
三角褐指藻DGAT1基因生物信息学分析及表达调控研究
2021-07-07 11:37:10王何瑜龚一富郑小恽李申睿
宁波大学学报(人文科学版) 2021年4期
王何瑜, 龚一富, 郑小恽, 李申睿
三角褐指藻1基因生物信息学分析及表达调控研究
王何瑜1, 龚一富2,3*, 郑小恽2,3, 李申睿2,3
(1.宁波大学 食品与药学学院, 浙江 宁波 315832; 2.宁波大学 海洋学院, 浙江 宁波 315832; 3.浙江省海洋生物工程重点实验室, 浙江 宁波 315832)
本研究采用转录组测序获得了三角褐指藻1基因cDNA全长序列(GeneBank登录号: 7200924), 并对其进行生物信息学分析和表达调控研究. 结果表明, 三角褐指藻1基因cDNA序列全长为1438bp, 开放阅读框(ORF)为1098bp, 编码365氨基酸序列, 含有脂肪酸蛋白特性(I)和DAG结合位点(II)等功能结构域. 预测三角褐指藻DGAT1蛋白为亲水性蛋白, 含有6个跨膜结构域和7个超强跨膜螺旋区, 无信号肽. 进化树分析表明, 三角褐指藻与海链藻DGAT1蛋白同源性最高. RT-qPCR结果表明, 光照强度和温度均显著促进三角褐指藻1基因的表达. 随着光照强度和温度的增加, 三角褐指藻1基因的表达量呈先升高后降低趋势, 在光照强度为2500lx或温度为25℃时, 三角褐指藻1基因的表达量达到最大, 这与三角褐指藻总脂含量的变化趋势一致, 同样揭示DGAT1蛋白与三角褐指藻总脂生物合成与积累密切相关.
三角褐指藻;1基因; 克隆; 生物信息学; 基因表达调控
二脂酰甘油酰基转移酶(Diglyceride Acyltrans- ferase, DGAT)是三脂酰甘油(TAG)合成途径中的唯一限速酶, 催化二脂酰甘油结合脂肪酸酰基形成TAG. 三角褐指藻()是海洋微藻中生长较快、脂质含量较高的藻种[1], 其油脂含量约占细胞干重的20%~30%, 是一种可以用来制备生物柴油的微藻[2]. 三角褐指藻碳储存的主要形式为油脂, 特别是三脂酰甘油(TAG). 甘油-3-磷酸酰基转移酶(Glycerol-3-Phosphate Acyltrans- ferase, GPAT)催化3-磷酸甘油形成溶血磷脂酸, 随后由1-酰基甘油-3-磷酸酰基转移酶(AGPAT)催化生成磷脂酸, 随后在磷脂酸磷酸酶(Phospha-tidic Acid Phophyhydrolase, PAP)的催化下形成甘油二脂, 最后在二脂酰甘油酰基转移酶(Diglyceride Acyltransferase, DGAT)的催化下生成TAG. DGAT是TAG合成途径中的最后一步, 也是TAG合成途径中的限速酶[3], 能调控脂肪代谢以及脂类在细胞中的沉积. 根据其结构差异, DGAT有DGAT1、DGAT2、WS/DGAT和胞质内DGAT (CytoDGAT)共4种类型[4]. DGAT1与DGAT2是普遍存在的2个亚型, DGAT1主要作用于TAG代谢, 而DGAT2则侧重于积累特殊脂肪酸[5]. DGAT通过影响种子发育过程中TAG含量, 对种子质量和脂肪酸组成成分起一定作用[6]. Jako等[6]发现将基因插入拟南芥染色体中, 能降低拟南芥种子中DGAT1活性, 从而减少TAG含量与油脂含量. Lardizabal等[7]将深黄伞形霉()2在大豆中超量表达后, 使其种子的含油量提高了1.5%. 研究表明, 在有利条件下微藻会积累大量脂质[8], 三角褐指藻可以作为生产生物燃料的有效原料广泛应用[9]. 目前1基因已在玉米()[10]、大豆()[11]、棉花()[12]与油茶()[13]等陆地植物中克隆出来, 但未见从海洋藻类, 特别是三角褐指藻中有克隆1基因的报道. 鉴于三角褐指藻是目前油脂生产的理想海洋藻类, 从三角褐指藻中克隆1基因, 深入研究藻类油脂积累分子机制, 并采用代谢工程策略提高藻类油脂含量, 以期提供优良基因资源, 为最终实现藻类的生物柴油生产提供前期的研究基础.
不同微藻藻种积累脂肪的能力不同, 但当它们处于不利生长时, 微藻往往会合成大量的脂肪. 影响微藻脂类合成的因素有光照、温度、pH、营养盐等. 光照可促进陆地植物油脂的积累, 光反应为质体中脂肪酸的合成提供ATP和NADPH, 激活Rubisco旁路, 使得在油脂合成过程中能更有效地利用碳流[4]. 吕文兵等[14]发现, 杜氏盐藻()的叶绿体基因的表达量在光照强度4500lx时显著升高, 但是过高的光强反而会抑制基因的表达. 温度通常会影响油料种子的脂肪酸组成, 其原因是温度影响了2基因和PEP羧化酶活性, 进而影响到油脂的合成[4]. 光照和温度可调控陆地植物脂类合成, 在微藻研究中也有类似报道[15-16]. 但光照强度和温度对微藻1基因的表达调控研究较少, 光照和温度是否通过调节1基因进而表达来促进三角褐指藻脂类的含量, 目前还未见相关报道. 因此, 本研究通过转录组测序获得三角褐指藻1基因cDNA全长序列, 通过研究三角褐指藻在不同光照强度和温度下,1基因转录差异及其与脂质含量之间的相关性, 以期为进一步研究三角褐指藻脂质合成与积累的途径、机制提供理论依据.
1 材料与方法
1.1 三角褐指藻培养与处理
三角褐指藻藻种由宁波大学海洋生物工程实验室微藻种质库提供, 培养方法参照文献[17]方法进行. 取高温灭菌的过滤海水, 以1:1000比例加入MVA母液[18], 以1:3(藻种:海水)比例接藻. 培养温度为(20±0.5)℃, 光暗周期比12h:12h, 每天摇瓶3次. 光照处理组的培养温度为(20±0.5)℃, 光照强度为0、1000、2500、5000lx, 每个处理组设置3个平行. 温度处理组的光照强度为1000lx, 温度设置为15、20、25、30℃, 每个处理组设置3个平行. 培养至第7天时, 测定总脂含量.
1.2 三角褐指藻DGAT1基因的获得与生物信息学分析
取处于对数生长期的三角褐指藻藻液, 用植物总RNA提取试剂盒(北京奥莱博生物技术有限公司)提取总RNA. 用反转录试剂盒(TaKaRa, 大连)将总RNA反转录成cDNA. 通过转录组测序分析获得三角褐指藻1基因cDNA全长序列, 设计扩增ORF的引物(F-DGAT: 5’-ATGTTGTGCC GGAAATACTTC-3’, R-DGAT: 5’-TCAACGAATC AAGCA GGA AT-3’)进行PCR验证, PCR产物经回收后连接T载体, 转化大肠杆菌DH5a, PCR验证后送公司测序.
使用Vector软件对三角褐指藻和其他物种的DGAT1蛋白进行多序列的比对(表1)[19]. 通过ProtParam (http://eapasy.org/tools/protparam.html)在线工具对三角褐指藻DGAT1氨基酸序列进行分析, 预测其相对分子量、等电点等特性, 并用ExPASy预测DGAT1蛋白质的二级结构[20]. 利用TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/)对三角褐指藻DGAT1蛋白进行跨膜区域预测. 用Clustal_X 1.8和Mega 3.0软件对DGAT1蛋白进行系统进化树构建.

表1 DGAT1氨基酸序列分析所用物种
1.3 三角褐指藻DGAT1基因的表达调控研究
对三角褐指藻光照处理组和温度处理组培养24h, 然后分别提取总RNA, 并反转录. 以三角褐指藻基因作为管家基因, 引物为-F (5’-CGTGACTTGACGGACTACCTG-3’)和- R(5’-TAGTTTTTGTCCAGAGCCGAG-3’). 三角褐指藻1基因的定量PCR引物为1-F(5’- GGTATGTTTGCGTAGACTTGT-3’)和1-R(5’- TCGTTATTTACTTCGGAGATT-3’), 由生工生物工程(上海)有限公司合成. 使用SYBR Premix Ex TaqTM(TaKaRa, 大连)进行荧光定量PCR. 荧光定量PCR反应体系为SYBR®Premix Ex TaqTM(2×) 10μL, F-prime、R-primer(10μmol∙L-1)各0.8μL, cDNA模板2μL, 超纯水6.4μL. 每组样品重复3次, 反应程序: 94℃3min; 94℃ 30s, 53℃ 40s, 72℃ 20s, 40个循环; 72℃延伸10min, 4℃保存. 三角褐指藻1基因相对转录水平用2-ΔΔCt方法计算, 使用SPSS 20.0软件对数据进行分析[21].
1.4 三角褐指藻总脂含量的测定
分别取光照处理组和温度处理组培养7d的三角褐指藻30mL, 4℃, 5000r∙min-1离心10min. 使用无菌水冲洗2~3次, 并在4℃, 8000r∙min-1条件下离心10min. 总脂含量的测定采用改良的Bligh- Dyer法[22].

加粗下划线标注为起始密码子; 加粗下划线下带有“*”标注为终止密码子; 下划线标注部分为保守序列.
2 结果与分析
2.1 三角褐指藻DGAT1基因的生物信息学分析
通过三角褐指藻转录组测序分析获得了1基因的cDNA全长序列, 并经过PCR测序验证. 三角褐指藻1基因cDNA序列全长为1438bp, 开放阅读框(ORF)为1098bp, 编码365氨基酸序列(图1).
蛋白质多序列比对结果表明(图2), 获得的DGAT1蛋白序列属于跨膜O-酰基转移酶超家族(Membrane-Bound O-Acyltransferases Superfamily, MBOAT). 三角褐指藻DGAT1氨基酸序列含有脂肪酸蛋白特性(I)和DAG结合位点(II)结构域, DGAT1具有ACC合成酶(ACC synthase)家族基因保守基团FYXDWWN.
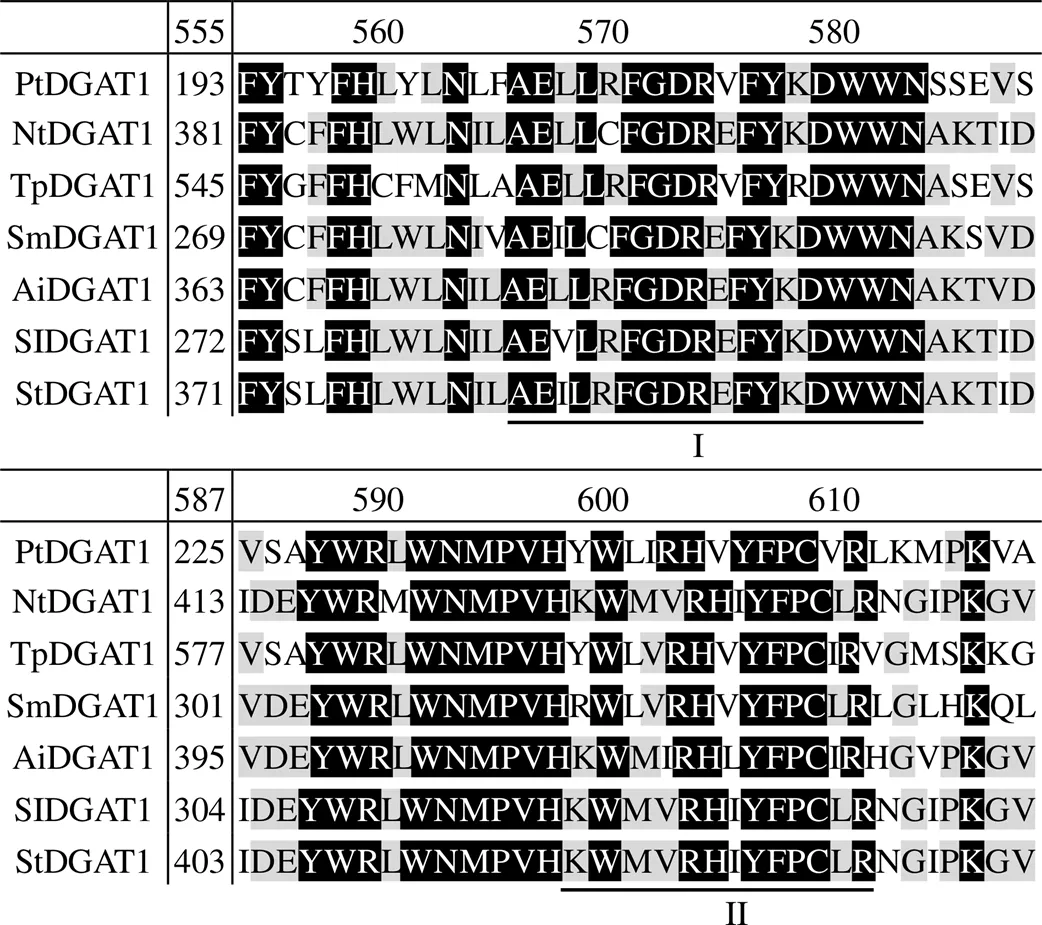
I为脂肪酸蛋白特征; II为DAG结合位点.
2.2 三角褐指藻DGAT1蛋白质基本理化分析
三角褐指藻DGAT1蛋白理化性质分析结果表明, 三角褐指藻DGAT1蛋白由365个氨基酸组成, 总原子数为6054, 分子式C2018H3020N494O500S22, 相对分子质量为42906.51, 等电点为8.77. DGAT1蛋白中亮氨酸(Leu)含量最高, 占总氨基酸的12.9%, 半胱氨酸(Cys)含量最少, 不含有吡咯赖氨酸(Pyl)和硒半胱氨酸(Sec). 三角褐指藻DGAT1蛋白的正电荷残基数(Asp+Glu)为25, 负电荷残基数(Arg+ Lys)为30, 不稳定指数为37.97, 推测该蛋白为稳定蛋白, 其脂肪指数为103.92.
三角褐指藻DGAT1蛋白的N端为亲水区, C端为疏水区, 位于第264位的苯丙氨酸(Phe)有最强亲水性, 最大值为2.867, 在约25~50、80~90、100~115、140~155、180~205、250~280和320~330位置AA处有7个亲水峰. 其他区域则均以疏水性区域为主, 339位的Gly具有最强疏水性, 值为-2.34, 总平均亲水系数为0.333, 推测DGAT1蛋白为亲水性蛋白.
通过TMHMM-2.0软件能够推测三角褐指藻DGAT1蛋白存在6个跨膜结构域, 其具体位置在13~35、39~61、134~153、186~208、259~281和314~336氨基酸处. 通过TMPRED推测, 三角褐指藻DGAT1蛋白可能含有7个强跨膜螺旋区域.
2.3 三角褐指藻DGAT1蛋白结构预测
二级结构预测结果表明, 三角褐指藻DGAT1蛋白中α-螺旋所占区域比例最高, 约为41.37%, 延长链约占25.48%, 无规则卷曲也同样约占25.48%, β-转角所占比例最低, 约7.64%.
2.4 三角褐指藻DGAT1蛋白系统进化树分析
相似性搜索及比对出17个物种18种DGAT1蛋白基因(图3), 进化树分为3个分支, 被子植物、藻类和动物各汇成分支, 三角褐指藻与海链藻的亲缘关系最近, 汇成一支. 而小球藻与其他藻类生物同源性不高, 反而与被子植物间的亲缘关系较近, 但也是植物中最原始的, 这与目前藻类在分类上将部分藻类划归植物, 而部分藻类划归为微生物的结果一致. 从进化树结果表明, 三角褐指藻可划入微生物类.
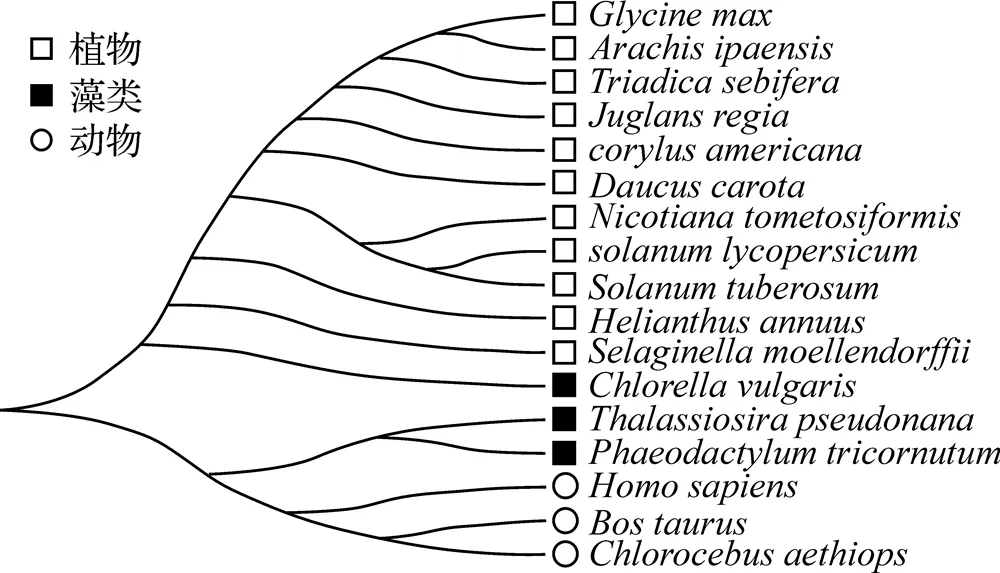
图3 三角褐指藻与其他物种的DGAT1氨基酸序列系统进化树
2.4 光照强度和温度对三角褐指藻总脂含量及DGAT1基因表达的影响
不同光照强度对三角褐指藻1基因的表达量和总脂含量结果表明(图4(a)), 随着光照强度的增强, 三角褐指藻1基因表达量不断上升, 在光强为2500lx时,1基因表达量达到最大, 此后呈下降趋势. 与此同时, 三角褐指藻总脂含量也与1基因表达量呈相同趋势, 总脂含量随光强增大而增加, 超过2500lx后总脂合成减少, 说明光照强度对三角褐指藻脂类合成的影响与1基因表达量相关.
不同温度对三角褐指藻总脂含量和1基因表达量的影响结果表明(图4(b)), 25℃时的三角褐指藻1基因表达量和总脂含量均达到最大值, 30℃与其相比差异并不显著. 而培养温度低于20℃时, 三角褐指藻1基因表达量受到明显抑制, 此时的总脂含量也减少明显, 说明温度对三角褐指藻脂类合成的影响与1基因的表达量相关.
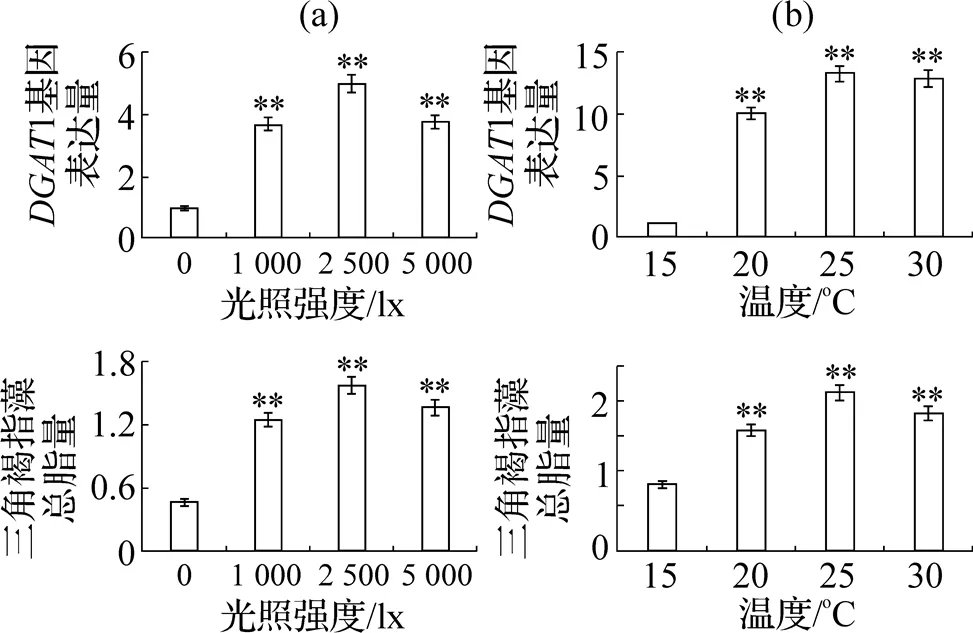
图4 不同处理条件下的三角褐指藻DGAT1基因表达量和总脂含量
研究三角褐指藻总脂的含量与1基因表达量的关系, 可以发现三角褐指藻总脂含量与1基因表达量呈正相关性.1基因表达受抑制时, 总脂含量也随之下降;1基因表达增加时, 总脂含量也相应增多. 两者具有高度的一致性, 表明DGAT1蛋白是三角褐指藻脂类合成和积累途径中的关键酶基因之一.
3 讨论
本文获得的三角褐指藻DGAT1氨基酸N端序列包括脂肪酸蛋白特征和DAG结合位点, 该特征与油菜DGAT1蛋白结构相类似[23]. 通过DGAT蛋白结构特点与序列比对, 认为该基因属于DGAT1. DGAT1蛋白一般含有6~9个跨膜区域[24], 蛋白较大, N端有亲水域位于内质网膜的胞质面[25]. DGAT1具有ATACS保守基团FYXDWWN, 可能与酰基辅酶A的形成有关[26]. 三角褐指藻DGAT1氨基酸序列第215~221也为该基团, 且位于脂肪酸蛋白特征处. 人与动物DAGT1蛋白N端功能研究表明, 此段序列可能与acyl-CoA互相作用形成二聚体或四聚体[27].
从进化树可看出, 三角褐指藻与海链藻的亲缘关系较近, 与其他陆生植物、动物的亲缘关系较远. 郝敬云等[27]则发现莱茵衣藻()2家族中的CreDGTT1-5与三角褐指藻有较近的关系. 而张晓琼等[28]研究模式生物2基因进化树时发现, 藻类植物DGAT蛋白独自形成进化分支, 并且与其他陆生植物的亲缘关系较远. 序列分析可以发现各个物种具有较高的多样性[29], 但是基因家族动植物之间的同源关系及其起源仍然不够清晰, 需要进行更加深入的探究.
在RT-qPCR结果中, 三角褐指藻1基因表达量和总脂含量对于光照和温度的变化呈相类似的趋势. 适当加强光照、升高温度可增加三角褐指藻1基因的表达量和总脂含量. 进一步研究发现1基因与三角褐指藻脂质合成与积累呈正相关, 说明1基因是脂类合成的关键酶基因. 尹航等[15]的研究结果也表明, 在一定温度和光照强度处理下, 三角褐指藻基因的表达量与总脂含量呈显著正相关, 说明光照强度和温度可通过促进三角褐指藻内TAG生物合成途径中的1和关键酶基因的表达, 增加关键酶活性, 进而加速藻内脂类合成与代谢. 而在高温和强光胁迫下, 不仅酶活性受到抑制, 藻细胞的生理活性也会遭到破坏, 导致细胞死亡. 沈城等[30]发现持续高温胁迫使珊瑚共生虫黄藻()基因的表达量随着温度的升高显著增加. 华雪铭等[16]实验发现, 35℃下的绿色巴夫藻()全部死亡, 10℃时的藻细胞大量死亡, 而在最适温度20℃下, 绿色巴夫藻生长最快, 合成脂质最多. 周洪琪等[31]研究发现, 不同藻类的最适温度和光照不相同, 但总是在最适时有最大生长量和总脂含量. 以上结果表明, 适宜的光照和温度能促进脂类生物合成, 除了与脂类生物合成途径基因1和的表达有关以外, 还有可能通过基因的表达与光合作用有关. 如果为了更深入地研究光照和温度促进微藻脂类合成的分子机理, 还需要采用转录组学技术、代谢组学技术等技术进行深入的研究.
4 结论
三角褐指藻1基因cDNA序列全长为1438bp, ORF为1098bp, 编码365氨基酸序列, 含有脂肪酸蛋白特性(I)和DAG结合位点(II)等功能结构域. 预测三角褐指藻DGAT1蛋白为亲水性蛋白, 含有6个跨膜结构域和7个超强跨膜螺旋区, 无信号肽. 三角褐指藻与海链藻同属于藻类聚类为同一支. 光照强度和温度处理均显著促进三角褐指藻1基因的表达, 在光照强度为2500lx或温度为25℃时, 三角褐指藻1基因的表达量达到最大, 这与三角褐指藻总脂含量的变化趋势一致, 暗示DGAT1蛋白可能与三角褐指藻总脂生物合成与积累密切相关, 是总脂合成途径的关键酶之一.
[1] Gouveis L, liveira A C. Microalgae as a raw material for biofuels production[J]. Journal of Industrial Microbiology & Biotechnology, 2009, 36(2):269-274.
[2] Chisti Y. Biodiesel from microalgae[J]. Biotechnology Advances, 2007, 25(3):294-306.
[3] 马海明, 施启顺, 柳小春. DGAT相关基因研究进展[J]. 遗传学报, 2005, 32(12):1327-1332.
[4] 唐桂英, 柳展基, 单雷. 二酰基甘油酰基转移酶(DGAT)研究进展[J]. 中国油料作物学报, 2010, 32(2):320-328.
[5] Shockey J M, Gidda S K, Chapital D C, et al. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the[J]. Plant Cell, 2006, 18(9):2294-2313.
[6] Jako C, Taylor D C. Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltrans- ferase enhances seed oil content and seed weight[J]. Plant Physiology, 2001, 126(2):861-874.
[7] Lardizabal K, Effertz R, Levering C,et al. Expression ofDGAT2A in seed increases oil in soybean[J]. Plant Physiology, 2008, 148(1):89-96.
[8] Gómez-Loredo A, Benavides J, Rito-Palomares M. Growth kinetics and fucoxanthin production of, and, cultures at different light and agitation conditions[J]. Journal of Applied Phycology, 2016, 28(2):849-860.
[9] Jaeyon L, Chan Y, Soyoung J, et al. Comparison of several methods for effective lipid extraction from microalgae[J]. Bioresource Technology, 2009, 101(1):75- 77.
[10] 李书霞, 刘伟, 李威, 等. 玉米DGAT基因家族的全基因组分析[J]. 核农学报, 2015, 29(4):643-650.
[11] 刘贵芹, 邵群, 黄荣峰, 等. 大豆DGAT基因家族的鉴定和表达分析[J]. 中国农学通报, 2013, 29(12):55-61.
[12] 李书霞. 玉米和棉花DGAT基因家族的全基因组分析[D]. 杭州: 浙江大学, 2015.
[13] 刘凯. 油茶DGAT1基因的全长cDNA克隆与原核表达[D]. 长沙: 中南林业科技大学, 2012.
[14] 吕文兵, 刘红涛, 薛乐勋. 不同盐浓度和光照强度对杜氏盐藻基因表达的影响[J]. 华中师范大学学报(自然科学版), 2010, 44(2):288-292.
[15] 尹航, 袁岚瑛, 俞凯, 等. 三角褐指藻基因生物信息学及表达差异分析[J]. 核农学报, 2019, 33(8): 1483-1489.
[16] 华雪铭, 周洪琪, 丁卓平. 温度和光照对微藻的生长、总脂肪含量及脂肪酸组成的影响[J]. 上海海洋大学学报, 1999, 8(4):309-315.
[17] 叶丽, 蒋霞敏, 毛欣欣, 等. 温、光、盐对三角褐指藻紫外诱变株生长、总脂及脂肪酸的影响[J]. 生态学杂志, 2015, 34(2):454-462.
[18] 蒋霞敏, 郑亦周. 14种微藻总脂含量和脂肪酸组成研究[J]. 水生生物学报, 2003, 27(3):243-247.
[19] 吕军, 张颖, 冯立芹, 等. 生物信息学工具BLAST的使用简介[J]. 内蒙古大学学报(自然科学版), 2003, 34 (2):179-191.
[20] Gasteiger E, Hoogland C, Gattiker A, et al. Protein Identification and Analysis Tools on the ExPASy Server[M]//The Proteomics Protocols Handbook. Totowa, NJ: Humana Press, 2005:571-607.
[21] 余舜武, 刘鸿艳, 罗利军. 利用不同实时定量PCR方法分析相对基因表达差异[J]. 作物学报, 2007, 33(7): 1214-1218.
[22] Bligh E G, Dyer W J. A rapid method of total lipid extraction and purification[J]. Canadian Journal of Biochemistry & Physiology, 1959, 37(8):911-917.
[23] 马燕斌, 吴霞, 王霞, 等. 甘蓝型油菜基因表达的特异性分析[J]. 西北植物学报, 2013, 33(10): 1958-1963.
[24] 周丹. 大豆二酰甘油酰基转移酶(DGAT)基因克隆及其功能的初步研究[D]. 南京: 南京农业大学, 2012.
[25] Lung S C, Weselake R J. Diacylglycerol acyltransferase: A key mediator of plant triacylglycerol synthesis[J]. Lipids, 2006, 41(12):1073-1088.
[26] Guo Z, Cromley D, Billheimer J T, et al. Identification of potential substrate-binding sites in yeast and human acyl- CoA sterol acyltransferases by mutagenesis of conserved sequences[J]. Journal of Lipid Research, 2001, 42(8): 1282-1291.
[27] Weselake R J, Madhavji M, Szarka S J, et al. Acyl-CoA- binding and self-associating properties of a recombinant 13.3 kDa N-terminal fragment of diacylglycerol acyl- transferase-1 from oilseed rape[J]. BMC Biochemistry, 2006, 7(1):24-36.
[28] 郝敬云, 周广航, 邵雪梅, 等. 莱茵衣藻基因家族的鉴定与功能分析[J]. 分子植物育种, 2016, 14(9): 2343-2352.
[29] 张晓琼, 乔琳, 胡利宗, 等. 棉花与模式植物基因的鉴定与分析[J]. 江苏农业科学, 2016, 44(10):69- 73.
[30] 沈城, 刘楚吾, 刘丽. 温度胁迫及恢复初期稀杯盔形珊瑚共生虫黄藻、、、基因表达分析[J]. 热带海洋学报, 2016, 35(3):72-78.
[31] 周洪琪, Renaud S M, Pany D L, 等. 温度对新月菱形藻、铲状菱形藻和杷夫藻的生长、总脂肪含量以及脂肪酸组成的影响[J]. 水产学报, 1996, 20(3):235-240.
Bioinformation analysis and expression regulation of1 gene in
WANG Heyu1, GONG Yifu2,3*, ZHENG Xiaoyun2,3, LI Shenrui2,3
( 1.College of Food and Pharmaceutical Sciences, Ningbo University, Ningbo 315832, China; 2.School of Marine Sciences, Ningbo University, Ningbo 315832, China; 3.Key Laboratory of Marine Biotechnology of Zhejiang Province, Ningbo 315832, China )
In the current study, the full-length cDNA sequence of1 (GeneBank Accession Number: 7200924) inwas cloned by transcriptome sequencing. Bioinformatics analysis and expression difference were also studied. The results showed that the full length of the1 gene fromwas 1438bp, had an open reading frame (ORF) of 1098bp with the fatty acid-binding protein signature (I) and DAG binding site (II) domains, encoding 365 amino acids. The protein prediction results showed that the DGAT1 protein fromwas a hydrophilicity protein including 6 transmembrane structure domains and 7 super transmembrane helical regions, without signal peptide. Phylogenetic analysis results showed that DGAT1 protein fromhad closed homologous relationship with the DGAT1 protein of. RT-qPCR results showed that light intensity and temperature could significantly promote the expression of1 gene ofWith the increase of light intensity and temperature, the expression quantity of1 gene fromshowed an increased first and then decreased trend. The expression quantity of1 gene reached the maximum at light intensity of 2500 lx or temperature of 25℃. The expression of1 gene was consistent with the total fat content. The results indicated that DGAT1 protein might be one of the key enzymes in the biosynthesis and accumulation of total lipid in.
;1gene; cloning; bioinformatics; gene expression regulation
S917.3; Q943.2
A
1001-5132(2021)04-0001-07
2019−11−20.
宁波大学学报(理工版)网址: http://journallg.nbu.edu.cn/
浙江省科技厅重点科技创新团队项目(2012R10029-07, 2010R50029); 宁波市科技攻关项目(2014C91023, 2013C10018); 宁波市社发重大项目(2017C510002).
王何瑜(1982-), 女, 重庆江津人, 助理实验师, 主要研究方向: 藻类生物技术. E-mail: wangheyu@nbu.edu.cn
龚一富(1973-), 男, 重庆开县人, 博士/副教授, 主要研究方向: 植物次生代谢产物及机理. E-mail: gongyifu@163.com
(责任编辑 章践立)
猜你喜欢
古今农业(2022年1期)2022-05-05 06:58:42
粮食与食品工业(2021年3期)2021-06-18 07:50:48
中国生殖健康(2020年5期)2021-01-18 03:00:06
水生生物学报(2019年3期)2019-05-29 06:58:48
中国生殖健康(2018年5期)2018-11-06 07:15:56
环境保护与循环经济(2017年3期)2017-09-26 11:42:30
中国油脂(2017年7期)2017-09-16 04:28:48
中国洗涤用品工业(2015年8期)2015-02-28 19:02:49
水生生物学报(2014年2期)2014-05-27 08:07:30
食品工业科技(2014年23期)2014-03-11 18:19:31
