
武汉地区2017年-2019年蚊类群落组成及其多样性分析
2021-06-10 03:22:24毛伟平李华刚陈晓敏田俊华
华中师范大学学报(自然科学版) 2021年3期
柳 静,毛伟平,丁 浩,李华刚,陈 龙,陈晓敏,田俊华*
(1.武汉市疾病预防控制中心,武汉 430015;2.武汉市东西湖区疾病预防控制中心,武汉 430040;3.武汉市青山区疾病预防控制中心,武汉 430080;4.武汉市黄陂区疾病预防控制中心,武汉 430345;5.武汉市江夏区疾病预防控制中心,武汉 430200)
蚊类属于昆虫纲、双翅目(Diptera)、蚊科(Culicidae),是最重要的医学昆虫类群.蚊类不仅叮咬吸血,严重骚扰人和动物,还可以传播多种蚊媒疾病.现已知在我国传播疟疾的媒介主要以嗜人按蚊(Anophelesanthropophagus)、中华按蚊(An.sinensis)、微小按蚊(An.minimus)和大劣按蚊(An.dirus)四种为主[1].传播班氏丝虫病的主要蚊种是致倦库蚊(Culexquinquefasciatus)/淡色库蚊(Cx.pipienspallens),而嗜人按蚊和中华按蚊则是马来丝虫病的主要传播媒介[2].三带喙库蚊(Cx.tritaeniorhynchus)是乙脑流行地区的主要媒介,白纹伊蚊(Aedesalbopictus)和埃及伊蚊(Ae.aegypti)则主要传播登革热[3].蚊类分布很广,种类很多.陆宝麟等人编撰了《中国蚊科志(上、下卷)》,记载中国蚊种18属361种(亚种)[4-5].标志着我国蚊类区系分类研究已达到成熟阶段[6].随着分子生物学、信息系统的发展,宏观与微观相结合的技术运用至蚊类多样性的研究中,越来越多的隐存种、亚种、新种被发现,目前中国已知的蚊类达2亚科46属、418种[7].对湖北地区蚊虫分布、地理区划研究多集中在20世纪80、90年代[8-9],且研究结果多是前人资料的整理和现场调查相结合,缺乏实验室技术的进一步考证,存在错漏等现象.近年来武汉地区蚊类研究多集中于蚊虫孳生地调查[10]、城市灭蚊经验探索[11]、蚊虫抗药性研究[12-13]、蚊虫种群密度监测[14-15]、种类及季节消长等.但蚊类作为重要的医学昆虫媒介在群落生态学方面研究较少,多样性指标在一定程度上可以反映群落的复杂程度和稳定程度[16-17],对武汉地区蚊类群落组成和多样性进行研究,可以为长江流域蚊类群落特征和多样性分布规律,物种多样性与疾病的关系,以及当地蚊类的防制、蚊传疾病的控制提供更多的理论依据.
1 材料与方法
1.1 调查方法 2017年-2019年在武汉共设20个调查点(图1).用诱蚊灯(“功夫小帅”诱蚊灯由武汉吉星环保科技有限责任公司生产)进行蚊虫采集.其中青山区和东西湖区分别选取2处居民区、2处公园、2处医院;黄陂区和江夏区分别选取2处农户、2处猪圈.每处选择远离干扰光源和避风的场所为挂灯点,诱蚊灯光源离地1.5 m,日落前1 h开启诱蚊灯,直至次日日出后1 h,密闭收集器后,再关闭电源,将集蚊袋取出.将捕获的蚊虫用冰箱冷冻处死后,记录种类及数量.鉴定方法主要参照陆宝麟等主编的《中国动物志双翅目:蚊科(上、下)》[4-5].该调查每月上旬、下旬各开展1次,相邻两次调查间隔不少于10 d,遇风(5级以上)雨顺延.

图1 武汉地区调查点位图Fig.1 Sampling sites of Wuhan area
1.2 数据分析 多样性分析分别采用Simpson指数D、Shannon-Wiener多样性指数H',Pielou的均匀度指数J',相似性分析使用Jaccard(1901)相似性指数Cj进行.其计算公式如下:
J'=H'/lnS,
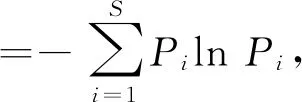
Cj=c/a+b-c,
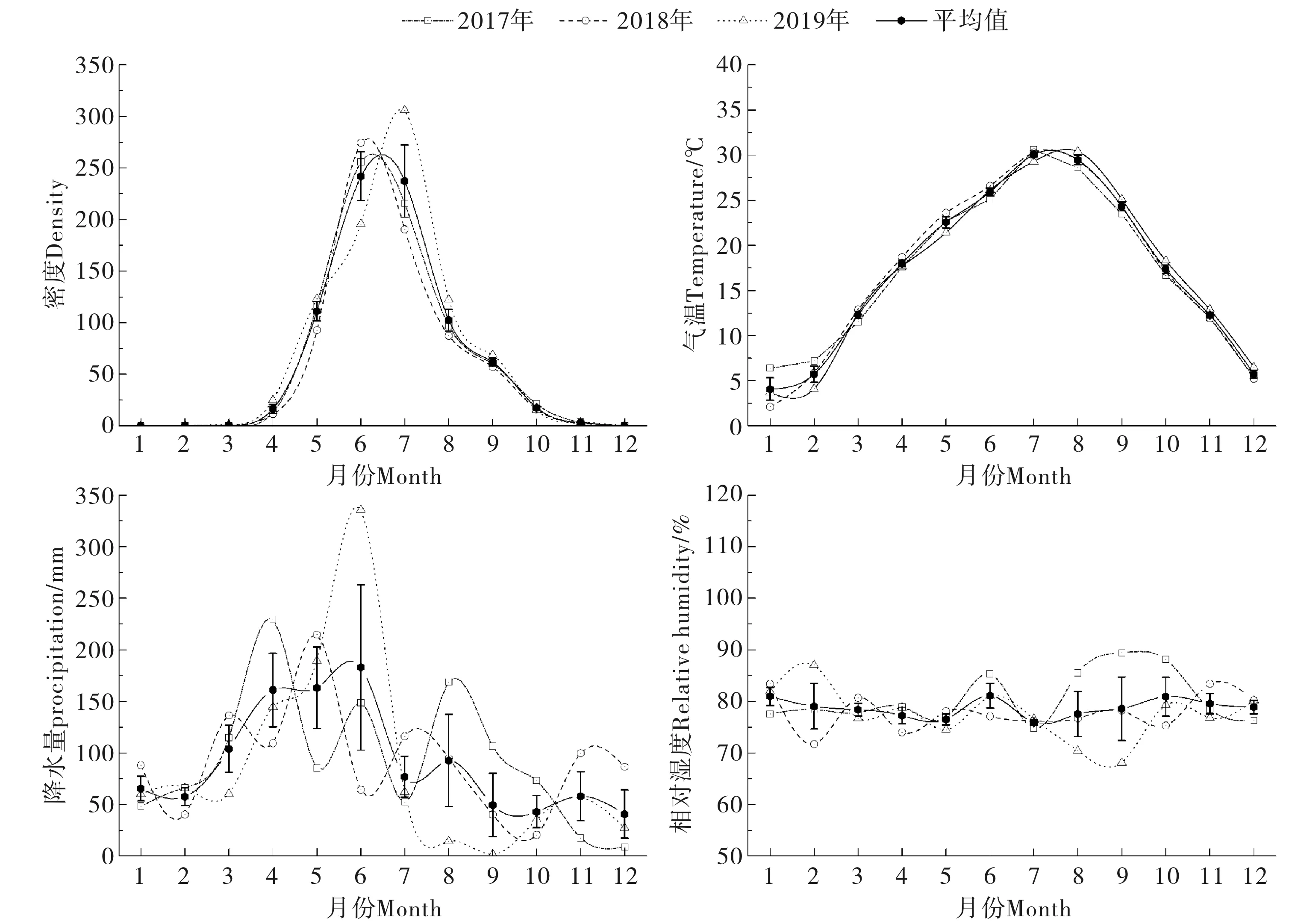
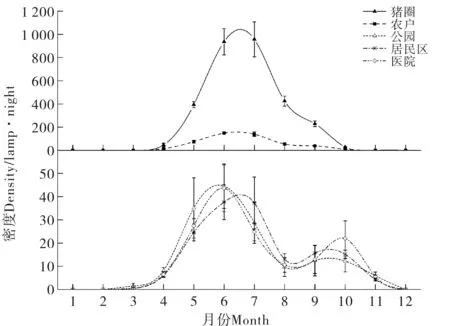
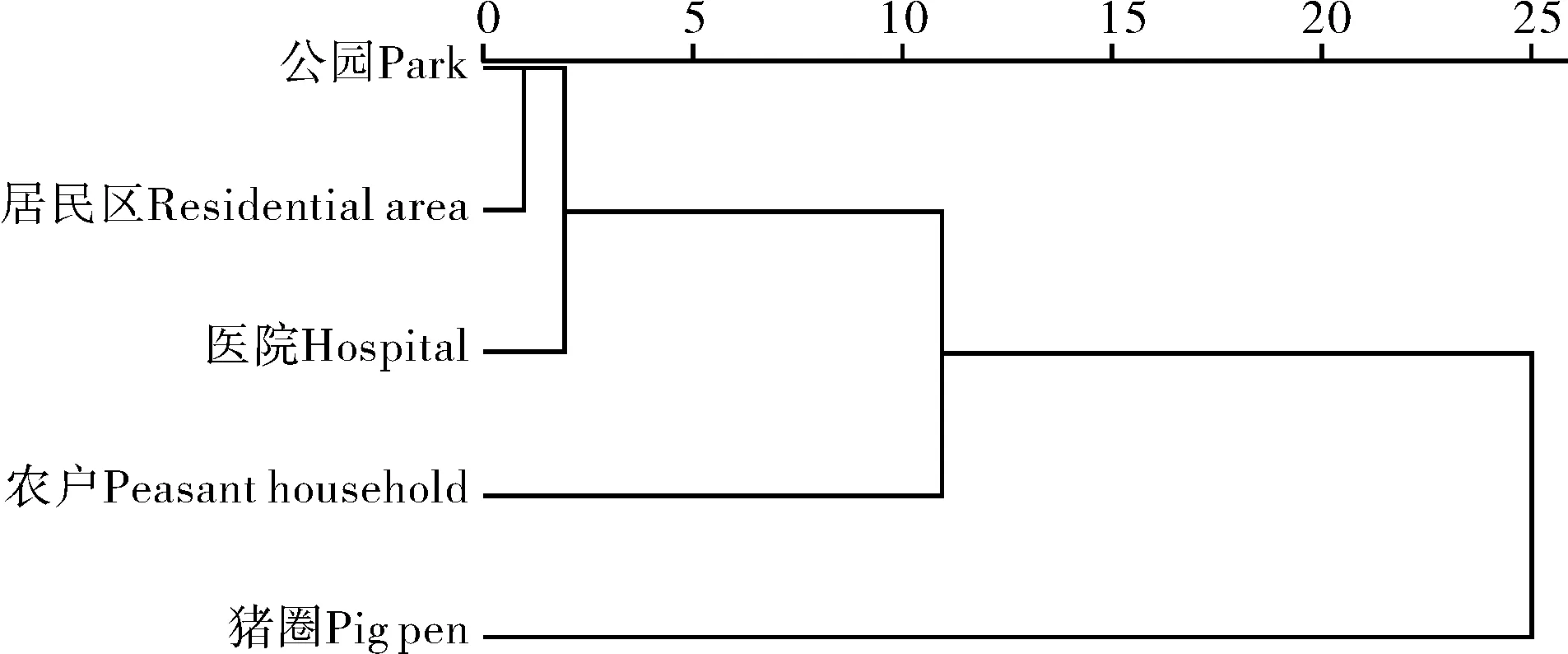
式中,S为物种数,Pi为物种i的个体数占群落总个体数的比例(即物种i的相对多度);ni为物种i的个体数,N为群落物种个体总数.根据Jaccard指数的原理,当0 2017年-2019年气温、降水量、相对湿度等气象数据来源于国家气象科学数据中心(http://data.cma.cn).使用Excel 2013、SPSS 21.0进行数据汇总、方差分析、相关性分析和聚类分析.点位图的绘制使用Adobe Illustrator 2018,其他图形绘制使用Origin 2016. 2017年-2019年,共布放诱蚊灯1 440个,捕获蚊类95 072只.经鉴定隶属1科5属8种,分别为三带喙库蚊(Cx.tritaeniorhynchus)、致倦库蚊(Cx.quinquefasciatus)、中华按蚊(An.sinensis)、骚扰阿蚊(Armigeressubalbatus)、刺扰伊蚊(Ae.vexans)、白纹伊蚊(Ae.albopictus)、常型曼蚊(Mansoniauniformis)、二带喙库蚊(Cx.bitaeniorhynchus).其中三带喙库蚊和致倦库蚊为优势种,平均相对多度分别为62.53%和20.46%;中华按蚊、骚扰阿蚊和刺扰伊蚊为常见种,平均相对多度为9.26%、5.84%、1.14%(见表1). 表1 2017年-2019年武汉市蚊虫种类和数量Tab.1 Species and numbers of mosquitoes obtained in Wuhan from 2017 to 2019 2017年-2019年,武汉地区蚊类活动一般从3月份开始,一直持续到11月份,蚊虫密度季节消长趋势呈单峰型,2017年、2018年密度高峰出现在6月份,2019年密度高峰略有后移,出现在7月份,最高值为每灯每夜305.98只.根据双因素方差分析,年度间蚊密度差异不显著(F=0.681,p>0.05),月份间蚊密度差异极显著(F=47.133,p<0.01)(图2). 图2 武汉地区2017年-2019年蚊虫密度季节消长及气候因子变化Fig.2 Seasonal fluctuation of mosquito density and climatic factors in Wuhan from 2017 to 2019 2017年-2019年平均气温、降水量、相对湿度等气象因子与蚊虫密度进行Spearman相关性分析发现,蚊虫密度与平均气温相关性最强(ρs=0.927,p<0.01),与降水量和相对湿度相关性不大(表2). 表2 蚊虫密度与平均气温、降水量、相对湿度的Spearman相关性Tab.2 Spearman correlation between mosquito density and average temperature,precipitation,and relative humidity 不同生境蚊虫的平均密度水平依次为猪圈>农户>公园>居民区>医院.其中城市生境如公园、居民区、医院蚊虫密度季节消长呈双峰型,其中第一个高峰出现在6月-7月,第二个高峰出现在9月-10月;而农村生境如猪圈、农户蚊虫密度季节则呈现单峰型季节消长,密度高峰出现在6月-7月.根据双因素方差分析,5种生境间蚊密度差异极显著(F=5.654,p<0.01),月份间蚊密度差异不显著(F=1.771,p<0.01)(图3). 图3 2017年-2019年不同生境蚊虫密度季节消长Fig.3 Seasonal fluctuation of mosquito density in different habitats from 2017 to 2019 根据2017年-2019年5种生境蚊类物种多样性指标的分析显示,猪圈物种丰富度最高为8,医院物种丰富度最低为3.比较5种生境蚊类Simpson优势度指数和Shannon-Wiener多样性指数,2017年、2018年农户多样性指数最高、猪圈次之、医院最低.2019年农户多样性指数最高、猪圈次之、公园最低.表明农户和猪圈蚊类多样性丰富,稳定程度高;公园、居民区、医院蚊类多样性水平较低,蚊类多样性的稳定程度较低. 比较2017年-2019年5种生境蚊类均匀度指数,其规律与多样性指数相似,农户最高,猪圈次之,其它三种生境均匀度指数相似,年度间差异不明显. 公园、居民区、农户、医院、猪圈5种生境间蚊类相似性指数年际间一致.其中公园、医院与居民区,猪圈与农户相似性指数范围在0.75~1.00,为极相似;其他生境之间相似性范围在0.25~0.50,为中等不相似(表3). 表3 武汉地区5种生境蚊类多样性指标(2017年-2019年)Tab.3 Species and numbers of mosquitoes obtained in Wuhan from 2017 to 2019 以各生境捕获的蚊虫种类与数量数据进行标准化处理后,根据欧氏距离原理,利用SPSS 21.0进行组间平均聚类法聚类分析.如图4所示,公园与居民区的蚊虫类群首先聚为一类,然后再与医院蚊虫类群聚为一类,三个蚊虫类群再与农户蚊虫类群聚为一簇,这4个调查点的蚊虫类群最后再与猪圈蚊虫类群合并. 图4 武汉地区五种生境蚊虫群系统聚类分析Fig.4 The hierarchical cluster dendrogram of mosquito species based on the Euclidean distance in Wuhan 蚊类多样性包括蚊类群落在时间生态位和空间生态位上蚊种丰富度、优势度、多样性及不同生境群落相似性等,反映蚊类在时空上与环境的相互关系.通过不同时期和不同生境蚊类群落生物多样性的调查和比较,可进一步了解蚊类在年际、季际、月际或昼夜不同时段的活动和消长规律[6].监测数据表明:武汉蚊类3月份开始活动,随着气温升高蚊虫密度逐月增加,一般6月-7月份达到密度高峰,可能与春末夏初雨量充沛、温度适宜有关,温度过高过低均不适应蚊类滋生繁衍;随后直至11月蚊虫密度逐渐降为0.12月到次年2月蚊虫一般以卵、幼虫或少数雌虫越冬或休眠[21].这与湖北地区历年监测结果保持一致[22-23].这提示了从3月份随着气温回升,在高峰期来临之前,应及时清理积水、疏通暗渠、管理好各类水体工作,培养居民良好的卫生习惯和防护意识等蚊媒综合防控措施. 比较五种生境蚊虫密度季节消长可以发现,城市生境如公园、居民区、医院蚊虫密度季节消长呈双峰型,9月-10月会出现第二个小高峰;而农村生境如猪圈、农户蚊虫密度季节则呈现单峰型季节消长.这可能与优势种、温度、降雨量、孳生场所和栖息场所有关.城市中以致倦库蚊为优势种,其相对多度在90%以上,它们一般孳生在污水里,如中小型容器、雨水井、下水沟、车库电梯井、污水明渠等生境;6月和9月武汉地区温湿度和降雨量更适合该蚊种的繁衍;而8月左右正值酷暑,武汉地区气温高,积水蒸发快,孳生地相对减少,同时高温也不利于幼虫发育,还会导致大量成蚊死亡,从而使成蚊密度急剧下降.而农村地区成蚊密度呈单峰型的原因可能是:1) 优势蚊种野栖型的三带喙库蚊相对多度极高,三年平均为70.87%;2) 灌木和草本植物丰富,为蚊类提供了丰富的栖息场所;3) 野栖型蚊类的主要孳生地稻田,因9月水稻进入成熟期而逐渐干涸. 群落中物种组成及其多样性是群落的重要特征,可以用来判断群落和生态系统的稳定性[24].在本研究中,农户生境的多样性指数最高,群落稳定性较强.可能由于农户和牲畜棚周边破坏程度较低,环境较天然,既多见稻田、沟渠、池塘等水体,同时政府干预农村灭蚊较少.另外牲畜为蚊类提供了充足的血源,这些都为更多的蚊类孳生提供了有利条件.而以公园、居民区、医院三种生境为代表的城市环境下,人类大规模生产、生活活动的长期干扰和选择下,破坏了生境的天然形态,导致蚊类孳生场所的异质性降低,蚊类群落稳定性、丰富度、多样性指数相应降低[25].同时也反应了城市生态系统下蚊类的多样性容易受到外界环境或者灭蚊干预措施影响[26]. 相似性分析采用了Jaccard指数原理,与不同群落蚊类的物种丰富度和蚊虫数量相关,而聚类分析则是基于欧式距离进行分析.结果表明,公园、居民区和医院蚊虫群落极相似,农户和猪圈蚊虫群落极相似.但农户和猪圈的欧式距离较远.可能由于猪圈中猪是一种很好的血源性引诱剂,对蚊类引诱效果极好,特别是三带喙库蚊.使得猪圈生境中,蚊虫密度比其他生境高出8~20倍,每灯每夜高达251.27只,其中三带喙库蚊为187.80只.而农户生境下,不仅适合三带喙库蚊、中华按蚊和骚扰阿蚊等野栖型蚊种的繁殖,也适合家栖型蚊种致倦库蚊的繁殖,蚊虫密度为每灯每夜39.94只. 上述结果提示,在进行蚊媒防控时,根据不同时间和空间优势种的生物学特性,可以采取有针对性的综合防控措施.一般来说,蚊虫的防制以孳生地管理为主,如对小型容器翻盆倒罐、大中型水体加强疏通和治理,结合市民防蚊宣传教育,能达到显著降低蚊虫密度的防制效果[27];其次是加强防蚊,如安装纱门纱窗和使用蚊帐;最后考虑使用苏云金芽孢杆菌类的生物杀虫剂和拟除虫菊酯、氨基甲酸酯类的化学杀虫剂,尽量减少杀虫剂的使用,以免造成环境的污染和蚊虫抗药性的升高. 本研究诱蚊灯法采用的是功夫小帅牌诱蚊灯,该灯的主要工作原理是以蓝紫光对蚊虫进行诱捕,对夜间活动频繁的蚊种如三带喙库蚊、中华按蚊等捕捉效率更高[28].但该灯缺乏CO2诱捕功能,且伊蚊活动高峰期主要在下午3点至7点[29],对伊蚊抓捕效率非常低,不能真实反应自然中实际的伊蚊种类和数量,有一定的局限性.2 结果
2.1 蚊虫种类与数量

2.2 蚊类季节性变化规律

2.3 多样性分析

2.4 群落聚类分析
3 讨论
猜你喜欢
牡丹(2021年23期)2021-12-11 20:05:38
大自然探索(2021年9期)2021-11-07 21:12:58
戏曲研究(2018年4期)2018-05-20 09:37:56
东方少年·阅读与作文(2018年6期)2018-05-14 14:32:43
扬子江(2017年4期)2017-07-19 08:41:32
环境昆虫学报(2016年4期)2016-08-23 03:13:46
新疆大学学报(哲学社会科学版)(2015年4期)2015-10-12 01:17:54
动物医学进展(2015年11期)2015-06-11 02:21:40
读者·校园版(2014年18期)2014-05-14 13:11:34
寄生虫与医学昆虫学报(2013年4期)2013-11-19 03:45:28
