
青鱼和鲢鱼肌球蛋白热诱导凝胶特性的比较
2021-06-16 10:26毕保良王晓雯
食品工业科技 2021年3期
高 宇,毕保良,贾 丹,胡 青,王晓雯
(云南农业大学动物科学技术学院,云南省教育厅高原渔业资源保护 与可持续利用重点实验室,云南昆明 650201)
青鱼(Mylopharyngodonpiceus)和鲢鱼(Hypophthalmichthysmolitrix),作为我国传统的“四大家鱼”,养殖历史十分悠久,其产量一直位居前列。到2018年我国青鱼和鲢鱼年产量已分别达69.1和385.9万吨[1]。鲢鱼产量虽大,但其凝胶特性较差。目前,关于鲢鱼肌球蛋白热诱导凝胶的形成机制及其高值化利用已有较多报道[2-6]。
青鱼具有生产快、采肉率高、凝胶特性好等特点,在我国常被用于加工成鱼糜制品。目前国内学者已对青鱼等大宗淡水鱼的原料基本特性和凝胶特性进行了比较[7]、对鱼皮胶原特性[8]以及贮藏中品质变化等开展了研究[9],也有学者对不同养殖模式下青鱼肌肉的质构特性[10]和营养品质[11]进行了比较分析。低温贮藏下,青鱼肉中蛋白和组织结构的变化会影响鱼肉中水分的分布状态,从而对品质造成影响[12]。张屹环等[13]比较了青鱼和其他大宗淡水鱼的凝胶特性差异,发现热处理鱼糜凝胶中,青鱼糜凝胶的凝胶强度仅低于鲤鱼,而高于鲂鱼、鳙鱼、鲢鱼、鲫鱼和草鱼。课题组前期研究得出MTGase催化谷氨酰胺残基的γ-羧酰胺基和伯胺之间发生的交联反应和二硫键共同导致了鱼糜凝胶网络的形成,蛋白质变性伸展,二硫键和疏水相互作用的增加,青鱼蛋白凝胶的网络结构进一步增强[14]。而肌球蛋白是鱼糜凝胶形成最主要的蛋白质[15],但目前关于青鱼肌球蛋白凝胶形成机制方面的研究几乎空白。
鱼糜的凝胶特性不仅受内源蛋白酶的降解和交联酶类交联作用的影响[16],而且还受构象和蛋白质热特性的影响。构象的变化能引起蛋白质的变性,从而引起蛋白质之间的聚集,适当的聚集是凝胶网络结构形成的提前条件[17]。蛋白质变性后,其构象、化学作用力、溶解性能、蛋白质粒径大小均会发生变化,因此用二级结构、总游离巯基、疏水相互作用、粒径分布可以表征蛋白质的变性和聚集[18]。可用流变仪研究其凝胶形成过程,动态流变曲线中的弹性模量G′、粘性模量G″和损耗角δ,这些参数和凝胶的结构紧密相关[19]。因此本研究以青鱼和鲢鱼肌球蛋白为研究对象,探讨肌球蛋白在热诱导凝胶形成过程中的构象变化,变性和聚集特性及凝胶形成的差异,旨在揭示青鱼糜凝胶形成机制,为开发高品质鱼糜凝胶制品提供一定的理论研究基础。
1 材料与方法
1.1 材料与仪器
鲜活青鱼和鲢鱼 购于华中农业大学菜市场,总重1500 g/条左右;三磷酸腺苷二钠(ATP-Na2)、5,5-二硫代双(2-硝基苯甲酸)(DTNB)、叠氮钠、乙二醇-双-(2-氨基乙基)四乙酸(EGTA) Amresco公司;1-苯胺萘-8-萘磺酸盐(ANS)、Tris 美国Sigma公司;β-巯基乙醇 上海化学试剂总厂;氯化钠、氯化镁等试剂 均为分析纯,国药集团化学试剂有限公司。
Discovery H-2动态流变仪 美国TA公司;J-8100圆二色谱 日本JASCO公司;ZEN3600马尔文激光粒度仪 Zetasizer Nano ZS马尔文有限公司;F-4600荧光光谱仪 日本Hitachi公司。
1.2 实验方法
1.2.1 青鱼、鲢鱼前处理 将鲜活青鱼和鲢鱼20 min内迅速拿回实验室,宰杀取肉在30 min内完成。
1.2.2 肌球蛋白的提取 参考Guo等[20]的方法。取尸僵前的青鱼、鲢鱼背脊肉分别用食品调理机破碎,向破碎后的肌肉中加入10倍体积的溶液A(pH7.5,含0.1 mol/L KCl和0.02% NaN3的20 mmol/L Tris-HCl缓冲液),并用高速分散均质机于23000 r/min的转速下均质1 min,均质液于5 ℃下放置15 min,3836×g离心5 min,沉淀于5倍体积溶液B(pH6.8,含0.45 mol/L KCl、5 mmol/Lβ-巯基乙醇、0.2 mol/L醋酸镁(Mg(CH3COO)2)、1 mmol/L EGTA的20 mmol/L Tris-HCl缓冲液)中悬浮,同时加入ATP-Na2使肌球蛋白与肌动蛋白解离,鱼肉肌球蛋白的提取需加ATP-Na2至终浓度为5 mmol/L。于5 ℃下放置1 h后在17696×g离心10 min,上清液用5倍体积1 mmol/L KHCO3稀释,于5 ℃下放置15 min,然后于22095×g离心10 min;沉淀重新用2.5倍体积溶液C(pH7.5,含0.5 mol/L KCl和5 mmol/Lβ-巯基乙醇的20 mmol/L Tris-HCl缓冲液)悬浮,于5 ℃下放置10 min,再用2.5倍体积的1 mmol/L KHCO3稀释并加MgCl2至终浓度为10 mmol/L,于5 ℃下放置过夜,然后23708×g离心10 min;沉淀物用1倍体积溶液 D(pH7.0,含0.6 mol/L NaCl的20 mmol/L Tris-HCl缓冲液)溶解,即为肌球蛋白溶液,于4 ℃保存,2 d内用完。
1.2.3 肌球蛋白的测定
1.2.3.1 圆二色谱的测定 将0.05 mg/mL的肌球蛋白溶液放入10 mm光程的石英样品池中,在远紫外区(198~250 nm)对肌球蛋白溶液进行扫描,实验参数:Data pitch:0.1 nm CD scale:200 mdeg/1.0 dOD;FL scale:200 mdeg/1.0 dOD;FL scale;Bandwidth:1 nm;Scanning speed:50 nm/min。平均残基分子量为110 g/mol。研究加热过程中肌球蛋白二级结构的变化,采用的升温速率为2 ℃/min(10~90 ℃),各二级结构(α-螺旋、β-折叠、β-转角和无规卷曲)所占比例用圆二色光谱仪自带的程序进行分析。
1.2.3.2 表面疏水性的测定 将提取出的肌球蛋白用溶液D稀释成0.001、0.005、0.01、0.04和0.06 mg/mL系列浓度,加入20 μL 8 mmol/L ANS的0.1 mol/L磷酸盐缓冲溶液(pH7.0),于20、30、40、50、60、70、80、90 ℃加热30 min后,在激发波长364 nm下,用荧光光谱仪测定其在发射波长534 nm下的荧光强度,表面疏水性(S0-ANS)以荧光值-蛋白质浓度曲线的斜率来表示[17]。
1.2.3.3 二硫键含量的测定 游离巯基(SHF),总巯基(SHT)和二硫键(S-S)含量的测定参考Jia等[14]的方法。取1 mg/mL的肌球蛋白溶液于20、30、40、50、60、70、80、90 ℃加热30 min后,即得到热诱导肌球蛋白溶液,冷却后涡旋混匀备用。测定SHF含量时,移取200 μL Ellman试剂(5,5′-dithio-bis-2-nitrobenzoic acid(DTNB)溶解于Tris-甘氨酸缓冲液,4 mg/mL),添加到4 mL的肌球蛋白溶液中,室温下反应1 h,于412 nm测定其吸光值。测定SHT含量时,1 mL肌球蛋白溶液中加入4 mL 20 mmol/L Tris-Gly缓冲液(pH8.0,含15 mg/mLβ-巯基乙醇,8 mol/L尿素和5 mol/L盐酸胍)于40 ℃ 1 h,然后用12%的TCA沉淀蛋白1 h,4000×g离心10 min,收集沉淀用12%的TCA漂洗3次,沉淀用10 mL的Tris-Gly缓冲液,含8 mol/L尿素溶解。取200 μL的Ellman试剂添加到4 mL的蛋白溶液中,然后于412 nm测吸光值,SHF、SHT和S-S含量用式(1)和(2)表示:
SH(μmol·g-1protein)=73.53×A412/C
式(1)
S-S(μmol·g-1protein)=(SHT-SHF)/2
式(2)
式中,C-蛋白浓度,mg/mL;A412-412 nm处的吸光值;73.53=106/13600,13600 mol/L·cm即是摩尔消光系数。
1.2.3.4 粒径的测定 将提取的肌球蛋白用溶液D调整到0.5 mg/mL,NaCl溶液采用高纯水配制,分别于20、30、40、50、60、70、80、90 ℃加热30 min。待冷却后涡旋混匀后采用ZEN3600马尔文激光粒度仪测定其粒径。
1.2.3.5 动态流变的测定 参考Gao等[21]的方法并略作修改,采用Discovery H-2动态流变仪的平行板(直径为4 cm,载物台与平板间的距离为1 mm)进行测定。将浓度为10 mg/mL的肌球蛋白溶液,用液体石蜡的密封盖封住样品以避免水分蒸发,按照2 ℃/min从10 ℃升温到90 ℃,采用1 Hz的振动频率,剪切应力为1 Pa。
1.3 数据处理
所有数据采用SAS 8.0进行分析,3个或3个以上样本的比较采用单因素方差分析(One-way ANOVA)Duncan检验,2个样本的比较采用独立样本t检验(Independent-Samples T Test)。
2 结果与分析
2.1 加热过程中肌球蛋白二级结构的变化
青鱼和鲢鱼肌球蛋白在加热过程中二级结构的变化如图1所示。青鱼肌球蛋白加热到20 ℃,二级结构没有发生明显的变化,α-螺旋含量随着温度的升高呈下降的趋势,到温度为80 ℃时其含量又有稍微的上升趋势。α-螺旋含量由加热前的69.2%(10 ℃)降至21.1%(90 ℃),即有48.1% 的α-螺旋解开,而β-折叠、β-转角和无规卷曲的比例分别从10 ℃的0、5.2%和25.5%变化为90 ℃时18.7%、30.7%和29.6%。β-折叠和β-转角的变化量为18.7%和25.5%,而无规卷曲的变化量仅为4.1%,说明加热使得青鱼肌球蛋白的α-螺旋大部分转变为β-折叠和β-转角。鲢肌球蛋白的α-螺旋含量也随温度的升高而降低,由加热前的83.4%(10 ℃时)降至30.8%(90 ℃时),即肌球蛋白分子中约有52.6%的α-螺旋解开,β-折叠由0(10 ℃)变化为2.2%(90 ℃),β-转角由0(10 ℃)变化为21.8%(90 ℃),从50 ℃开始鲢肌球蛋白才开始有β-折叠,而β-转角的形成则是从60 ℃开始的,β-折叠和β-转角的变化量为2.2%和21.8%。而无规卷曲的含量随着温度的升高呈增大趋势,从17.7%变化到45.3%,变化量为27.6%。这说明青鱼肌球蛋白在加热过程中β-折叠增大程度明显高于鲢鱼肌球蛋白。Yongsawatdigul等[18]发现当马鲅鱼肌动球蛋白从10 ℃升温到80 ℃时,α-螺旋含量从66%下降到24.2%,且在高于30 ℃时α-螺旋含量开始下降,这和本研究趋势基本一致;Liu等[22]研究证明加热过程中α-螺旋的解开和β-折叠的形成都有利于肌球蛋白的胶凝作用。

图1 加热温度对肌球蛋白二级结构相对含量的影响Fig.1 Effects of heating temperatures on the relative content of secondary structures of myosin注:a:青鱼;b:鲢鱼。
2.2 加热对肌球蛋白疏水性的影响
表面疏水性是蛋白表面特性的重要指标之一,S0-ANS的增加源自于蛋白的解折叠从而导致疏水性基团的暴露。暴露的疏水氨基酸残基可以促进疏水相互作用的形成而促进蛋白的聚集[23-24]。由表1可知,随着加热温度升高,青鱼和鲢鱼肌球蛋白的S0-ANS显示先增加后下降的趋势。青鱼肌球蛋白的S0-ANS在70 ℃达到最大值,而鲢鱼肌球蛋白的S0-ANS在60 ℃时达到最大,这说明随着温度的升高,肌球蛋白α-螺旋的解折叠作用使得疏水氨基酸残基暴露,故S0-ANS增大。但随着温度的升高,S0-ANS的降低可能是疏水性相互作用导致的蛋白聚集体的形成,从而使疏水基团包裹在聚集体内的结果[12]。整体来看,青鱼肌球蛋白的S0-ANS显著高于鲢鱼(P<0.05)。这是因为青鱼肌球蛋白在热诱导凝胶形成过程中的展开程度高于鲢鱼,导致青鱼肌球蛋白疏水性侧链暴露程度高,故在升温过程中青鱼肌球蛋白疏水相互作用高于鲢鱼。

表1 加热温度对肌球蛋白表面疏水性的影响Table 1 Surface hydrophobicity of myosin fish under different heating temperatures
2.3 加热对肌球蛋白二硫键的影响
由表2可知,青鱼和鲢鱼肌球蛋白的二硫键含量随着温度的升高而升高,青鱼和鲢鱼肌球蛋白二硫键含量在60~90 ℃时均无显著性差异(P>0.05)。鱼糜肌原纤维蛋白含有大量的半胱氨酸的巯基,可被氧化形成分子间或分子内的二硫键,从而增强鱼糜凝胶的网络结构和凝胶强度[25]。这说明在升温过程中二者肌球蛋白部分游离巯基均被氧化成二硫键。且青鱼肌球蛋白在热诱导凝胶形成过程中的二硫键含量与鲢鱼无显著性差异(P>0.05)。

表2 加热温度对肌球蛋白二硫键的影响Table 2 S-S bonds of myosin fish under different heating temperatures
2.4 肌球蛋白溶液的平均粒径
实验进行粒径分布测试时,每个样品所得的PDI值均在0.3~0.7范围内,说明蛋白溶液体系较均一。青鱼和鲢鱼肌球蛋白溶液在不同加热温度下的平均粒径(dz)如表3,由表3可以看出青鱼和鲢鱼肌球蛋白的dz随着温度的升高而增加,青鱼肌球蛋白的dz在90 ℃达到最高,而鲢鱼肌球蛋白的dz在80 ℃达到最高。且当加热温度超过40 ℃后,青鱼肌球蛋白的平均粒径显著大于鲢鱼的(P<0.05)。当在50和90 ℃加热30 min后,青鱼肌球蛋白的平均粒径分别为1303.67和4880.00 nm,而鲢鱼的分别为596.46和2013.67 nm。青鱼肌球蛋白的dz约为鲢鱼肌球蛋白的2倍。说明在温度高于40 ℃时,青鱼肌球蛋白比鲢鱼形成更大的聚集体,这可能是因为在加热过程中,青鱼肌球蛋白较鲢变性程度更高,结构更为伸展,较强的疏水相互作用使得青鱼肌球蛋白更容易发生聚集。
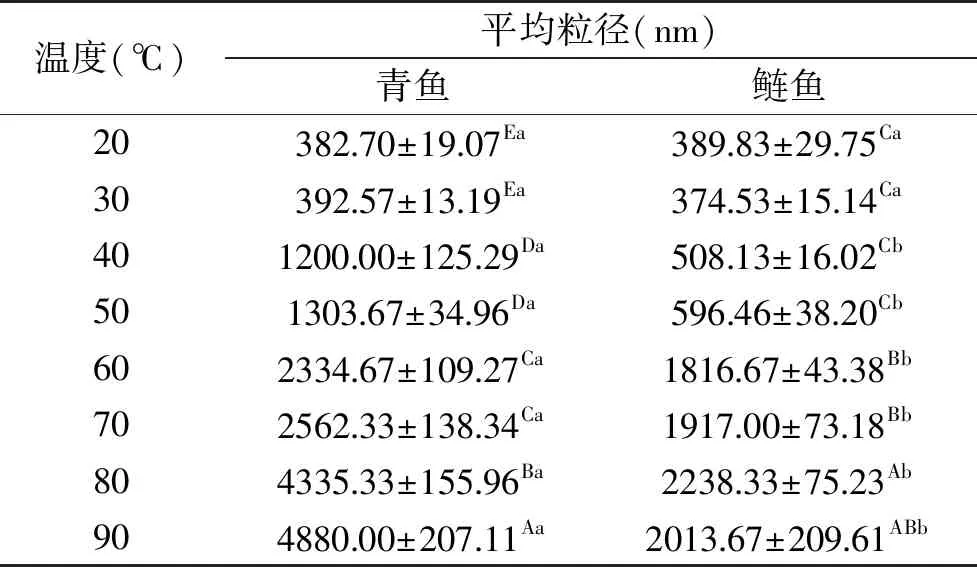
表3 加热温度对溶液中肌球蛋白平均粒径(dz)的影响Table 3 Average size(dz)of myosin solutions under different heating temperatures
2.5 加热过程中肌球蛋白的流变学特性
G′和δ可以用来表征蛋白在加热过程中的动态流变的变化,图2显示青鱼和鲢鱼肌球蛋白热诱导凝聚体形成过程中的流变学特性,由图2可知,在10~37 ℃的范围内,青鱼肌球蛋白的G′略呈下降的趋势,而鲢鱼肌球蛋白在此阶段的保持G′基本不变的趋势,而两者肌球蛋白的δ呈下降的趋势。在37~48 ℃的范围内,两种肌球蛋白的G′均呈现上升,δ下降的趋势,这是因为在低温阶段,肌球蛋白的流变学特性的变化主要是来自于蛋白质分子中热稳定性较弱的区域的结构的改变。30~40 ℃时肌球蛋白轻链会发生解链[26],蛋白质之间发生交联,形成不稳定的聚集体,使体系的黏度上升,从而使得G′上升,δ下降。G′达到第一个峰值及达到峰值的温度随着鱼种、加热速度、蛋白浓度等的不同而不同。淡水鱼G′达到峰值的温度存在差异,青鱼和鲢鱼肌球蛋白G′达到峰值的温度分别为48.9和46.6 ℃。且两者的峰值G′分别为278.48和83.95 Pa。进一步加热,G′迅速下降,于55.02~58.80 ℃达到最小值,之后稳步上升。G′的降低可能是由于肌动球蛋白解离和肌球蛋白解折叠引起的[27],且高温破坏了以氢键为主要作用力的低温凝胶网络结构,从而使得G′降低[28]。在48.4~58.80 ℃,δ的值是呈现先上升后下降的趋势。继续升高温度,肌球蛋白G′的上升和δ下降,有可能是高温使得二硫键和疏水相互作用等作用力的形成,从而促进凝胶网络结构的形成[23,26]。青鱼肌球蛋白的最终G′和δ分别为65.11 Pa和2.80 °,而鲢鱼肌球蛋白的为37.67 Pa和4.23°。青鱼肌球蛋白的热诱导凝聚体形成能力高于鲢鱼,其主要原因是青鱼肌球蛋白分子的展开程度、疏水相互作用高于鲢鱼。

图2 肌球蛋白热诱导凝胶形成过程中的 动态流变学性能的变化Fig.2 Changes in dynamic rheological properties of myosin during thermal gelation
2.6 肌球蛋白二级结构在热胶凝过程中的作用
为了研究肌球蛋白的二级结构在热凝胶形成过程中的作用,分析肌球蛋白在加热过程中G′的变化与α-螺旋、β-折叠和β-转角含量之间的关系。根据图2中G′的变化可将肌球蛋白凝胶形成过程分成3个阶段:10~45 ℃、45~60 ℃和60~80 ℃,对各阶段G′与肌球蛋白α-螺旋、β-折叠和β-转角含量分别用线性模型(1)、模型(2)和模型(3)进行拟合。模型如下:回归结果见下表4。

表4 肌球蛋白的弹性模量G′与α-螺旋、β-折叠和β-转角比例的相关性分析Table 4 Regression model for G′ of surimi on α-helical,β-sheet and β-turn contents
G′=k1A+b1
式(1)
G′=k2B+b2
式(2)
G′=k3C+b3
式(3)
由表4可知,在10~45 ℃温度范围内,青鱼肌球蛋白的G′与α-螺旋含量呈显著负相关(P<0.05),而鲢鱼肌球蛋白的G′与α-螺旋含量呈极显著负相关(P<0.01),青鱼肌球蛋白的G′与β-折叠呈极显著正相关(P<0.01),而鲢鱼肌球蛋白与β-折叠无相关性。在45~60 ℃温度范围内,青鱼肌球蛋白的G′与β-转角含量无相关性,而鲢鱼肌球蛋白的G′与β-转角含量呈显著负相关(P<0.05)。而在60~80 ℃温度范围内,青鱼和鲢鱼肌球蛋白的G′均与α-螺旋含量呈显著负相关(P<0.05),青鱼肌球蛋白的G′与β-转角呈显著正相关(P<0.05),鲢鱼肌球蛋白的G′与β-转角呈极显著正相关(P<0.01)。综合来说,青鱼和鲢鱼肌球蛋白分子α-螺旋向β-折叠或β-转角转变有利于肌球蛋白的热凝胶作用。
3 结论
青鱼和鲢鱼肌球蛋白在热诱导凝胶形成过程中,两者二级结构存在一定的差异,青鱼肌球蛋白自身的α-螺旋含量较低,β-转角含量较高,青鱼蛋白结构更为伸展,在热诱导凝胶形成过程中,青鱼肌球蛋白的展开程度高于鲢鱼,青鱼肌球蛋白较强的疏水相互作用使其聚集速度较快,聚集体的dz大,较高的变性和聚集程度使得蛋白凝胶形成能力强于鲢鱼,且肌球蛋白分子α-螺旋向β-折叠或β-转角转变有利于肌球蛋白的热凝胶作用。但二者热诱导蛋白凝胶的嫩度、咀嚼性、保水性、颜色等最终影响产品的品质和口感等特性仍需进一步研究。
猜你喜欢
石油化工腐蚀与防护(2022年1期)2023-01-06
食品科学技术学报(2022年5期)2022-10-11
食品科学(2022年7期)2022-05-13
食品科学(2022年8期)2022-05-09
现代食品科技(2022年4期)2022-04-28
化工设计通讯(2020年11期)2020-11-17
食品与机械(2020年5期)2020-07-03
文萃报·周二版(2019年22期)2019-09-10
分析化学(2019年4期)2019-05-13
清明(2016年6期)2016-12-17
