
用材树种高山栲潜在适生区分布区及影响因子*
2021-06-24 13:48张春华雷晨雨田瑞杰冯德枫孙永玉
西部林业科学 2021年3期
张春华,雷晨雨,田瑞杰,冯德枫,孙永玉
(1.中国林业科学研究院资源昆虫研究所,云南 昆明 650233;2.国家林业局云南元谋荒漠生态系统定位研究站,云南 昆明 650233;3.西南林业大学,云南 昆明 650224)
物种与环境的相互关系一直是生态学和生物地理学研究的热点问题[1-2],由于人口膨胀所导致的农业过渡开发及由此产生的物种生境退化、破碎化、外来物种入侵是造成物种分布面积减少甚至灭绝的直接或间接原因[3-4]。生境选择是进行物种种群恢复过程中最重要的环节之一[4],也是任何生态系统重建的先决条件[5-6]。物种潜在地理分布预测模型在保护生物学中具有重要意义[7-8],如入侵物种的传播[9-10]、物种多样性的空间格局[11],气候变化对植物分布的影响[12-13]。最大熵(MaxEnt)模型是起源于统计力学的一种物种潜在分布模型[14-15],较其它分布模型而言只需要物种存在(或发生)数据和环境信息[16-17]。
高山栲(CastanopsisdelavayiFranch.)为壳斗科(Fagaceae)栲属(Castanopsis)多年生常绿阔叶乔木树种,是亚高山松栎混交林的主要树种,散生于四川西南部、云南、贵州西南部海拔1 500~2 800 m的山地杂木林中,偶成小片状分布[18]。其心材黄棕色,材质坚硬,强度高,耐水湿,是较理想的桩、柱及建筑和家具用材。历史上,高山栲在云南分布广泛,但由于木材燃烧值高,加之生长区域内大范围的农垦生产,压缩了其生长、生活空间,很难见到大面积分布区。目前只在人迹罕至的横断山区、物种保护启动相对较早的森林公园、水源地、寺庙等神山圣水之旁。因此,了解和掌握影响高山栲适生环境分布的气象因子是其种群恢复的理论基础,可为其优良种质资源库建设选址提供理论依据。
1 材料与方法
1.1 环境及高山栲分布数据
用于分析高山栲适生环境分布的20个生物气候变量(表1)来源于WorldClim (www.worldclim.org),精度30 s(约1 km)[19]。依据《中国植物志》对高山栲的相关分类性状描述记载,于2018年9月至2019年9月对其在云南省的现有实际分布进行踏查。平坦地带每50~80 km 调查1次,山区地带则海拔每变化80~100 m 调查1次,将现存胸径大于60 cm 的个体且面积大于1 hm2的区域认定为其原生分布区,并应用精度为±5 m的多通道全球定位系统(GPS)记录地理坐标,共得到71个高山栲自然分布点位数据,在室内利用Excell整理成仅含物种名、分布坐标的CVS格式文件备用[20]。研究涉及的中国行政区划矢量地图(比例1︰4000000)从国家基础地理信息系统(http://bzdt.ch.mnr.gov.cn/index.jsp)下载得到。另外,在ArcGIS 13.0中完成中国行政区范围内的环境数据图层提取及高山栲在不同气候条件下的潜在分布图制作和各类适生等级区域面积估算。

表1 环境变量及贡献率Tab.1 Environment variable used in the study and their percentage contribution
1.2 空间模型构建
MaxEnt模型是一个基于最大熵的机器学习程序,可根据环境相似性估算物种分布概率[21-22]。首先采用Pearson相关系数(r)进行物种分布点间的多重共线性检验并剔除相关系数r>|±0.8|的变量[14],选择等温性(Bio3)、温度季节变化标准差(Bio4)、最冷月最低温(Bio6)、年均降雨量(Bio12)和海拔(Alti)作为构建高山栲潜在适生区分布的限制因子。其次选择70%、30%的分布数据分别用于机器学习和测试,最大叠代运行次数为100次,其它设置为系统默认值,并运用刀切法评估变量重要性[23]。最后,采用工作曲线下面积(AUC)评估模型的优良性,AUC值越大,模型精度越高[24]。并将高山栲的MaxEnt生态位模拟模型输出的分布概率P(0~1)划分为高适生区(P≥0.6)、中适生区(0.4≤P<0.6)、低适生区(0.2≤P<0.4)和非适生区(P<0.2)[25]。
2 结果与分析
2.1 模型精度检验及适生区划分
应用等温性(Bio3)、温度季节变化标准差(Bio4)、最冷月最低温(Bio6)、年均降雨量(Bio12)和海拔(Alti)等环境因子构建的高山栲适生区分布MaxEnt模型的平均训练AUC(Mean training AUC)=0.983以及平均测试AUC(Mean test AUC)=0.984,据MaxEnt 预测模型精度判定标准即AUC>0.9表明该模型能准确预测高山栲的潜在适生区,且精度和可信度较高。
研究结果表明等温性(bio3)、年平均降雨量(bio12)、温度季节变化方差(bio4)、最冷月最低温(bio6)和海拔(Alti)等5个环境因子是决定高山栲适生区分布的关键因素,也是其适生区分布模型的主要构建因子(表1)。它们对模型的贡献率分别为44.5%、19.7%、23.5%、8.8%和2.6%。Jackknife检验(图1)表明,温度季节变化方差(bio4)和昼夜温差与年平均气温比值(Bio3)对标准训练增益率达2.61、2.63,说明它们单独使用时能比其它环境因子包含更有用的环境背景信息。年平均降雨量(bio12)和最冷月最低温(bio6)均有一定的增益,而海拔(Alti)在单独使用时增益较低,表明它本身没有包含很多信息。环境因子一致性或趋同性是植物引种或在未知区域寻找目标物种可能存在的前提条件,根据环境因子响应曲线(图2),高山栲适生区环境条件为:海拔1 400~2 800 m、年平均降雨量900~1 260 mm、等温性1.2~1.3、温度季节变化标准差27~46 ℃、最冷月最低温0~6.7 ℃。其中海拔2 000 m、昼夜温差与平均温比值1.250、温度季节变化标准差40 ℃、最冷月最低温4 ℃、降雨量1 100 mm是高山栲最佳适生环境。
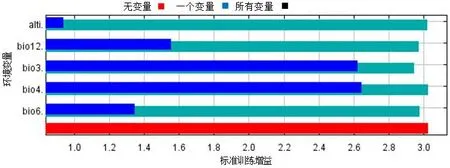
图1 刀切法检验环境因子对标准训练增益率影响Fig.1 Jackknife test of variable importance to regularized training gain
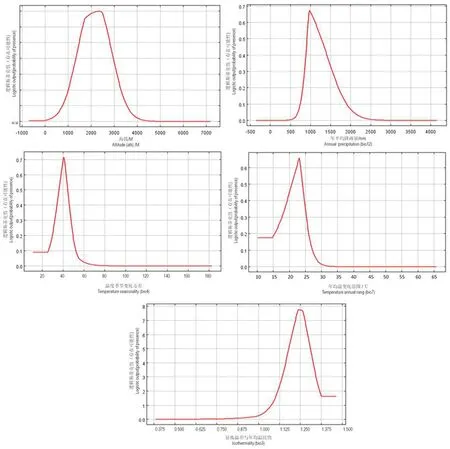
图2 环境因子响应曲线Fig.2 Response curves of environment variables
2.2 高山栲当代适生区
研究结果显示高山栲当代(1950—2000年)总适生区面积(表2)为12 741.98×104hm2,占中国陆地国土面积的13.27%,主要分布于西藏东南部-四川南部及中部—贵州—湖南—江西—浙江一线以南的海拔1 400~2 800 m山地。其中,低度适生区面积为9 719.75×104hm2,占总适生区面积的76.28%;中度适生区面积为2 852.22×104hm2,占总适生区面积的22.38%,集中分布于云南和四川南部;高度适生区面积为170.01×104hm2,占总适生区面积的1.34%,集中分布于云南。
2.3 高山栲末次间冰期和末次盛冰期适生区
高山栲在末次间冰期(~120—140 Ka BP)的适生面积(表2)较当代稍有增加,为14 188.09×104hm2,分布范围与当代基本一致。其中,低度适生区面积为1 1251.87×104hm2,占总适生区面积的79.30%;中度适生区面积为2 426.19×104hm2,占总适生区面积的17.10%,分布区较当代增加了海南分布区;高度适生区面积为510.04×104hm2,占总适生区面积的3.40%,面积近当代的3倍。

表2 不同年代高山栲分布面积预测Tab.2 Predicted suitable habitats for C.delavayi in different period ×104 hm2
高山栲在末次盛冰期(~21 Ka BP)的适生区面积(表2)10 892.84×104hm2,较当代萎缩了14.51%。其中,中度适生区面积2 253.17×104hm2,占总适生区面积20.68%,主要分布于云南;低度适生区面积8 638.66×104hm2,占总适生区面积79.32%。
2.4 高山栲未来的适生区预测
高山栲在2050年、2070年气候条件下的适生区分布范围与当代基本一致,适生区总面积较当代有所减少。总适生区面积分别较当代减少了425.03×104hm2和125.01×104hm2(表2)。
3 讨论与结论
3.1 讨论
(1)影响高山栲地理分布因素 本研究以高山栲当代现存分布数据和环境背景数据为基础,应用MaxEnt最大熵模型推演不同气候条件下的地理分布概况。客观地获得了高山栲在不同气候条件下的适生区潜在分布信息及变化。研究得到的高山栲在当代的适生区分布于西藏东南部—四川南部及中部—贵州—湖南—江西—浙江一线以南与其实际分布区[17]高度一致;另据Swets提出MaxEnt最大熵模型预测精度判定标准(AUC>0.9表明该模型具有较好预测精度)[24],由此推断该模型推演得到的不同气候条件下高山栲潜在适生区分布具有较高的可信度和准确性。
在构建高山栲适生区分布模型的因子中,等温性对预测模型的贡献率最高达44.5%,是第1因子影响因子;温度季节变化是第2影响因子,贡献率为23.5%;年平均降雨量是第3影响因子,贡献率为19.7%;最冷月最低温和海拔分别为第4和第5影响因子,贡献率依次为8.8%和2.6%。并依据环境因子响应曲线得到高山栲分布区气候条件为:海拔1 400~2 800 m、年平均降雨量900~1 260 mm、等温性1.2~1.3、温度季节变化标准差27~46 ℃、最冷月最低温0~6.7 ℃。在云南省海拔1 400~2 800 m区域均能满足高山栲对年降雨量、昼夜温差与年平均温比值、温度季节方差、最冷月最低温的要求,可根据海拔确定其适生区。
(2)高山栲在末次间冰期和末次盛冰期适生区分布 在距今12~14×104a的间冰期,我国境内多年平均温度与当代相似,而距今2.1×104a前的盛冰期较当代低12 ℃左右,剧烈气候波动是导致植物适生区分布区变迁的最主要因素[26]。表现为低温胁迫使植物南迁或迁移至低海拔区域的“冰期避难所”,温度升高其适生区又会北移或向高海拔迁移。间冰期的温度与当代相似且导致了高山栲的适生区面积略高于当代;但是进入盛冰期,高山栲适生区面积较当代缩减了1 849.14×104hm2,其中最明显的变化是适生区南移明显,最南端出现在海南的西海岸。
(3)高山栲在未来气候条件下适生区分布 在全球变暖的大背景下[27],对高山栲未来几十年的适生区变迁预测结果表明,在分布区范围不变的前提下其适生区面积有一定幅度减少,但降幅较小(小于4%),这有利于该物种的生长繁衍[1,9-10,13]。
(4)高山栲种质资源保存库选址 高山栲在不同气候条件的潜在适生区分布和面积变化较小,这说明高山栲对全球气候变化不敏感。而其中度以上适生区绝大部分分布于云南省,这可能与云南地处北亚热带和多变的山地气候有关[28]。因此,云南横断山区、滇中高原海拔1 400~2 800 m区域是高山栲理想的生活繁衍栖息地,也是其种质资源异地保存库建设首选址场地。
3.2 结论
研究应用MaxEnt最大熵模型得到高山栲在不同气候条件下在我国境内的潜在适生区分布模型,未来气候变化对其适生区面积影响较小,且90%以上的中高度适生区分布于云南省海拔1 400~2 800 m区域,这为高山栲种群恢复、针叶树种纯林的结构调整及生态功能提升提供了理论依据。同时指出云南省是高山栲优良单株和异地种质资源保存林建设的理想区域。
猜你喜欢
西部林业科学(2022年6期)2023-01-03
西北植物学报(2022年3期)2022-04-18
治淮(2022年4期)2022-01-01
数学小灵通·3-4年级(2021年6期)2021-07-16
植物资源与环境学报(2020年2期)2020-04-20
军事文摘(2018年24期)2018-12-26
竹子学报(2018年2期)2018-10-25
今日重庆(2016年2期)2016-08-11
西部广播电视(2015年7期)2016-01-16
中国科技术语(2011年3期)2011-12-31
