
云南万掌山林场思茅松林分结构特征*
2021-06-24 13:48苏俊武刘永刚陈强刘云彩孙志刚李先泽鲁兴武槐可跃周红
西部林业科学 2021年3期
苏俊武,刘永刚,陈强,刘云彩,孙志刚,李先泽,鲁兴武,槐可跃,周红
(1.云南省林业和草原科学院,云南 昆明 650201; 2.普洱市思茅区万掌山林场,云南 普洱665000;3.保山市昌宁县西山国社合作林场,云南 昌宁 678100)
思茅松(Pinuskesiyavar.langbianensis)是云南特有的乡土树种,天然分布于云南热带北缘和亚热带南部半湿润地区,分布的海拔范围一般为700~1 800 m,而多集中分布于1 100~1 700 m之间。思茅松是脂、材兼用树种[1],具有重要的经济价值和生态价值,其人工林在20世纪80年代后得到快速的发展,面积达10.56×104hm2以上[2],在其集中分布区的经济和社会发展中占有举足轻重的地位。
目前,有关思茅松的研究主要集中在良种选育[3-8]、无性扩繁[9-10]、丰产栽培[11-13]、病虫害防治[14-15]、松脂利用[16]等方面,对于其林分结构的研究多集中于天然次生林,如:胥辉等[17]利用Normal、Weibull、Beta和Gamma分布函数拟合并预测思茅松天然次生林的直径分布,结果显示Weibull分布在思茅松天然次生林直径分布的应用中效果最佳;欧光龙等[18]利用Weibull函数拟合思茅松天然林林分直径、用幂函数模型拟合树高结构,结果直径符合Weibul函数分布,树高符合幂函数分布;李超等[19]在思茅松天然林林分直径大小多样性及环境解释研究中采用Shannon指数、Simpson指数对林分大小多样性进行量化排序分析,表明林分直径大小多样性随立地条件和环境因子变化。针对思茅松人工林林分结构特征方面的研究仅见张文勇[20]对思茅松人工林开展过直径分布的研究。林分结构是林分功能的基础和表现[21],是森林经营和分析中的重要因子,也是实施森林经营活动的具体对象[22-23],分析林分结构有助于人们了解森林的发展历史、现状和生态系统将来的发展方向[24]。本文通过对思茅松人工幼、中龄林林分结构进行研究,以揭示思茅松人工林幼、中龄林林分直径、树高、蓄积结构状态和演变规律,有助于预判林分生长趋势、确定林分间伐强度和调整林分结构。
1 材料与方法
1.1 研究区概况
万掌山林场(22°33′7″~23°0′23″N,100°29′54″~101°6′56″E)位于云南南部普洱市思茅区,现有林地面积17 440 hm2,林场管护区内最高海拔2 010 m,最低海拔570 m,相对高差1 440 m。该区域属低纬山地南亚热带高原性季风气候区,年平均气温19.4 ℃,最冷月平均气温10.5 ℃,最热月平均气温22.7 ℃,≥10 ℃活动积温6 302 ℃,无霜期318 d;年均降雨量1 340.9 mm,相对湿度87%,雨水充足,气候湿润。思茅松林大多数为纯林,伴生树种在林分内所占的比重小,林中常见的树种主要有红锥(Castanopsishystrix)、短剌栲(C.echidnocarpa)、杯状栲(C.calathiformis)、西南木荷(Schimawallichii)、西南桦(Betulaalnoides)等,林下多见密花树(Myrsineseguinii)、银柴(Antidesmaspp.)、余甘子(Phyllanthusemblica)、算盘子(Clochidionspp.)、水锦树(Wendlandiaspp.)、岗柃(Euryaspp.)、野牡丹(Melastomaspp.)、斑鸠菊(Vernoniaspp.)、粽叶芦(Thysanolaenamaxima)、山菅兰(Dianellaensifolia)、白花蛇舌草(Hedyotisdiffusa)、紫茎泽兰(Ageratinaadenophora)等植物。土壤主要为赤红壤、红壤和紫色土。
1.2 样地设置与调查
2018年对云南省普洱市思茅区万掌山林场8、11、15 a思茅松人工林进行调查,在3种林龄的思茅松林分中各设置15块样地,共计45块,样地大小为20 m×20 m。调查记录每块样地的基本情况,包括海拔、坡向、坡度和土壤等因子;对样地内林木进行每木检尺,实测胸径、树高、冠幅和郁闭度等因子。用围尺测量胸高直径,精确至0.1 cm;连接多根短测杆以延长其高度并举至与树同高,计算测杆总长作为树高,精确到0.1 m;用皮尺分别测量南、北向冠幅,精确到0.1 m;采用目测法调查郁闭度,记载到小数点后2位。
1.3 数据处理
利用各样地调查数据,计算其胸径、树高的平均数、标准差、变异系数以及偏度、峰度,以及各样地株数密度、郁闭度、蓄积量等。
胸径以2 cm为间距,采用上限排外法划分径阶,并按径阶统计株数、蓄积量(m3/hm2)、平均单株蓄积(m3/株)、累积蓄积量(m3/hm2)等。
以林分平均直径1个径阶以上的林木为优良木,平均直径1个径阶以下的林木为被压木进行量化分级,优良木为5级分类法中的Ⅰ、Ⅱ、Ⅲ级木,被压木为Ⅳ级和Ⅴ级木。按林木分级类别统计其株数、蓄积量。
调查数据采用Excel进行统计分析和柱状图、散点图的绘制。采用思茅松二元材积公式[25]V=0.000 075 592 9D1.941 31H0.823 88计算单株材积。
2 结果与分析
2.1 幼、中龄林林分郁闭度
调查样地分布于海拔1 300~1 500 m之间,处于不同的坡位和坡向,所有样地林分郁闭度都在0.7以上,其中郁闭度在[0.70~0.80]区间的林分占调查样地数40%,郁闭度0.80以上的占调查样地数60%,林分生长正常。各林地抚育措施不尽相同,现有保存密度、郁闭度也存在差异: 8、11 a林分密度相对较大,期间未进行过人为干预,其郁闭度0.80以上样地占比分别为80%、100%,表现出林龄大的林分其郁闭度大的特点;15 a林分幼林期曾进行过抚育间伐(间伐强度为20%),现林分郁闭度相对较小,其郁闭度0.80以上样地占比为0%(表1)。

表1 林分郁闭情况Tab.1 Forest canopy closure 块
2.2 直径结构特征
林分直径分布是林分中林木株数在径阶上的分配,是最重要、最基本的林分结构。思茅松幼、中龄林林分直径分布范围较大,立木最小胸径为2 cm,最大胸径为24.2 cm。从直径分布特征因子(表 2)可看出:8 a思茅松林分平均直径为9.7 cm,最大胸径为17.3 cm,最小胸径为2 cm,变异系数为23.51%,直径偏度系数为-0.077 2,峰度系数为-0.034 6;11 a思茅松林分平均直径为9.3 cm,最大胸径为18.3 cm,最小胸径为3 cm,变异系数为23.50%,直径偏度系数为0.196,峰度系数为-0.009 3;15 a思茅松林分平均胸径为12.5 cm,最大胸径为24.2 cm,最小胸径为5.6 cm,变异系数为22.26%,直径偏度系数为0.213 2,峰度系数为0.055 7。从3种不同的幼、中龄林林分直径的变化来看,其极差在15.3~18.6 cm之间,变异系数在22.26%~23.51%之间,各林分的极差和变异系数均较大。

表2 林分直径分布特征因子Tab.2 Characteristic factors of stand diameter distribution
思茅松人工林幼、中龄林林分中包含径阶株数序列的特征,是以林分平均直径为中心的概率对称分布,其林分直径分布接近正态分布(图1)。8 a思茅松人工林直径分布曲线呈单峰左偏(偏度<0),峰值出现于平均直径9.7 cm的位置,并以此为中心向两侧递减,且株数下降较快,曲线较陡(峰度<0);11 a林分直径分布曲线呈单峰右偏(偏度>0),峰值为9.3 cm,并以峰值为中心向两侧株数递减,且下降较快,曲线较陡(峰度<0)。15 a思茅松人工林直径分布曲线呈现单峰右偏(偏度>0)且峰值偏移至大径阶区,在平均直径12.5 cm时达到最大,并以此为中心向两侧逐步递减,但曲线下降较平缓(峰度>0)。8、11 a林分以8、10、12 cm径阶的林木个体数量最多,15 a林分以10、12、14、16 cm径阶的林木个体数量最多,林木数量向中心径阶两侧依次递减。

图1 思茅松幼、中龄林林分直径分布注:¯D为林分平均直径Fig.1 Frequency distribution of diameter classes in young and middle-aged forests of P.kesiya var.langbianensis


表3 思茅松幼、中龄林不同直径林木百分数Tab.3 Proportion of number of stands in young and middle-aged P.kesiya var.langbianensis forest
2.3 树高结构特征
林木个体受遗传品质和立地条件的影响,高生长速度不同,形成树高大小差异及不同层次的林分。幼、中龄林中优良木占林分株数的85.8%~89.6%,被压木占林分株数的10.4%~14.2%。8、11 a思茅松幼、中龄林优良木(Ⅰ、Ⅱ、Ⅲ级木)径阶≥8 cm,立木处于林冠上层,分别占林分株数的88.8%、85.8%;处于林冠下层的被压木(Ⅳ、Ⅴ级木)径阶<8 cm,分别占林分株数的11.2%、14.2%(表4)。15 a思茅松优良木(Ⅰ、Ⅱ、Ⅲ级木)径阶≥10 cm,占林分株数的89.6%,立木处于林冠上层;被压木(Ⅳ、Ⅴ级木)径阶<10 cm,占林分株数的10.4%,立木处于林冠下层,呈被压状态(表4)。在3种林龄思茅松林分中,直径小的林木树高生长量也小,随着林木直径生长的增加,树高相应地增大(图2);思茅松林木树高与直径呈线性正相关(8、11、15 a林分其R2分别为0.974 4、0.976 9、0.946 3),树高分布亦如直径分布一样近似于正态分布。
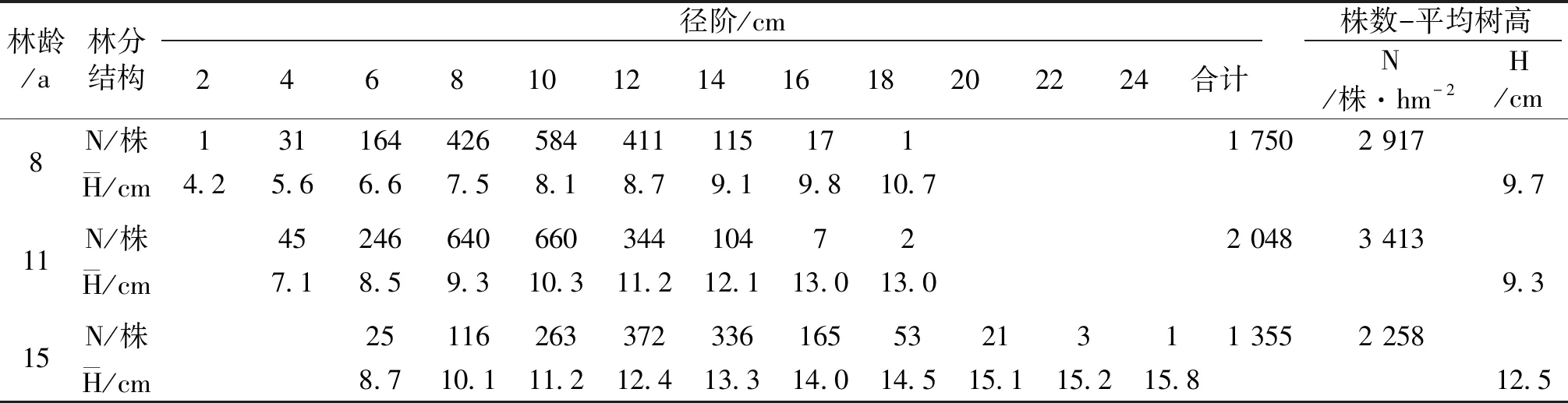
表4 各林分“株数-树高”结构状态Tab.4 “Number of plants-tree height” structure status of various forest stands
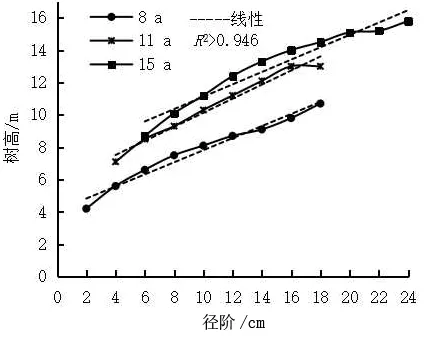
图2 思茅松幼、中龄林林分树高曲线注:¯D为林分平均直径Fig.2 The curve of tree heights of various diameter stages of P.kesiya var.langbianensisyoung and middle-aged forests
2.4 株数-蓄积量结构特征
统计结果(表5、表6)显示,思茅松幼、中龄林林分的蓄积量69.2%~72.0%分布在平均直径以上的上层林木中,其余28.0%~30.8%分布在平均直径以下的林木中。随着林龄的增长,密度大的林分下层被压木数量增大,林分中产生的死亡木也增多,这种特点在现有保存密度较大的11 a林分中尤为明显。林分蓄积的生长量主要产生于上层林木(Ⅰ、Ⅱ、Ⅲ级木),被压木(Ⅳ、Ⅴ级木)立木蓄积量仅占3.2%~4.8%。3种林分中材积的增长和积累不是均匀分布在各林木个体和各径阶中,而是形成各径阶“株数-蓄积量”的非均衡结构。在3种林龄林分中,随着径阶增大,平均单株材积遵乘幂函数(8、11、15 a林分其R2分别为0.997 9、0.998 7、0.999 9)单调递增,但单株材积太大或太小的林木株数都较少(图3a)。蓄积在不同径阶中的分布则近似于钟形,但与直径分布相比其峰值的出现都有所滞后,8、11、15 a林分蓄积量最大值分别出现在12、10、14 cm径阶(图3b),这是因为蓄积除受与之呈正相关的单株材积影响外,还受随径阶增大而减少的林木株数影响。随径阶增大,蓄积量累积速度呈现“慢-快-慢”的规律性变化,蓄积累积曲线呈“S”形(图3c)。

表5 各林分“株数-蓄积量”的结构状态Tab.5 The structure status of “number of plants-stock volume” of various forest stands
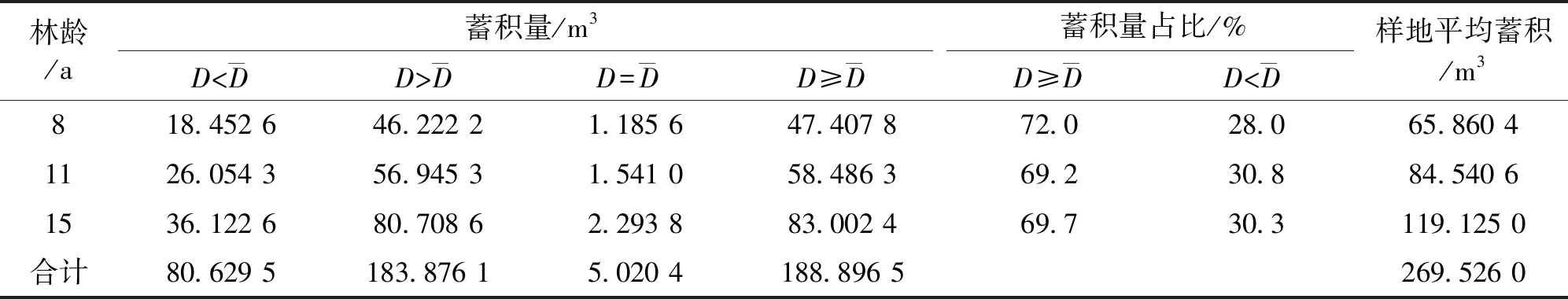
表6 思茅松幼、中龄林林分不同类型林木蓄积量及其占比Tab.6 Proportion of stand volume of P.Kesiya var.langbianensis young forest and middle-age forest



图3 思茅松幼、中龄林林分各径阶的蓄积分布注:¯D为林分平均直径
3 讨论与结论
森林结构泛指不同植物种类和大小的空间配置与分布,而林分结构则是森林结构的基本单元[23]。 森林结构由空间结构(林木的点格局及其属性的空间分布)和非空间结构(林分密度、树种组成、直径分布、树高分布、树种多样性及林分活力等)构成,非空间结构的分析方法均以指数即单值形式表达林分单方面的结构特征,描述的是林分特征的一种平均状态,这种状态常与个体空间属性无关,即忽略了林分中个体大小的变化以及不同树种在林地上的空间分布[26]。这些简单的平均数量特征虽然很难对复杂的森林结构做出较为完整的描述和判断,但是因其数据较易获取,同时很容易与森林经营活动相联系,因而在森林经营管理中被广泛使用。尤其是对于人工林纯林来说,其空间结构特别是林木大小的水平、垂直分布变异性较小,明显有别于其它森林,可用林分非空间结构变量来表征人工林纯林林分特征的平均状态,以此作为制订森林经营管理措施的参考。
45块思茅松人工林样地皆为纯林,林层较为简单,由上、下2个亚林层构成,优良木处于林分上层,被压木处于林分下层。林分平均直径以下的林木株数与平均直径以上的株数总的来说大致各占50%。径阶株数序列特征是以林分平均直径为中心的概率对称分布。林分蓄积结构中,下层木蓄积量占比小,低于林分蓄积量的30%,上层木产生的蓄积量大,可占林分蓄积量的70%以上。采用径阶分级“刻划”思茅松林分株数-蓄积量结构,能较好地描述林分蓄积量结构状态。这与张裕农等[27]研究结果不尽相同,不同之处主要是因为平均直径取值不同所致,本研究中为算术平均值而非林分平方平均直径。
8、11、15 a思茅松人工林其树高生长量、蓄积量随林龄增加而增加(图2、图3c);平均单株材积亦随林龄增加而增加(8、11、15 a林分其值分别为0.037 6、0.041 3、0.087 9 m3);胸径生长在不同林龄、密度、立地条件的林分间皆有差异,且其生长受密度效应影响较大,表现为11 a林分平均胸径小于8 a林分。3个林龄思茅松人工林的生长速度都较快,这是因为思茅松较为速生的生长特性使然,按10 a龄级划分,3种林分正处于幼龄林后期至中龄林中期,此期间思茅松林木生长速度处于快速增长时期。
林木个体受遗传、立地条件和种植密度等因素的影响,生长过程中树高、胸径产生大小差异,在林分中形成优良木或被压木,优良木在生长过程中总是不断地分化和演替,或分化为被压木生长衰退而死亡[28],其数量随时间的转移而产生变化,这就是林木的自然稀疏过程。对人工林进行适时抚育间伐,有利于促进优良木生长,形成整齐林相,保持良好的林分卫生条件,减少病腐木,提高人工林质量与产量。8、11 a未间伐林分在幼、中龄林交接期郁闭度较高,林木生长竞争激烈、个体分化程度大,宜及时开展抚育间伐,这与刘永刚等[29]研究结果相一致。
密度与林木生长、林分蓄积结构相互关联。幼林期造林密度对林木胸径、树高等生长作用不太明显,但随着林龄的增加,林木之间的生长竞争日趋激烈,林分密度对林木生长的影响逐渐增加。在同等条件下,胸径随密度的增加而减小[30],密度大的林分年平均直径生长量小[31],不同密度对思茅松树高、胸径和单株材积影响极显著[32]。8、11、15 a林分间其树高、胸径、材积生长差异充分反映了密度效应作用下的思茅松生长特点:3种密度的林分中,密度从大到小排序为11 a(3 413株/hm2)、8 a(2 917株/hm2)、15 a(2 258株/hm2),平均胸径、树高及平均单株材积从大到小排序皆是15、8、11 a,表现出与密度的负相关效应。此外,随着林龄的增加,林木之间的生长竞争日趋激烈,林分密度对蓄积增长的影响逐渐增加。这种影响主要体现在单株材积随径阶变化的乘幂式增长导致的林木大小分化及被压木淘汰后林木株数的减少,但总体上遵循株数递减、总蓄积“S”形增长的规律,这种规律性变化至少能保持到成熟林时期。
除密度因素外,人为干预如幼林期施肥对林木树高、直径生长影响也较为显著[33],揭示了除密度调控手段外,可通过林地施肥以促进思茅松林木生长。
综上所述,8、11、15 a 思茅松人工林正处于快速生长期,但分别处于不同的林分生长状态:15 a林分曾进行过1次抚育,通过抚育间隔期的生长,其林木近似对称地分布在平均胸径两侧,林木虽有分化但不剧烈;8 、11 a林分林木分化较15 a林分剧烈,尤其是11 a林分,其林木分化剧烈,被压木生长量甚微,濒临死亡,另外,8 a林分亦处于分化调整期,林木开始剧烈分化,宜对此2种林龄的林分及时开展抚育间伐。
猜你喜欢
临床检验杂志(电子版)(2020年1期)2020-04-03
锦绣·下旬刊(2020年12期)2020-01-03
新农民(2019年9期)2019-02-19
农村经济与科技(2018年15期)2018-05-14
森林工程(2018年5期)2018-05-14
绿色科技(2017年16期)2017-09-22
现代农业科技(2017年12期)2017-07-29
学苑创造·B版(2015年12期)2016-06-23
林业与生态(2016年2期)2016-02-27
中南林业科技大学学报(2014年12期)2014-12-27
