
牛肝菌液态发酵及活性成分诱导培养研究进展
2021-06-21 15:54范方宇
食品工业科技 2021年8期
郭 磊,范方宇,刘 云,阚 欢
(西南林业大学生命科学学院,云南昆明 650224)
牛肝菌科(Boletaceae)真菌属真菌界(Fungi)、担子菌亚门(Basidiomycotina)、层菌纲(Hymenomycetes)、伞菌目(Agaricates),它既是高营养价值的真菌,也具有多种生理功效[1]。牛肝菌在全世界范围内都有分布,在欧洲的瑞典、法国,美洲的美国,亚洲的马来西亚、日本等国都有大量文献记录,中国已知牛肝菌有28 个属387 个种(变种),主要分布在云、贵、川、粤等地区[2-3]。牛肝菌属外生菌根真菌,对寄主有一定的选择性,很难实现人工栽培[4],故液态发酵培养将是以后牛肝菌研究的重要方向。
菌丝是牛肝菌液态发酵过程的重要产物,受各种发酵因素如培养基组成与比例、初始pH、发酵温度及发酵时间等的相互影响,牛肝菌菌丝发酵体系的流变特性也随之改变,进而影响到质量、热量和动量的传递[5],最终会影响到菌丝和发酵液中营养成分的生成和效率。同时,真菌液态发酵会产生多糖、色素、三萜类化合物及黄酮类化合物等次生代谢产物,具有抑菌、抗氧化、抗肿瘤、促进植物生长等作用[6]。有些次生代谢产物结构新颖,生物活性独特,是活性化合物研究的重要来源,且在抗肿瘤治疗上具有不可替代的作用,近年来所发现的新结构、新活性次生代谢产物超过50%来源于真菌[7]。另外,通过物理、化学及生物外源诱导手段可以提高牛肝菌液态发酵过程中目标生物活性成分的含量,生产过程具有可再生性,且不受自然环境或资源限制,已成为牛肝菌资源高效利用的创新研究方向。
1 牛肝菌科真菌菌丝的液态发酵培养基研究
真菌培养基是供真菌菌丝生长繁殖或积累代谢产物的营养基质,根据菌丝对营养物质的需求来配制,培养基一般有碳源、氮源、无机盐、生长素及水分等物质。不同培养基的成分是不同的,其中任何一种成分或含量的改变都会影响到菌丝的生长繁殖,各种培养基的成分与比例对菌丝的生长都会有较大影响。在真菌液态发酵过程中,除发酵培养基配方的影响外,初始pH、发酵温度、发酵时间及转速等也是影响真菌菌丝生长繁殖和代谢的重要指标。不同牛肝菌科真菌的液态发酵条件及菌丝生长情况见表1。

表1 不同牛肝菌液态发酵培养条件及菌丝生长情况Table 1 Liquid fermentation conditions and hyphae growth state of different Boletaceae fungi
通过优化培养基组成及发酵条件,牛肝菌菌丝体在营养素含量上远高于子实体,有着更高的营养价值保健功能,具有非常大的发展潜力。美味牛肝菌号称“牛肝菌之王”,是世界著名的食药用真菌[19],冮洁等[20]使用马铃薯200 g/L、葡萄糖25 g/L、蛋白胨7 g/L、K2HPO41.5 g/L、CaCl20.005 g/L 在初始pH5.0 时制备美味牛肝菌液态培养基,研究发现菌丝体蛋白质含量(28.40%)高于子实体(25.10%),菌丝体氨基酸总量(22.56%)也高于子实体(17.93%)。通过优化牛肝菌液态发酵的各种条件,能够在短时间内产生大量的菌丝体,克服了人工栽培难度大和对给生态环境带来的破坏,同时也能够获得和子实体相似的营养成分和代谢产物,具有广阔的市场发展前景。
2 牛肝菌中生物活性成分的液态发酵培养
牛肝菌含有多糖、萜类、甾体、色素及生物碱等多种成分,具有抗氧化、抗肿瘤和抗菌等多种生物活性,是安全、有效、低毒的天然药物分子来源,在疾病治疗上有着较大的发展潜力[2]。目前,牛肝菌生物活性成分的获取多来自于子实体,受自然资源条件的限制,原料来源有限且产量少,而液态发酵可以大量生产目标代谢活性成分,具有可再生性,有重要的科学研究意义。
2.1 胞外多糖
真菌胞外多糖(exopolysaccharides,EPS)具有高黏性、抗氧化、抗病毒和抗肿瘤等多种生物学活性[21]。但目前对牛肝菌发酵产EPS 的研究较少(见表2),如何获得并通过真菌诱导提高牛肝菌EPS 产量已经是科研人员新的研究方向。
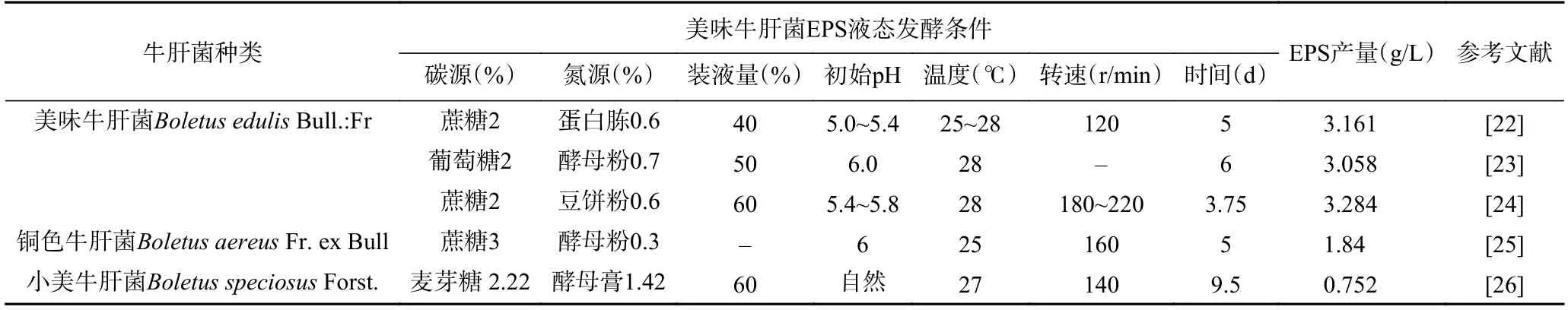
表2 牛肝菌EPS 发酵培养条件Table 2 Fermentation conditions of EPS from Boletaceae fungi
杨玉红等[23]对美味牛肝菌EPS 发酵因素研究后发现,影响EPS 产量较大的三个因素分别是葡萄糖、硫酸锰和磷酸二氢钾。在EPS 生产过程中,除通过控制发酵培养条件外,提取及纯化技术同样是提高EPS 产量及质量的关键因素。矫丽曼[27-28]研究了美味牛肝菌EPS 的提取纯化工艺及单糖组成,结果表明超声波协同酶法提取EPS 优于热水法、超声波法、酶法及超声波协同酶法对美味牛肝菌菌丝多糖的提取效率,EPS 提取率达到8.315%。采用DEAE 纤维素柱和G-100 葡聚糖凝胶柱对其纯化后发现EPS 是由D-木糖、D-甘露糖和D-半乳糖组成,摩尔比为3.1:63.3:1.0。阚国仕等[29]通过抗氧化活性研究发现美味牛肝菌EPS 对超氧阴离子自由基和羟基自由基有显著的清除能力,还能够抑制小鼠肝组织中丙二醛的生成、肝组织自发性脂质过氧化及减少红细胞氧化溶血。许春平等[25]分析了脱蛋白后铜色牛肝菌EPS 的体外抗氧化活性,研究得出EPS 对ABTS 自由基、DPPH 自由基及超氧阴离子自由基具有很好的清除能力。
真菌EPS 生产周期短,不受季节、地域和病虫害等限制,已作为增稠剂、乳化剂及稳定剂在食品工业中得到广泛应用,并作为血浆替代品、抗肿瘤、降血糖及降血脂药物在医药行业中有着非常广泛的应用[30]。
2.2 色素
真菌色素是真菌发酵产生的次生代谢产物,其生产安全性高且易于大规模生产,具有广阔的市场的发展前景[31-32]。在过去的二十年里,关于陆地和海洋生态系统中微生物色素有了深入的研究,包括微生物色素和癌症相关性的研究[33]。到目前为止,微生物,包括细菌、真菌和藻类,都已经被证明是天然色素的良好替代来源。方琼等[34]通过Plackett-Burman 试验筛选出影响远东疣柄牛肝菌(Leccinum extremiorientale(L.Vass.)Sing.)发酵产色素的影响因素主要是装液量和培养时间,并通过响应面设计优化出色素的最佳发酵条件是麦芽糖20 g/L、玉米浆干粉5 g/L、KH2PO43 g/L、MgSO4·7H2O 1.5 g/L、VB10.01 g/L、培养温度27 ℃、培养时间141.85 h、装液量87.70 mL,此条件下发酵液的吸光度为0.532,比优化前提高了11.37 倍。
真菌的次生代谢物色素是食品天然色素和食品添加剂的良好来源,包括红曲黄素(ankaflavin)、β-胡萝卜素(β-carotene)、角黄素(canthaxanthin)、核黄素(riboflavin)、番茄红素(lycopene)、红斑素(rubropunctatin)及黑色素(melanin)等,具有抗氧化、抑菌、抗癌及抗细胞毒性等多种生理功能[35-36]。目前,虽然对牛肝菌科真菌发酵产色素的研究相对较少,但由于其安全性高、生产周期短且具有较高的营养价值和药理活性,必将成为牛肝菌科真菌中天然色素生产的新途径。
2.3 其他
三萜类化合物(triterpenoids)是真菌中除了多糖外最重要的生物活性成分,是由数个异戊二烯去掉羟基后首尾相连构成的萜类化合物,具有预防心脑血管疾病、增强免疫力及抗肿瘤等多种生理功能[37-38]。近年来已对灵芝(Ganoderma lucidum(Leyss.ex Fr.)Karst.)[39]、粗毛纤孔菌(Inonotus hispidus(Bull.)Pat.)[40]及桦褐孔菌(Fascoporia oblique(Pers.Fr.))[41]等药用真菌液态发酵菌丝的三萜类化合物进行了提取和生物活性研究。但截止到目前,对牛肝菌子实体及发酵菌丝中的三萜类化合物都没有相关报道,因此对于牛肝菌三萜类化合物的发酵培养、诱导合成及生物活性研究是牛肝菌活性成分研究和资源开发利用的重要方向。
壳聚糖(chitosan)是唯一的天然碱性阳离子多糖,因其独特而优越的成膜性、抗菌性、生物相容性及安全性等而受到国内外学者的广泛关注,已应用到食品、药品和化妆品等多个行业[42-43]。李昊[44]筛选出葡萄糖和酵母浸出粉是影响美味牛肝菌发酵产壳聚糖的主要因素,通过响应面优化出最佳的发酵条件是葡萄糖30.53 g/L、酵母浸出粉7.02 g/L、温度28 ℃,装液量77.42 mL,初始pH 5.64,发酵6 d,此条件下壳聚糖产量可达3.81 g/L,占菌丝体干重的15.43%,且具有较高的脱乙酰度和纯度,对大肠杆菌也具有较好的抑制活性。
L-茶氨酸(L-theanine)是茶叶中特有的非蛋白氨基酸,也是茶叶的重要活性成分,因其具有抗氧化、改善情绪与认知及减轻焦虑等生理作用,于2014 年被我国批准其为新食品原料[45-46]。L-茶氨酸也存在于牛肝菌科真菌中,李健[47]通过毛细管电泳分析出褐绒盖牛肝菌(Xerocomus badius(Fr.)Kiihner ex Gilb.)、粘盖牛肝菌、黄柄牛肝菌和茶褐牛肝菌(Boletus brunneissimusChiu)的L-茶氨酸含量分别是29.86、27.35、25.61 和15.60 mg/100 g,并研究获得了褐绒盖牛肝菌液态发酵产L-茶氨酸的最佳条件是29.17 g/L 葡萄糖、6.38 g/L 酵母膏,装液量61.43 mL和初始pH5.63,L-茶氨酸产量达到17.22 mg/mL。粘盖牛肝菌产L-茶氨酸的液态发酵组合是30.12 g/L葡萄糖、6.77 g/L 酵母膏,装液量60.10 mL 和初始pH5.58,在此条件下L-茶氨酸的产量达到15.15 mg/L,研究结果为牛肝菌液态发酵产L-茶氨酸提供了一条新路径。
利用液态发酵技术生产牛肝菌代谢物质已经是获取生物活性成分的有效途径,通过改变培养基组成和发酵条件来获取不同的牛肝菌代谢物,进一步分析代谢物质的结构和功能,为保健食品和药品的研发提供了新的依据,也牛肝菌资源的开发与利用指出了新的研究方向。
3 牛肝菌液态发酵产物的诱导培养
在真菌液态发酵的过程中,胞外多糖、三萜类化合物、酚类化合物及生物碱等多种具有生物活性的代谢产物也随之产生,这些代谢产物生物活性强,毒副作用低,成为安全、有效、低毒且来源丰富的天然活性物质来源。近年来,为了促进真菌液态发酵过程中生物活性代谢成分的生成,常通过紫外光照射、添加茉莉酸甲酯、水杨酸、金属离子及真菌诱导子等物理、化学和生物外源诱导技术手段进行调控,进而诱导真菌液态发酵过程中代谢产物的生成,己成为真菌活性成分及其药物研究领域的新热点。
3.1 物理诱导培养
光照、紫外光照射、温度变化、机械损伤、超声、X 射线及电磁处理等是影响真菌液态发酵的物理诱导方式,其中紫外光照射会对真菌次生代谢产生重要影响,尤其会显著提高黄酮类化合物的产量[48-49]。紫外辐射作为一种危害性胁迫,可以激发植物体产生一系列的防御性反应,这些反应增加了大量黄酮类化合物及其它能够吸收紫外光的化合物,能够抵御照射到植物的大部分紫外光,减轻了植物被紫外光伤害的程度,同时也促进了大量类胡萝卜素的产生,保护了植物的光合体系[50-51]。
阳飞[52]首次采用原生质体紫外线-氯化锂(UVLiCl)复合诱变的方法选育出美味牛肝菌EPS 高产菌株,并对诱变株(UL-7)发酵工艺进行了研究,在相同液体培养基下EPS 产量可达到4.361 g/L,高出未经诱导的EPS 产量的37.8%。随着次生代谢活性成分研究的不断的发展,物理诱导也为促进牛肝菌代谢产物的产生开辟出一条新途径。
3.2 化学诱导培养
化学诱导剂如中药、中药提取物、油脂类物质及金属元素都是提高真菌代谢产物的重要手段,它能改变真菌代谢途径中关键酶的活性,通过快速、专一和选择性地诱导真菌特定基因的表达而促进细胞的合成和特定代谢产物的积累,从而达到促进真菌代谢产物产生的目的[53]。
王明慧等[54]研究培养基中水杨酸对褐环粘盖牛肝菌菌丝生长量的影响,发现水杨酸浓度在(4.76×10-10)~(4.76×10-6)g/mL 之间对菌丝产量有促进作用,当水杨酸浓度在2.38 × 10-10g/mL 时,菌丝生物量达到最大值420 g/L 左右,远高于没有经过化学诱导的菌丝产量91.86 g/L。苗廷廷[55]研究了砖红绒盖牛肝菌菌丝在含有大米和麸皮的培养基中的生长状况,发现在接种7~10 d 后,菌丝在含有大米的培养基上生长速度快,而在添加麸皮的培养基上,直到20~25 d 后才表现出了极好的生长趋势,且菌丝粗壮致密,菌丝在添加大米或者麸皮的培养基上的生长性状更好。
目前,将化学诱导剂添加到牛肝菌发酵培养基中来调控次生代谢产物的合成还未见报道,但在其他真菌方面有过报道,如在灵芝液体发酵液中加10 mmol/L Ca2+后,多糖含量比对照提高34%,三萜含量比对照提高64%,而在发酵液中加150 μmol/L水杨酸后,多糖含量比对照提高57.54%,三萜含量比对照提高37.19%,灵芝液体发酵12 d 时加10 mmol/L Ca2+和150 μmol/L 水杨酸,三萜含量比对照可以提高46.9%[56]。在桦褐孔菌液态培养第6 d 时添加50 mmol/L 茉莉酸甲酯(添加量为2 μL/mL)时,总三萜产量在发酵第9 d 时,最大达81.74 mg/100 mL,比对照的总三萜产量(60.76 mg/100 mL)提高了34.5%。而亚油酸、油酸和软脂酸也能促进桦褐孔菌在发酵过程中总三萜的积累[57]。由此说明,通过化学诱导促进牛肝菌三萜类化合物的合成是完全可行的,也为药物研究者指明了开发高效、低毒的抗癌药物新的研究方向。
3.3 生物诱导培养
生物诱导子是指微生物或者来源于微生物的成分,包括细菌、真菌(孢子和菌丝体)、酵母、微生物细胞壁成分、微生物蛋白质及脂类成分、微生物代谢产物、酵母提取物等[50],采用真菌诱导子诱导特定次生代谢产物已成为近年来的研究热点。真菌诱导子(fungal elicitors)是一类来源于真菌且能够诱导细胞生长和次生代谢产物合成的化学物质,包括有真菌菌体、菌液浓缩物、菌丝和菌液提取物、菌丝体高温处理后的可溶性成分、细胞壁降解成分、肽类和蛋白质等[58]。
段慧[59]分析了平菇汁不同处理方式对褐环粘盖牛肝菌菌丝生长量的影响,结果发现添加10%平菇汁的培养基在发酵15 d 时菌丝生长速度最快,菌丝干重比对照提高了158%。平菇汁75%乙醇萃取后得到的沉淀进行脱蛋白分离得到粗多糖,将不同浓度粗多糖溶液添加到培养基后发酵12 d,并在粗多糖溶液添加量为0.2 g/50 mL 时,菌丝干重高于对照达98%。赵磊[60]研究得出平菇汁乙醇透析液中的多肽含量与褐环粘盖牛肝菌菌丝生长量呈正相关,促进菌丝生长的真菌诱导子为透析液中的非脂溶性蛋白质或多肽。
在真菌液态培养基中加入外源诱导因子可以改善代谢途径中的基因表达与酶活,进而提高真菌代谢产物的生成量,具有较强的特异性[61]。目前,虽然对于诱导子的筛选及在真菌液态发酵中的应用方面有一定的研究,但对新型诱导子的研究特别是真菌诱导子筛选、诱导机理及调控机制方面,还存在很多的研究空白。因此,深入研究诱导子的诱导机理,筛选出高效诱导子加入到牛肝菌液态发酵培养基中,提高目标代谢产物及其生物合成产量,实现牛肝菌资源的高值化利用,具有重要的科学研究价值。
4 结论
采用真菌液态发酵方式培养菌丝及生物活性成分,不受自然资源环境限制,具生产周期短、便于精确控制等优点,能够进行大批量生产。另外,通过改变液态培养基基质组成、发酵培养条件、外源诱导等手段优化真菌次生代谢产物的生成,已成为食品及医药行业获取功效成分的新途径,具有重要的科学研究意义。牛肝菌资源来源丰富、营养价值和药用价值高,在液态发酵过程中利用外源诱导技术促使其产生丰富且生物活性好的代谢产物,不仅为牛肝菌科真菌液态发酵制备生物活性成分奠定一定的理论基础,也将成为牛肝菌科真菌资源高效利用的新方向。
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
文萃报·周五版(2021年30期)2021-09-05
科学技术创新(2021年19期)2021-07-16
食用菌(2021年3期)2021-06-03
浙江农业学报(2021年5期)2021-05-31
表面工程与再制造(2019年3期)2019-09-18
表面工程与再制造(2019年1期)2019-05-11
蔬菜(2018年8期)2018-08-14
资源再生(2017年3期)2017-06-01
中国食用菌(2017年2期)2017-01-14
