,红花的明显高于粉白花的;当蒸腾速率达到最大峰值时,粉白花的仅为红花的59.68%。

图4 蒸腾速率的日变化曲线Fig.4 Diurnal change of transpiration rate
3.1.5 叶片温度的日変化规律
两种花色野生巨紫荆叶片温度的日变化曲线如图5所示。由图5可知,两者叶片温度的日变化曲线均为单峰曲线。红花巨紫荆叶片温度的日变化范围为27.35~35.37 ℃,早上最低,随后开始逐步上升,最高值出现在14:00 时左右,接着又逐渐降低;粉白花巨紫荆叶片温度的日变化范围为26.51~36.48 ℃,早上最低,之后逐步上升,最高值出现在14:00 时,随后又逐渐降低。红花和粉白花巨紫荆的最高叶片温度和最低叶片温度之差分别为-0.83 和1.12 ℃,说明其叶片温度的变化均不大。

图5 巨紫荆叶片温度的日变化曲线Fig.5 Diurnal changes of temperature of leaf
3.1.6 水分利用率的日変化规律
两种花色野生巨紫荆水分利用效率的日变化曲线如图6所示。由图6可知,红花巨紫荆的水分利用效率,从早上开始逐步降低,最低值(1.14 μmol·mmol-1)出现在16:00 时,之后逐步回升;粉白花巨紫荆的水分利用效率,10:00 时后迅速下降,12:00 时左右降至最低值(0.73 μmol·mmol-1)后又开始回升。红花巨紫荆的水分利用效率大部分时间一直处于较高水平上,且其水分利用率的最低值比粉白花巨紫荆的最低值高出56.16%。

图6 水分利用效率的日变化曲线Fig.6 Diurnal changes of water use rate
3.1.7 气孔限制值的日変化规律
两种花色野生巨紫荆气孔限制值的日变化曲线如图7所示。由图7可知,两者气孔限制值的日变化曲线均呈双峰曲线,其首峰值均低于其次峰值,12:00 时均出现了明显的“午休”现象;红花巨紫荆的气孔限制值在14:00 时达到了最高值(0.22 μmol·mol-1),粉白花的气孔限制值在16:00时达到了最高值(0.26 μmol·mol-1)。这一结果说明,巨紫荆叶片的气孔效应在中午的表现较为明显,这可能是其“午休”现象产生的主要原因之一。

图7 气孔限制值的日变化曲线Fig.7 Diurnal changes of stomatal limitation value
3.2 环境因子的日変化规律
两种花色野生巨紫荆所在环境中的光照强度、大气中的CO2浓度、水气压差、气温、相对湿度的日变化曲线分别如图8、9、10、11、12 所示。由图8可知,两者所在环境中的光照强度的日变化曲线均为单峰曲线;8:00 时后光照强度迅速上升,红花巨紫荆的光照强度在10:00 时达到最大值(822.47 μmol·m-2s-1);粉白花巨紫荆的光照强度在12:00 时达到最大值(686.38 μmol·m-2s-1),之后逐渐下降。由图9可知,大气中的CO2浓度,从8:00 时开始逐步下降,至16:00 时降至最低值后又开始回升;红花和粉白花巨紫荆所在环境中CO2浓度的最低值分别为384.24 与383.61 μmol·mol-1。红花和粉白花巨紫荆所在环境中的水气压差(图10),从8:00 时至14:00 时,分别逐步上升至最大值2.76 与3.30 kPa 后逐步下降。红花和粉白花巨紫荆所在环境中的气温(图11),从早上开始上升至最大值后逐步下降;红花巨紫荆的最高气温(36.00 ℃)出现在16:00 时,而粉白花巨紫荆的最高气温(36.16 ℃)出现在14:00 时。红花和粉白花巨紫荆所在环境中的相对湿度(图12),从8:00 时至午后逐步下降,降至最低值后又开始逐步回升;红花巨紫荆相对湿度的最低值(43.69%)出现在16:00 时,而粉白花巨紫荆相对湿度的最低值(42.02%)出现在14:00时;相对湿度的变化趋势与气温的变化趋势正好相反,早上最高,然后随着气温的上升逐渐下降,降至最低值后又逐步上升,红花和粉白花巨紫荆的相对湿度的变化范围分别为43.69%~70.85%与42.02%~73.35%。

图8 光照强度的日变化曲线Fig.8 Diurnal changes of photosynthetically active radiation

图9 大气中CO2 浓度的日变化曲线Fig.9 Diurnal changes of air CO2 concentration

图10 水气压差的日变化曲线Fig.10 Diurnal changes of vapor pressure deficit
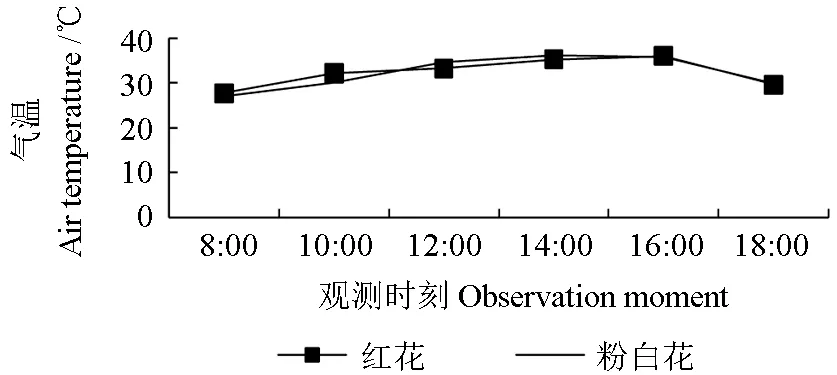
图11 气温的日变化曲线Fig.11 Diurnal changes of air temperature

图12 相对湿度的日变化曲线Fig.12 Diurnal changes of relatively humidity of air
3.3 巨紫荆的光合速率与其生理、环境因子间的相关性分析
为了探明光合作用与环境因子间的相互关系,对红花与粉白花巨紫荆的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、气温、大气中的CO2浓度、水气压差、相对湿度、叶片温度和光照强度之间的相关性进行了分析,结果分别见表1和2。

表1红花巨紫荆的净光合速率与各影响因子间的相关系数†Table 1 Correlations of photosynthetic rate and other factors of C.gigantea(red flower)?† “**”表示极显著相关(P<0.01),“*”表示显著相关(P<0.05)。“**”Correlation is significant at the 0.01 level,“*” Correlation is significant at the 0.05 level.

分Water use效用利水率efficiency 1相Relatively度湿对humidityof air 1 0.899*†数pinkwhiteflower)系关相的间子因响影各与率速合光净的荆紫巨花白2 粉表Table 2Correlations of photosynthetic rate andotherfactorsof C.gigantea(温气Air temperature差压汽水Vapor pressure deficit的度中大CO2 浓气Air CO2 concentration度强照光Photosynthetically activeradiation度温片leaf叶Temperature of率速腾rate蒸Transpiration间Intercellular度CO2 浓CO2胞concentration气Stomatal度导孔conductivity率速合Net 1光rate净photosynthesis率关数速相参Correlation parameters合光rate净Net photosynthesis 1 0.914*度导孔气Stomatalconductivity 1 0.156-0.228胞Intercellular CO2度CO2 浓间concentration 1-0.789 0.130 0.403率速腾蒸Transpiration rate 1 0.855*-0.824*-0.337-0.071度温片叶Temperature of leaf 1 0.703 0.778-0.320 0.157 0.184度强照光Photosynthetically activeradiation 1-0.348-0.855*-0.620 0.811 0.493 0.202 CO2中Air CO2的气度大浓concentration 1-0.850*0.602 0.980**0.795-0.818*-0.477-0.191差压汽水Vapor pressure deficit 1 0.992**-0.846*0.664 0.996**0.828*-0.826*-0.395-0.122温气Air temperature-0.971**-0.991**0.886*-0.501-0.953**-0.740 0.822*0.561 0.260度湿对相Relativelyhumidity ofair-0.861*-0.875*0.903*-0.471-0.856*-0.520 0.586 0.693 0.509率效用利分水Water useefficiency 0.05)。(P <关相著显示表“*”0.01),(P <关相著显极示表**”† ““**”Correlation is significantat the0.01 level,“*”Correlationis significantat the0.05 level.
由表1可知,红花巨紫荆的净光合速率与蒸腾速率、气孔导度之间均呈正相关,其相关系数分别为0.989 和0.706,而与胞间CO2浓度呈负相关,其相关系数为-0.339;净光合速率与叶片温度、光照强度、大气中的CO2浓度、水汽压差、气温、相对湿度、水分利用率等环境因子之间均呈正相关,其相关系数分别为 0.274、0.881、0.083、0.034、0.229、0.007、0.220。显著性检验结果表明,光合速率与气孔导度呈极显著正相关,与光照强度呈显著正相关,而与其它参数间的相关性均不显著。
由表2可知,粉白花巨紫荆的净光合速率与蒸腾速率、气孔导度之间均呈正相关,其相关系数分别为0.914 和0.403,而与胞间CO2浓度呈负相关,其相关系数为-0.228;净光合速率与环境因子中的光照强度、大气中的CO2浓度、相对湿度、水分利用效率之间均呈正相关,其相关系数分别为0.184、0.202、0.260、0.509,而与叶片温度、水汽压差、气温之间均呈负相关,其相关系数分别为-0.071、-0.191、-0.122。显著性检验结果表明,光合速率与气孔导度呈显著正相关,而与其它参数间的相关性均不显著。
4 结论与讨论
植物光合作用的日变化曲线较为常见的有双峰型曲线和单峰型曲线[13-15]。但是,不同的植物其光合“午休”现象的产生原因各异,主要影响因素有气孔限制因素和非气孔限制因素[12,15-17]。相关研究者认为,当净光合速率下降时,胞间CO2浓度与气孔导度同时下降,则表明净光合速率的下降是由气孔限制因素造成的;当净光合速率下降时,胞间CO2浓度或气孔导度却上升,则说明净光合速率的下降是由非气孔限制因素造成的[18-19]。研究中发现,红花和粉白花野生巨紫荆的净光合速率日変化曲线均为双峰型曲线,其峰值分别出现在10:00 和14:00 时左右,光合“午休”现象出现在12:00 时左右,这与许大全[20]有关光合作用的研究结果一致,只是出现峰值的时间先后不同。巨紫荆的净光合速率随着外界光强的升高而升高,在10:00 时就达到了全天中的最大值;10:00—12:00 时,随着外界光强和气温的持续增高,气孔导度下降,胞间CO2浓度降低;12:00 时左右,红花和粉白花野生巨紫荆的净光合速率均最小,其胞间CO2浓度与气孔导度均呈下降趋势,与其光合速率的变化趋势均相同。因此,可以初步判定,气孔限制因素是红花和粉红花野生巨紫荆“午休”现象产生的主要影响因素。
根据分析结果还可以看出,红花巨紫荆的日平均光合速率(4.47 μmol·m-2s-1)及蒸腾速率(4.44 μmol·m-2s-1)及气孔导度的第1 个峰值(0.22 μmol·m-2s-1)均明显高于粉白花巨紫荆的(日平均光合速率为2.59 μmol·m-2s-1,蒸腾速率为2.65 mmol·m-2s-1,气孔导度的第1 个峰值为0.13 μmol·m-2s-1),这一结果表明,红花巨紫荆能够将过多的自然光能转化为热能,减少光抑制现象的发生,使光合速率维持在较高水平上,同时对环境中CO2的固定能力较强。该研究结果与白文玉等[21]对不同种源桤木嫁接幼苗光合特性的研究结果在一定程度上相似,其原因可能是,红花及粉白花巨紫荆叶片生理指标有差异。水分利用效率是衡量植物体单位水量通过蒸腾散失时光合作用所形成的有机物含量的指标之一[22]。根据植物的水分利用效率可以初步判断植物的节水、抗旱及适生能力,一般情况下,植物的水分利用效率越大,则表明植物的节水能力越强,其耐旱能力和适应能力均越强[23-24]。研究中发现,随着光照强度的增加,红花巨紫荆的水分利用效率一天之中的大部分时间均处于较高的水平上,且其水分利用效率最低(1.14 μmol·mmol-1),比粉白花巨紫荆的最低值(0.73 μmol·mmol-1)高出56.16%。这一结果表明,在同样的生存条件下,红花巨紫荆具有较高的合成有机物的能力,且与粉白花巨紫荆相比,其适生与抗旱能力均更强。
植物光合作用是一个复杂的生理过程,其变化不仅与植物内在生理因子有关,还受外在环境因子(如光照、温度、湿度、大气 CO2浓度等)的综合影响[24]。研究中发现,红花巨紫荆的净光合速率与蒸腾速率、气孔导度、叶片温度、光合速率、环境中的CO2浓度、水气压差、气温、相对湿度、水分利用效率之间均呈正相关;粉白花巨紫荆的净光合速率与蒸腾速率、气孔导度、光照强度、环境中的CO2浓度、相对湿度、水分利用率均呈正相关,且红花与粉白花巨紫荆的净光合速率与其气孔导度间的相关性均达到显著水平,表明两种花色野生巨紫荆的光合速率受环境影响的差异不大;红花和粉白花野生巨紫荆的蒸腾速率与其水分利用效率间均呈负相关,且其相关系数均较高,这体现了巨紫荆的净光合速率、蒸腾速率和气孔导度具有较好的协同效应,表明蒸腾速率和气孔导度分别作为影响水分利用率的因子与其均有极其密切的联系,这与魏晓芸等[25]、郑淑霞等[26]、陈香波等[27]的研究结果均一致。
本研究仅对红花和粉白花野生巨紫荆的净光合速率日变化规律及其影响因子进行了观测和分析,虽然能够初步得出两者的光合作用规律及光合特性的差异,但仍有一定的局限性,下一步需从巨紫荆的光合作用与植株生理特性及抗旱适生性等方面的相互关联作用开展深入研究,以探明巨紫荆光合生理的生物学特性,从而为野生巨紫荆的引种驯化、栽植环境的选择及苗木的高效培育和管理提供更为科学的理论依据。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
流行色(2020年2期)2020-04-28
中国农业科技导报(2020年12期)2020-03-15
海峡姐妹(2019年9期)2019-10-08
求学·素材版(2017年12期)2017-12-26
