
天津近岸海域底拖网无脊椎动物群落结构特征及其与环境因子的关系
2021-07-08 11:10张达娟张树林姚冬梅
海洋渔业 2021年3期
张 迎,张达娟,张树林,姚冬梅
(天津农学院水产学院,天津市水产生态及养殖重点实验室,天津 300384)
天津近岸海域位于38°20′~39°30′N、117°17′~118°20′E,地处华北平原东北部、渤海湾西端,其范围东起陡(涧)河口,南至岐口,从最高潮线向陆域推进10 km,至海域-20 m等深线。该海域渔业资源十分丰富,是我国鱼、虾、蟹、贝的重要养殖区,也是我国北方渔业的“产房”[1]。
无脊椎动物占了海洋动物的绝大部分,是海洋动物中门类最为繁多的一类,部分种类具有非常重要的生态价值与经济价值[2]。其中,贝类、海蟹、对虾是人类重要的动物性蛋白质来源。同时,无脊椎动物作为海洋生态系统的重要组成部分,在海洋生态系统物质循环和能量流动中具有不可替代的作用。此外,因其具有对逆境状况逃避能力较差、易受环境影响的特点[3],可用于反映其所处环境长期和宏观的变化状况。
20世纪70年代至今,国内对于海洋无脊椎动物群落及多样性的研究主要包括渤、黄海的渔业资源群落及栖息环境研究[4-8],以及其他各海域和河口无脊椎动物群落及多样性的研究[9-22]。国外学者也分别从种类组成、数量变动、群落时空格局和群落结构影响因子等方面对海洋无脊椎动物群落进行了研究[23-30]。而天津近岸海域无脊椎动物的系统性研究可上溯至20世纪80年代,张闰生和孙秋岩[1]系统调查了天津近岸海域无脊椎动物的种类组成和时空分布。此后,该区域无脊椎动物相关研究多集中于对其生物学特性的研究[31-32],而对于天津近岸海域无脊椎动物的群落结构特征与环境因子关系的研究尚少见报道。通过广义加性模型(generalized additive model,GAM)可以直观地评估各环境因子对生物的影响程度及重要性,揭示渔业资源与多环境要素之间的关系。本文通过2019年4月和6—11月天津近岸海域无脊椎动物及环境因子的调查数据,结合GAM模型方法,分析了该区域内无脊椎动物群落结构特征及其与环境因子间的关系,以期为天津近岸海域渔业资源的可持续开发利用提供科学数据和参考资料。
1 材料与方法
1.1 站点设置
2019年4 月和6—11月在天津近海开展了每月1航次的调查,共计7航次,调查区域为117°40′~118°00′E、38°30′~38°50′N,共设9个调查站位(图1)。
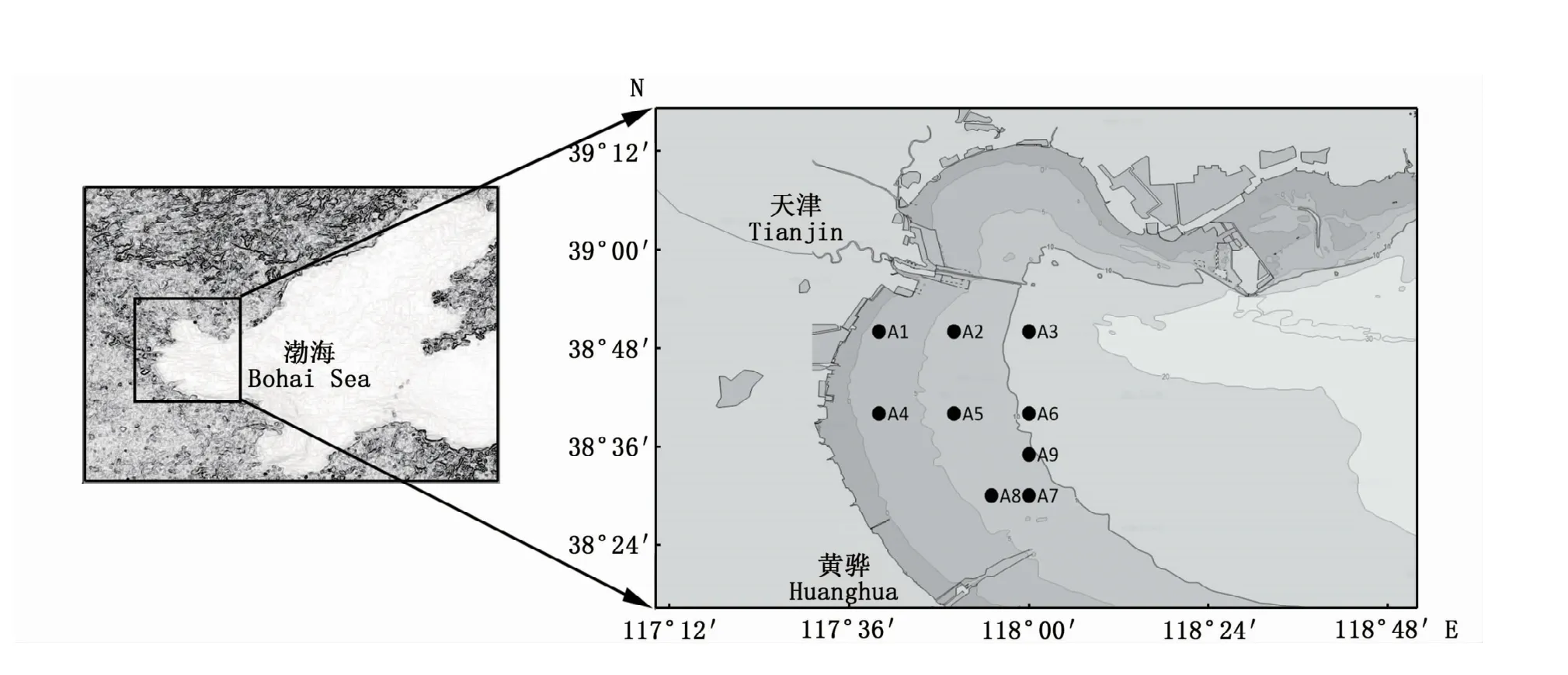
图1 2019年天津近海调查站位Fig.1 Tianjin offshore survey stations in 2019
1.2 采样方法
调查采用110.3 kW渔船,拖网速度为2 kn,每个调查站位拖网时间均为1 h,调查网具为底拖网,网口宽度8 m,网囊网目20 mm。将渔获物中的无脊椎动物鉴定种类,并记录各种类生物量及个体数,数据均进行标准化处理,换算成每小时的生物量(kg·h-1)和丰度(个·h-1)。同时现场测定pH、氧化还原电位、水温、溶解氧、盐度等环境因子。
1.3 数据分析
1.3.1 多样性分析
Shannon-Wiener多样性指数(H′)计算公式:
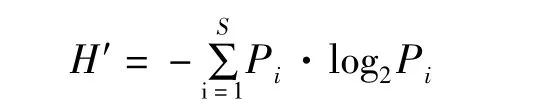
式中,H′为多样性指数;S为物种总数;Pi为样品中第i种的丰度(Ni)与各站位总丰度(N)比值。
Margalef丰富度指数(D)计算公式:
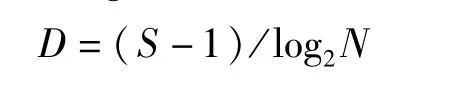
式中,D为丰富度指数;S为物种总数;N为各站位总丰度。
Pielou均匀度指数(J)计算公式:
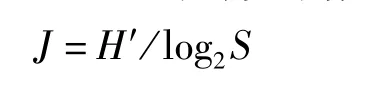
式中,J为均匀度指数;H′为多样性指数;S为物种总数。
1.3.2 优势种
利用Pinkas相对重要性指数确定每一种类在群落中的重要性。相对重要性指数的计算公式为:
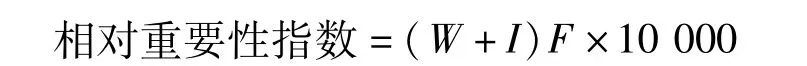
式中,W为某一种类的生物量占总生物量的百分比;I为某一种类的个数占总个数的百分比;F为某一种类出现站位数占调查总站位数的百分比。将相对重要性指数≥1 000的种类定义为优势种,100≤相对重要性指数<1 000定义为重要种,10≤相对重要性指数<100定义为常见种,1≤相对重要性指数<10定义为少见种,相对重要性指数小于1定义为偶见种[6]。
1.3.3 群落结构分析
采用PRIMER 6.0进行多元统计分析:首先对物种的丰度数据进行四次方根转换,以平衡优势种和稀有种在群落中的影响,并计算站位间的Bray-Curtis相似性系数,构建相似性矩阵;然后采用cluster等级聚类方法和非度量多维尺度(nonmetric multi-dimensional scaling,NMDS)排序方法分析无脊椎动物的群落分布格局。通过ANOSIM(analysis of similarities)分析不同聚类组间的差异性,并通过SIMPER(similarity percentage)分析不同物种对各群落组内相似性和组间差异性的平均贡献率[33-34]。
1.3.4 群落与环境因子相关性分析
应用广义加性模型(generalized additive model,GAM)分析无脊椎动物与环境影响因子之间的关系,模型表示为:
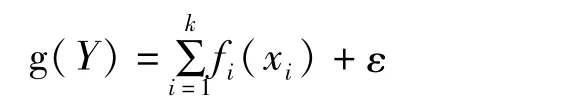
式中,Y为无脊椎动物丰度;g(Y)为连接函数;xi为各环境因子;fi(xi)为g(Y)与第i个环境因子的非参数函数;k为环境因子个数;ε为误差项。
本研究中,连接函数为自然对数,模型采用样条平滑法,分布函数族为泊松分布,模型的误差分布估计为高斯分布,分析通过R 3.6.3软件及mgcv包实现。
2 结果与分析
2.1 天津近岸海域无脊椎动物群落结构特征
2.1.1 无脊椎动物物种组成
2019年天津近岸调查海域共发现无脊椎动物34种,其中节肢动物和软体动物均为16种,各占总物种数的47.06%,其余分别为螠虫动物1种(2.94%)和棘皮动物1种(2.94%)(表1)。

表1 2019年天津近海无脊椎动物物种组成Tab.1 Species composition of invertebrates off the coast of Tianjin in 2019
以月份和站位为因素对物种数进行双因素方差分析,结果显示,2019年物种数在月份间有显著性差异(F=5.245,P=0.000),在站位间无显著性差异(F=0.676,P=0.710),表明2019年物种数仅存在时间变化特征。
2.1.2 无脊椎动物群落优势种
2019年优势种为凸壳肌蛤(Arcuatula senhousia)、火 枪 乌贼(Loligobeka)、日本蟳(Charybdisjaponica)、口虾蛄(Oratosquilla oratoria);重要种为三疣梭子蟹(Portunus trituberculatus)、天津厚蟹(Helicetientsinensis)、中国明对虾(Fenneropenaeuschinensis)。各优势种类别及相对重要性指数见表2。
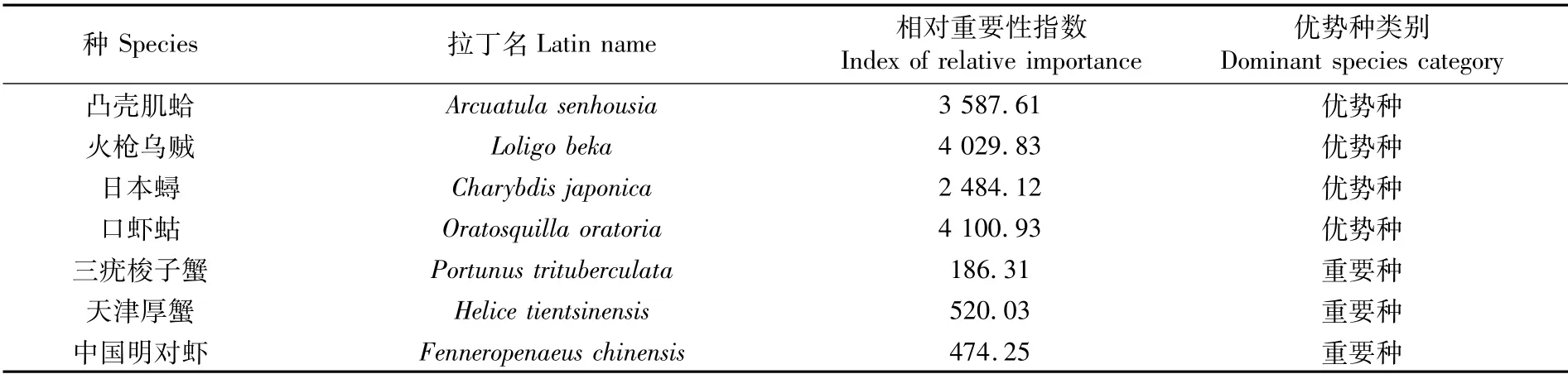
表2 2019年无脊椎动物优势种及相对重要性指数Tab.2 Dominant species and relative importance index of invertebrates in 2019
2.1.3 无脊椎动物丰度和生物量
由图2可知,2019年无脊椎动物平均丰度为1 267.9个·h-1,平均生物量为9.58 kg·h-1。以月份与站位为因素对2019年无脊椎动物丰度、生物量进行双因素方差分析,结果显示,2019年丰度在月份间有显著性差异(F=2.400,P=0.042),在站位间无显著性差异(F=0.748,P=0.649);2019年生物量在月份间有显著性差异(F=10.357,P=0.000),在站位间无显著性差异(F=1.868,P=0.087),表明无脊椎动物群落丰度、生物量仅存在时间变化特征。
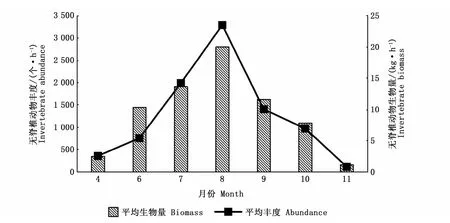
图2 2019年天津近海无脊椎动物丰度、生物量时间变化Fig.2 Temporal variations of invertebrate abundance and biomass off the coast of Tianjin in 2019
2.1.4 无脊椎动物多样性
从表3可见,丰富度指数D、均匀度指数J、多样性指数H′的变动规律基本相似。其中,多样性指数H′A1站位最高,A3站位其次,A2站位最低;从整体上看,2019年多样性指数H′平均值为2.81(2<2.81<3),表明物种丰富度高,个体分布均匀。
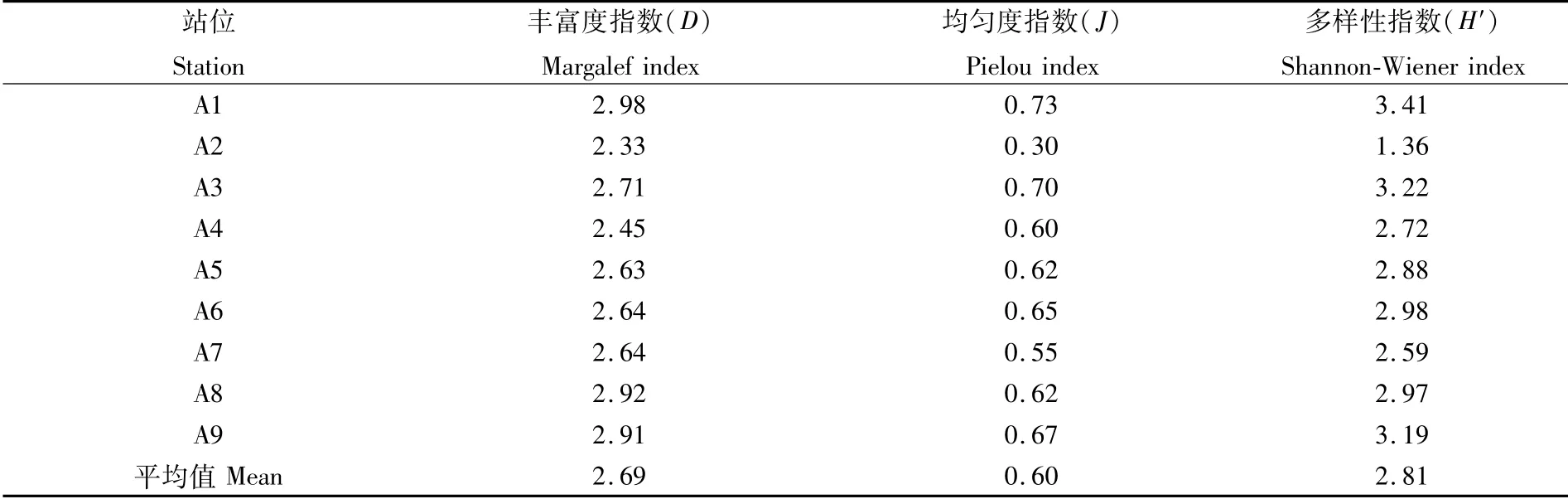
表3 2019年天津近海无脊椎动物物种多样性Tab.3 Species diversity of invertebrates off the coast of Tianjin in 2019
2.2 无脊椎动物群落结构分析
无脊椎动物群落结构聚类分析结果见图3,且NMDS分析与cluster聚类分析结果一致。ANOSIM分析表明,2019年调查的不同聚类组间均存在显著性差异(R=0.827,P<0.05,global test)。
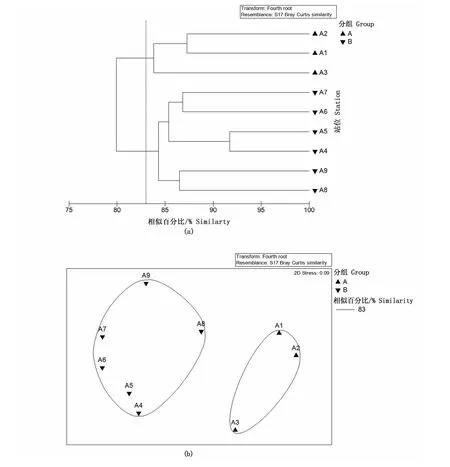
图3 2019年天津近海无脊椎动物站位聚类和非度量多维尺度分析Fig.3 Site clustering and multi-dimensional scale sequencing analysis of invertebrates off the coast of Tianjin in 2019
稀有种去除后,以83%的相似性将2019年调查区域的无脊椎动物群落分为2组,A1、A2和A3为A组,区域分布集中在北部,分组等级较高(pairwise test);A4、A5、A6、A7、A8和A9为B组,区域分布集中在南部,分组等级较高(pairwise test)。
A组:由3个站位组成,组内平均相似性为84.96%,其中包括15种贡献物种,各物种贡献率均低于10%。
B组:由6个站位组成,组内平均相似性为85.40%,其中包括16种贡献物种,主要贡献物种火枪乌贼贡献率为10.26%,其余各物种贡献率均低于10%。
2.3 环境因子相关性分析
基于2019年无脊椎动物丰度及环境调查结果,运用GAM分析无脊椎动物丰度与温度、氧化还原电位、溶氧、pH、盐度等环境因子的关系(表4)。结果表明,除pH和盐度外(P>0.05),氧化还原电位、溶解氧、温度对2019年无脊椎动物丰度均有显著影响(P<0.05),解释偏差依次为7.06%、51.60%、63.60%。

表4 2019年无脊椎动物丰度与相关环境因子GAM的解释偏差及显著性Tab.4 Explanatory bias and significance of invertebrate abundance and related environmental factors GAM in 2019
基于无脊椎动物丰度与环境因子的GAM预测结果可以看出,除pH和盐度外,氧化还原电位、溶解氧、温度对2019年无脊椎动物丰度预测的置信区间小,预测的可靠度高。随着氧化还原电位、溶解氧、温度的升高,无脊椎动物丰度整体上呈现上升的趋势(图4)。
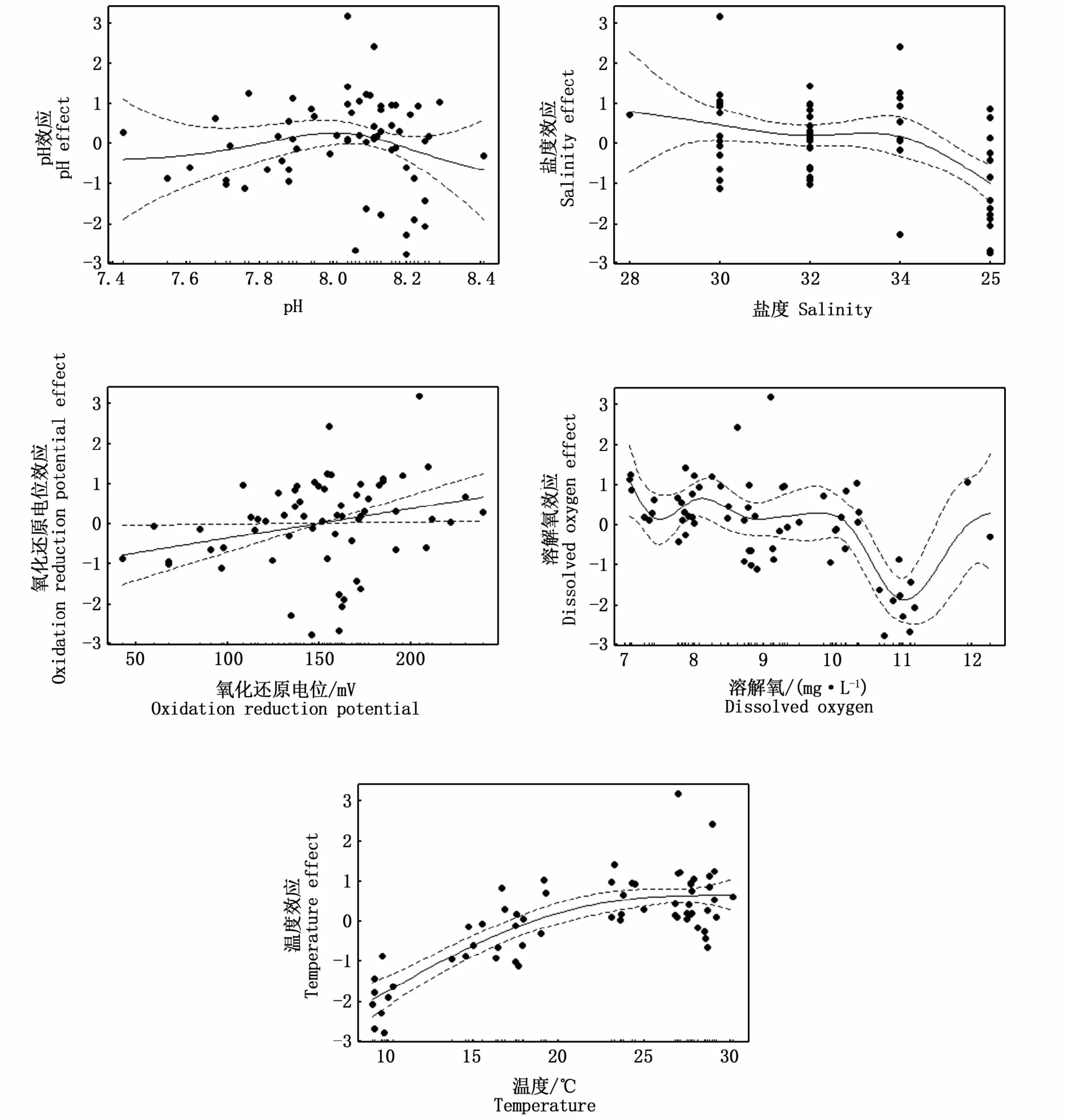
图4 2019年无脊椎动物丰度与环境因子GAM分析Fig.4 GAM analysis of invertebrate abundance and envirnmental factors in 2019
3 讨论
3.1 天津近岸无脊椎动物种类、优势种及群落结构变化
无脊椎动物作为一类活动能力相对较弱的海洋生物类群,在受到人类活动的影响后,其生物种类及群落组成都会发生变化[35]。结合20世纪80年代天津市海岸带和海涂资源综合调查[1]数据可知,20世纪80年代该区域有无脊椎动物40种,而本次调查(2019年)共捕获到34种。从较长历史跨度分析,本次调查较20世纪80年代减少的种类为泥脚隆背蟹(Carcinplaxvestitus)、脊腹褐虾(Crangonajjinis)、鹰爪虾(Trachypenaeus curvirostris)、周氏新对虾(Metapenaeusjoyneri)、曼氏无针乌贼(Sepiellamaindroni)和日本枪乌贼(Loligojaponica);增加的种类有白带三角口螺(Trigonostomascalariformis)、凸壳肌蛤、日本拟平家蟹(Heikeopsisjaponica)和颗粒拟关公蟹(Paradorippegranulata)。推测出现这种种类明显交替现象的原因,一方面可能是由于20世纪80年代调查采样的15个站位与本研究选取的调查站位有所差异,另一方面可能是由于随着时间推移,区域内群落结构发生了较大变化,且部分调查站位受人为影响严重,目前已禁止捕捞。张闰生和孙秋岩[1]的研究也提及受调查时间、站点位置、数目和环境等影响,调查统计结果会有所差异。
优势种由于其高度的生态适应性,可用于分析判断群落结构的变化[36]。20世纪80年代无脊椎动物优势种为中国明对虾、鹰爪虾、曼氏无针乌贼、火枪乌贼、日本枪乌贼和口虾蛄,而本次调查除火枪乌贼、口虾蛄仍为优势种外,优势种类型发生了很大的变化,新增了三疣梭子蟹、日本蟳、凸壳肌蛤等优势种。经济价值较高的无脊椎动物种类的转变,究其原因,可能是由于经济利益的驱使,盲目发展捕捞船只和网具所导致。随着捕捞压力的急剧增加,无脊椎动物资源受到了严重的影响,种群数量和质量降低,资源补充群体数量也急剧减少,从而使优势种结构发生了改变。
通过研究2019年天津近岸海域无脊椎动物群落资源量变化情况发现,无脊椎动物群落丰度、生物量在各站位间的空间分布无显著差异,但存在随时间变化的特征。这可能是由于春、秋两季,季节洄游性种类如甲壳类动物游出湾内,以及常栖性种类进行深浅水的短距离洄游,导致丰度、生物量处于较低的状态。而占据优势的口虾蛄、火枪乌贼于春季产卵繁殖,在夏季出现大量幼体群,同时在补充群体的主导作用下,丰度、生物量在夏季达到最大值,导致了其在时间上的明显差异。由于海上取样和站位设置的差异,进行可靠的比较异常困难,但通过与历史资料对比还是可以看出一些变化。因此仅就与渤海湾相同采样方式的历史资料对比而言,本次调查结果显示,天津近海无脊椎动物的资源生物量四十多年间已发生了不同程度的变化。1982—1983年[1]和1992—1993年[37]的 资料 显 示,1982—1983年无脊椎动物资源生物量为21.38 kg·h-1;1992—1993年和10年前相比,无脊椎动物资源量下降了56.82%,资源生物量呈下降趋势。根据本文研究结果,本次调查的无脊椎动物资源生物量明显低于历史调查数据,表明天津近海无脊椎动物目前处于一个较低的水平,资源有明显衰退的迹象。
结合cluster及ANOSIM分析可知,按照无脊椎动物丰度水平,可将2019年无脊椎动物群落划分为2个区域组群,虽然丰度在站位间差异不明显,但区域组间贡献物种的贡献率存在明显的差异,例如A组站位凸壳肌蛤的偶然性大量采集,导致其整体分布趋向于北部;而火枪乌贼作为影响分组的主要贡献物种,因受季节及环境因子的影响较大,导致其分布趋向于南部,进而影响到了整体空间格局的分布。相关研究也表明,由于不同区域一些环境因子的差异,无脊椎动物群落结构呈不同的表现[12],此外,站位设置的密集程度也可能是影响群落空间格局的重要因素,因此还需进一步扩大研究区域并分析总结。
3.2 无脊椎动物丰度变化及其与环境因子的关系
海洋环境是海洋生物的生存基础,对海洋生物生长的各个阶段都具有重要影响[38]。水温是影响无脊椎动物群落最重要且稳定的环境因子[9],溶解氧是海洋无脊椎动物呼吸生存的必要条件,并且对海洋生物栖息环境中众多物质的氧化分解起主导作用;pH对调节海洋无脊椎动物体内的酸碱平衡、气体交换、血氧运输、渗透压等极为重要[39]。此外,还包括水深、盐度等[40],有研究表明,盐度与水温显著影响东海和长江口及邻近海域无脊椎动物的群落结构[41]。但海洋环境经过长时间的变化,各环境因子对天津近岸海域无脊椎动物的影响以及影响范围、程度仍有待系统性的研究。
本次调查选取了pH、氧化还原电位、溶解氧、温度和盐度等环境因子,并结合无脊椎动物丰度进行GAM分析。结果表明,温度、氧化还原电位和溶解氧是影响无脊椎动物丰度的主要因素(P<0.05),随着温度、氧化还原电位和溶解氧的升高,无脊椎动物丰度整体呈现上升的趋势,这与金显仕等[39]认为无脊椎动物丰度与溶解氧显著相关的结果相似,与金显仕等[39]认为无脊椎动物丰度与pH显著相关的结果相反;与陈大刚[40]认为无脊椎动物丰度与温度显著相关的结果相似,与陈大刚[40]认为无脊椎动物丰度与盐度显著相关的结果相反。这可能是因为本次调查海域的盐度变化范围较小,且天津近海环境中盐度变化也始终保持较为稳定的态势,因此导致本研究中盐度与无脊椎动物丰度间的相关性不显著;而pH与无脊椎动物丰度无显著相关性的结果仍需进一步研究。且上述研究多是无脊椎动物与环境因子的关联程度,未从丰度的整体变化来综合考虑,因此本研究运用GAM分析无脊椎动物丰度与环境因子的整体变动,更能体现无脊椎动物与环境因子的关系。
对天津近岸海域无脊椎动物丰度和环境因子进行周年监测,将其变化情况可视化,一方面可以为渔业资源的可持续开发利用提供科学资料,另一方面通过了解无脊椎动物群落变化的规律和影响因素,可适时采取措施来应对日益加重的环境胁迫和人为影响导致的无脊椎动物群落结构的剧烈变化。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
昆明医科大学学报(2022年2期)2022-03-29
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
人大建设(2019年5期)2019-10-08
