
黑鲷ELO基因编码蛋白结构与功能的生物信息学分析
2021-06-02 10:30:10许广平张志勇任忠宏张志伟
生物技术进展 2021年3期
许广平, 张志勇, 任忠宏, 张志伟
江苏省海洋水产研究所, 江苏省海水鱼类遗传育种重点实验室,江苏 南通 226007
黑鲷(Acanthopagrusschlegelii)隶属鲈形目(Perciformes)鲷科(Sparidae),俗称海鲋、黑加吉、海鲫、黑立等,具有营养价值高、抗逆性强且不作长距离洄游等特点,是我国沿海重要的海水养殖经济鱼类[1],但其在长江以北无法在室外自然越冬,每年进行室内越冬又耗时耗力[2],亟需展开耐低温品系培育。
高不饱和脂肪酸(highly unsaturated fatty acids,HUFA)具有供应能量、维持细胞膜结构与功能和基因调控等功能,在生物体的生命活动中起重要作用[3],在一定范围内,鱼体中HUFA含量与机体低温耐受呈正相关,即机体HUFA含量越多,机体低温耐受性越强[4]。脂肪酸延长酶(fatty acid elongase,ELO)是影响HUFA合成的关键酶之一,参与脂肪酸代谢,与膜脂代谢密切相关[5-7]。目前,ELO基因在鲷科鱼类中研究报道较少,且该基因编码的蛋白质在理化性质、结构与功能等方面分析尚不全面。通过对ELO基因编码蛋白的结构与功能分析,对其在黑鲷体内开展表达调控研究,有望培育黑鲷耐低温品系。
脂肪酸延长酶以长链脂肪酸延长酶(elongation of very long chain fatty acids,ELOVL)或ELO分别存在于生物体内,两者为垂直同源。一般ELO以基因家族(ELO1、ELO2和ELO3)存在于生物体内,如酵母、拟南芥等[8-9];但有时也以单独基因存在,如鱖鱼等[10]。ELOVL家族是在ELO家族的研究基础上发现的,存在于哺乳动物体内的不同组织中,家族共有7个成员(ELOVL1~ELOVL7)[11]。目前,在鱼类研究中利用反转录聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR)和cDNA末端快速扩增技术(rapid amplification of cDNA ends,RACE)克隆了鱖鱼等鱼类ELO基因cDNA全长,鳜鱼ELO基因编码294个氨基酸[10],鳜鱼ELO基因克隆和功能解析为进一步研究其HUFA合成能力、合成途径、调控机理及在不同鱼类中的分子进化机理奠定了基础。不同鱼类 ELO之间有较好的蛋白同源性,均含有高度保守的氧化还原中心组氨酸簇、内质网停留信号和跨膜区等特征结构功能区[10-12]。本研究以黑鲷、金头鲷、鱖鱼、斜带石斑鱼、鲤鱼等5种鱼类的ELO基因编码蛋白的氨基酸序列作为研究对象,通过序列比对和同源性分析,探讨5种鱼的亲源关系。再运用多种生物信息学方法,对黑鲷ELO基因编码蛋白的理化性质与组成结构进行分析和预测;对该蛋白质在细胞内的定位、与蛋白质功能调控和翻译修饰相关的作用位点进行预测和分析;对蛋白质功能预测及蛋白质相互作用进行分析;并进行了同源建模及三维结构预测,旨在通过研究确定黑鲷中ELO基因及其编码蛋白的主要功能与作用途径,从而达到对该蛋白质表达水平的调控,为提高黑鲷低温耐受性的研究提供理论基础。
1 数据与分析
1.1 数据来源
所用黑鲷ELO基因编码蛋白的氨基酸序列为本实验室和华大基因黑鲷全基因组测序并序列比对所得;金头鲷(Sparusaurata,登录号:ADD50001.1)、鱖鱼(Sinipercachuatsi,登录号: ACH53603.1)、斜带石斑鱼(Epinepheluscoioides,登录号: ACJ26847.1)、鲤鱼(Cyprinuscarpio,登录号: AER39746.1)4种鱼类ELO基因编码的完整氨基酸序列均来源美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)GenBank 数据库。
1.2 物种间ELO氨基酸序列同源性及进化关系分析
运用DNAman 8软件对黑鲷、金头鲷、鱖鱼、斜带石斑鱼、鲤鱼等5种鱼类的ELO蛋白进行氨基酸序列比对,并开展同源性和进化关系分析。
1.3 ELO蛋白结构与功能分析
参照王亚琪等[13-14]的方法,利用各种生物信息软件对黑鲷ELO基因编码蛋白进行理化性质(氨基酸组成、等电点、原子总数、正负电荷残基数、不稳定指数、摩尔消光系数、脂肪族氨基酸指数和总平均亲水性等)、疏水性、亚细胞定位、跨膜区、信号肽、剪切位点、蛋白磷酸化、糖基化、二级结构、结构域及三维结构等方面的预测与分析。相关生物信息软件见表1。
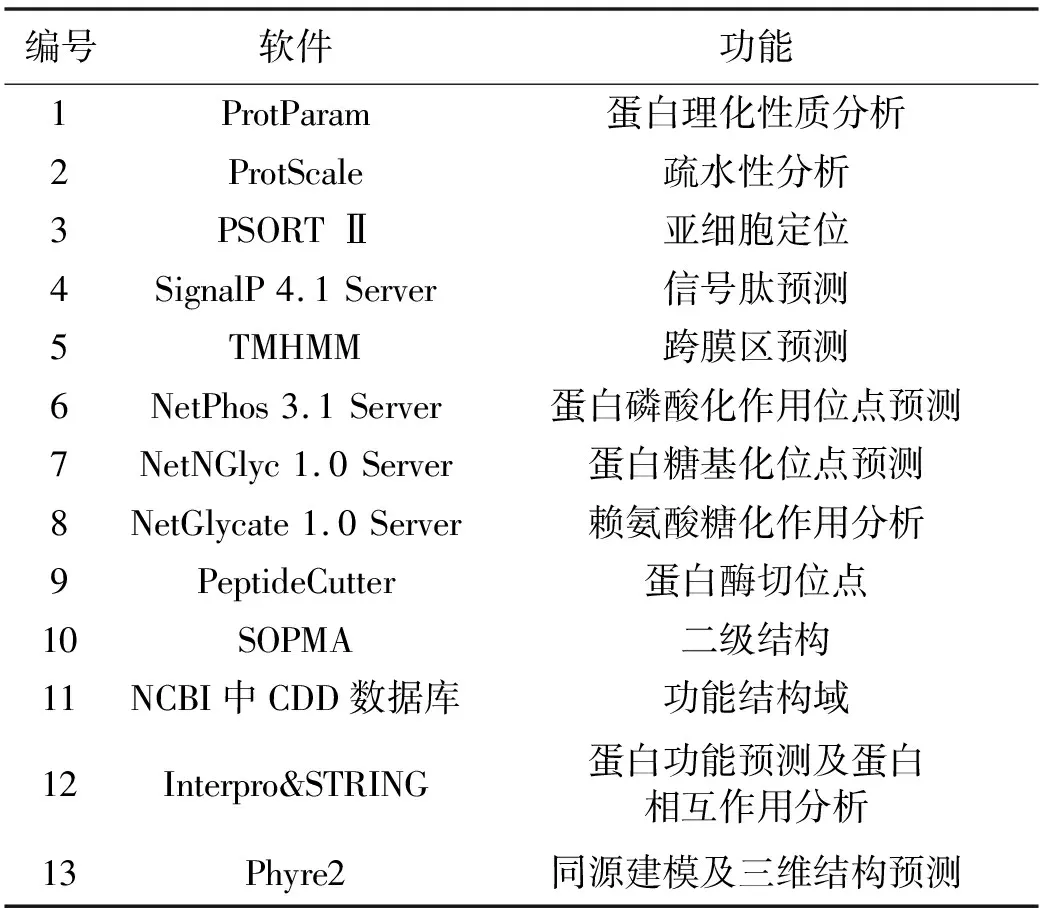
表1 本研究预测蛋白质结构与功能所应用的软件
2 结果与分析
2.1 物种间ELO氨基酸序列同源性及进化关系分析
将黑鲷、金头鲷、鱖鱼、斜带石斑鱼、鲤鱼的ELO氨基酸序列进行比对分析,发现黑鲷与金头鲷之间的序列同源性最高,在系统进化树中距离最近;与鲤鱼同源性最低,在系统进化树中距离最远(图1A)。进化聚类分析结果表明,与已知物种进化关系相比较,ELO基因构建的生物进化关系图谱基本与之相一致(图1B)。
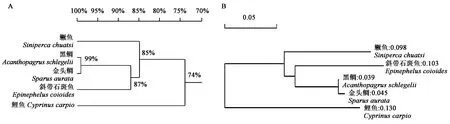
A:ELO氨基酸序列同源性;B:ELO氨基酸序列进化树
2.2 黑鲷ELO蛋白理化性质分析
基于黑鲷的ELO氨基酸序列,用ProtParam和ProtScale 工具中的Kyte &Doolittle法对ELO蛋白的理化性质(氨基酸组成、分子质量和理论等电点)进行分析。结果表明,黑鲷ELO蛋白由294个氨基酸残基组成,分子式为C1669H2403N403O410S18,原子总数为4 903个,相对分子质量为35 249.97。黑鲷ELO蛋白的氨基酸组成(图2):亮氨酸含量最高(10.5%),苯丙氨酸与酪氨酸次之(8.2%),半胱氨酸与丝氨酸最低(1.4%)。生物体内大部分苯丙氨酸氧化成酪氨酸,这2种氨基酸一起合成重要的神经递质和激素,参与机体的糖和脂肪代谢;同时,亮氨酸和丝氨酸在促进机体正常生长发育方面也起到至关重要的作用[4]。
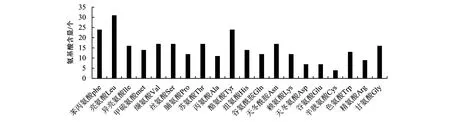
图2 黑鲷ELO蛋白各种氨基酸含量
ELO蛋白的理论等电点与不稳定系数分别为9.08和36.23,说明ELO蛋白为碱性蛋白质且性质稳定。在蛋白质疏水性分析中:正值代表疏水性,负值代表亲水性,分值(绝对值)代表其疏(亲)水性大小。ELO的总平均疏水指数为-0.49,整条氨基酸链中疏水性氨基酸残基少于亲水性氨基酸残基(图3),说明ELO蛋白为亲水性蛋白质,由此推测其为水溶性蛋白质。
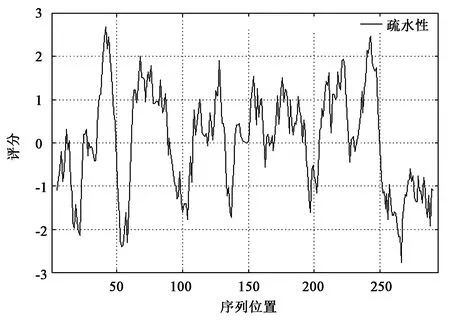
图3 黑鲷ELO蛋白疏水性分析结果
2.3 亚细胞定位预测
利用PSORT Ⅱ[15-17]工具中K-NN算法[18]对黑鲷ELO蛋白进行亚细胞定位,预测结果表明:ELO位于细胞质的概率最大,占整体的65.23%;其次是细胞核,概率为21.74%;位于线粒体内的概率是13.03%。说明ELO可能存在细胞质、细胞核和线粒体中,但在细胞质内发挥生物学作用的可能性最大。
2.4 信号肽和跨膜区预测
利用黑鲷的ELO氨基酸序列,采用SignalP 4.1 Server程序对ELO信号肽进行分析预测。C值(C-score)和S值(S-score)分别指原始信号肽裂解位点记分和信号肽评分,分别表示出现裂解位点可能性及氨基酸位于信号肽区域概率;Y值(Y-score)是最有可能的信号肽裂解位点;D值(D-score)表示S均值和最大Y值加权平均值,可以区分信号肽和非信号肽[19]。ELO蛋白预测D值为0.132(小于0.450),表明ELO蛋白不含信号肽,且不属于分泌性蛋白(图4)。
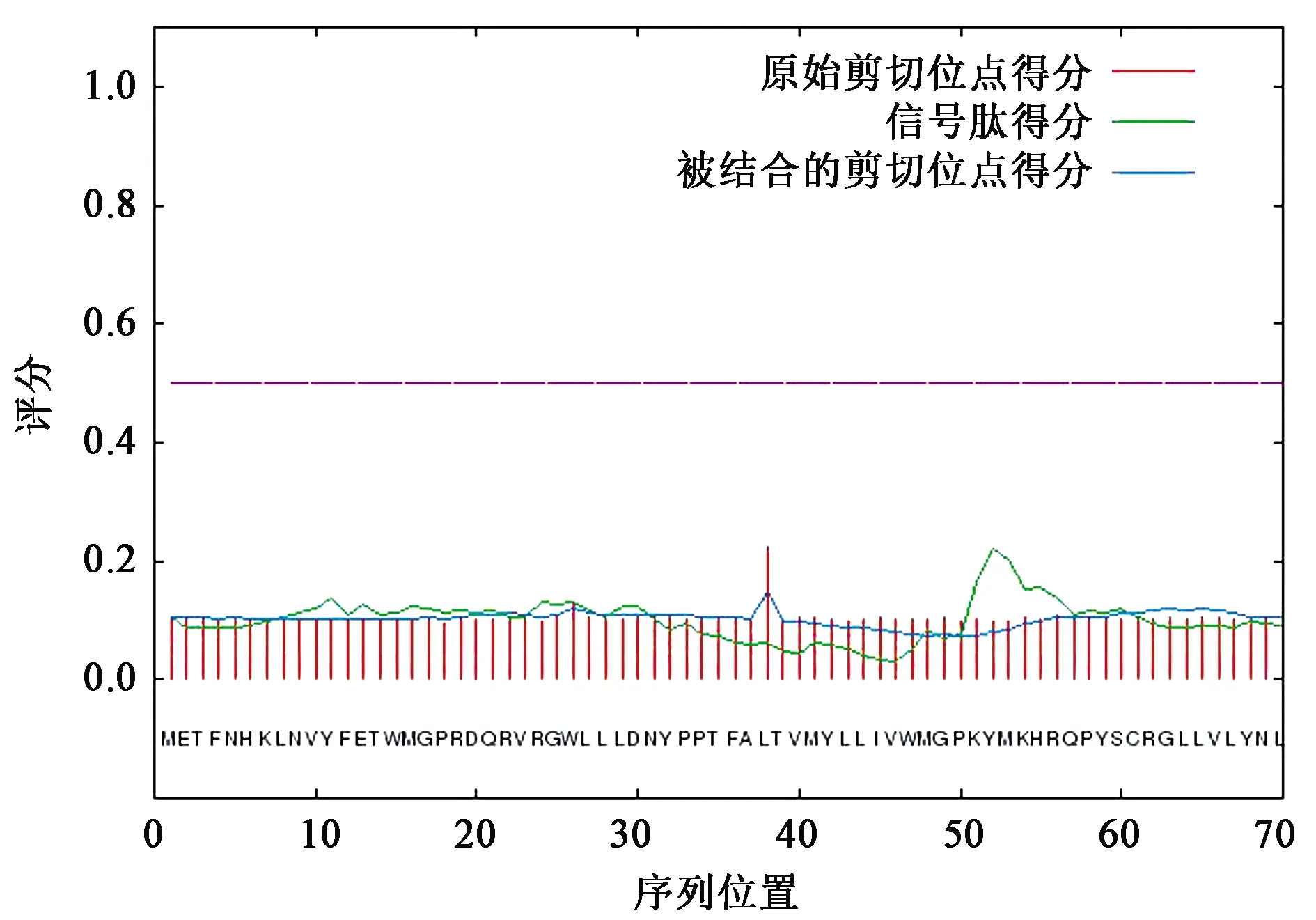
图4 黑鲷ELO基因编码蛋白的信号肽分析
使用TMHMM工具[20]在线分析发现黑鲷ELO有7个跨膜区(跨膜螺旋),非跨膜螺旋区(ExpAA)预测值为149.03164(大于18),说明预测蛋白为跨膜蛋白,蛋白N端位于膜内一侧的可能性是0.034 97%,总体来看该序列位于膜外,即ELO基因所编码蛋白质为跨膜蛋白。预测结果显示,黑鲷ELO可能在25~47、110~132、176~198、231~250氨基酸的位置形成4个膜外到膜内的跨膜区域;在68~90、139~161、205~227氨基酸的位置形成3个膜内到膜外的跨膜区域,表明黑鲷ELO蛋白可能行使细胞内外信号转导的作用(图5)。
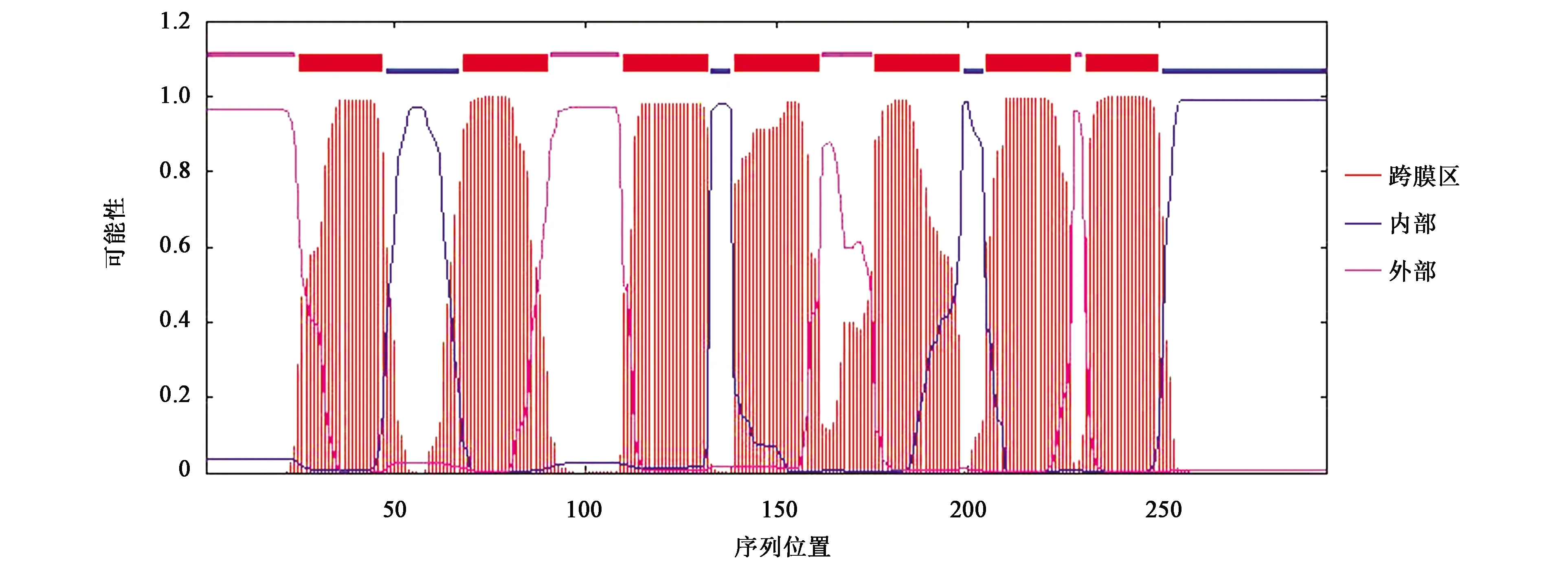
图5 黑鲷ELO蛋白质跨膜区预测结果
2.5 蛋白质翻译后修饰分析
蛋白质翻译后修饰一般指蛋白质磷酸化、糖基化及赖氨酸糖化作用等。蛋白质磷酸化是控制蛋白质是否发挥功能的开关,是细胞里面对蛋白质功能进行调控的普遍机制。而糖基化是一种翻译修饰,有些蛋白质发挥作用需要糖分子、糖链等与别的分子相互作用,是细胞产生不同功能蛋白质的一种形式。对这些修饰作用的研究,对了解蛋白质的完整功能与功能发挥具有重要意义[21]。利用NetPhos 3.1 Server软件[22]预测黑鲷ELO蛋白磷酸化作用位点,结果表明,该蛋白质具有多个不同的磷酸化作用位点,其中包括4个酪氨酸(Tyr;y-52、y-201、y-233、y-250)、4个苏氨酸(Thr;t-3、t-203、t-253、t-280)和11个丝氨酸(Ser;s-76、s-101、s-149、s-171、s-174、s-215、s-247、s-258、s-260、s-269、s-282)磷酸化作用位点(图6)。其中,s-282的分数为0.976,表示极有可能的磷酸化位点;而s-149(0.503)、s-171(0.503)、y-201(0.500)、t-203(0.508)、y-233(0.508)的分数仅略高于阈值(0.500),表明该5个位点为真正的磷酸化位点的可能性极低。
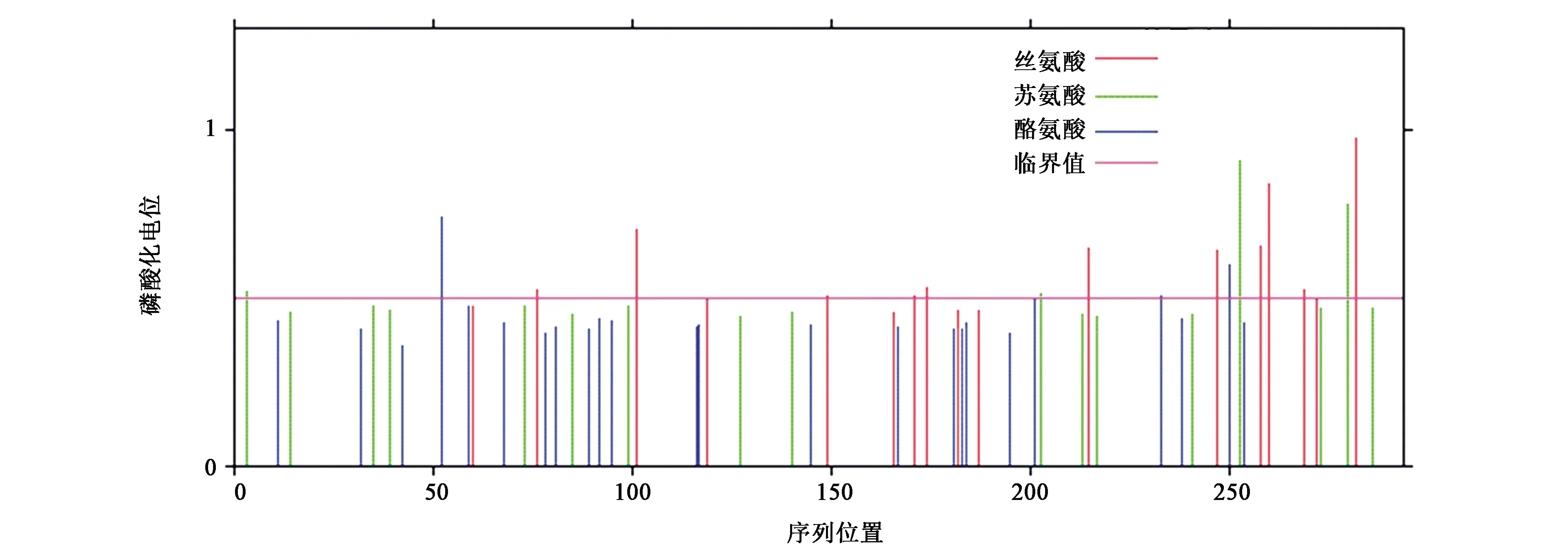
图6 黑鲷ELO基因编码蛋白磷酸化位点预测
利用NetNGlyc 1.0 Server与NetGlycate 1.0 Server在线分别预测ELO蛋白糖基化位点和赖氨酸糖化作用,并未发现糖基化位点,但有9个位点(Pos.7、Pos.54、Pos.108、Pos.120、Pos.134、Pos.199、Pos.200、Pos.263、Pos.290)可能存在赖氨酸糖化作用(图7)。
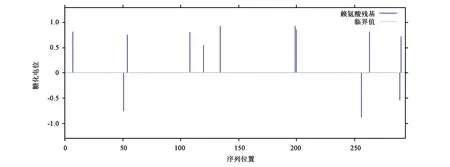
图7 黑鲷ELO蛋白赖氨酸糖化作用预测结果
2.6 ELO蛋白酶切位点预测
利用PeptideCutter工具预测ELO蛋白酶切位点,对该蛋白质有剪切作用的酶和裂解作用的酸如下:精氨酸-C 蛋白水解酶(Arg-C proteinase)、Asp-N 内切酶(Asp-N endopeptidase)、Asp-N内切酶 + N端亮氨酸(Asp-N endopeptidase + N-terminal Glu)、BNPS粪臭素(BNPS-Skatole)、溴化氢(CNBr)、高特异性糜蛋白酶(chymotrypsin-high specificity)、低特异性糜蛋白酶(chymotrypsin-low specificity)、梭菌蛋白酶(clostripain)、甲酸(formic acid)、谷氨酰基肽链内切酶(glutamyl endopeptidase)、羟胺(hydroxylamine)、氧代苯甲酸(iodosobenzoic acid)、LysC、LysN、NTCB(2-nitro-5-thiocyanobenzoic acid)、胃蛋白酶(pepsin,pH 1.3)、胃蛋白酶(pepsin, pH>2)、脯氨酸肽链内切酶(proline-endopeptidase)、蛋白水解酶K(proteinase K)、staphylococcal peptidase Ⅰ、嗜热菌蛋白酶(thermolysin)、胰蛋白酶(trypsin)。
2.7 ELO二级结构及结构域预测
利用SOPMA在线程序预测ELO蛋白二级结构,发现该蛋白质的二级结构以α-螺旋为主,由137个氨基酸残基组成,占整个二级结构的46.6%;无规则卷曲由92个氨基酸构成,占整体的31.29%;延伸带和β-转角分别由55、10个氨基酸残基构成,在整体中所占比例分别为18.71%和3.4%(图8)。

注:h: α-螺旋; e: 伸展链; c: 无规卷曲; t: β-转角。
应用 NCBI 中的 CDD 数据库(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测蛋白质功能结构域。预测结果如图9所示,结果表明: 黑鲷ELO基因编码蛋白的第28~256个氨基酸区域为ELO超家族(ELO superfamily)结构域。

图9 黑鲷ELO基因编码蛋白结构域预测
2.8 功能预测及蛋白质相互作用
利用Interpro预测ELO功能,发现该蛋白质在GO数据库中的注释信息分3个部分:生物学进程、分子功能和细胞定位(表2)。该蛋白质主要功能为脂肪酸延伸和超长链脂肪酸合成,而蛋白质行使功能不仅与自身特性有关,同时还与相互作用的蛋白质相关。利用STRING[22]分析其他蛋白质与ELO蛋白相互作用信息,选择比对生物为斑马鱼(Daniorerio),发现黑鲷ELO蛋白与斑马鱼elovl 5相似度最高(bitscore和e-value分别为417.2和1.1e-116),蛋白同源性74.83%。斑马鱼elovl 5蛋白与其他蛋白质相互作用关系如图10所示,10个蛋白质与其存在直接的相互作用关系,分别为srbd1、ptplb、sycp2l、elovl2、hsd17b12b、hsdd17b12a、scd、pcyt2、srebf1和fads2。斑马鱼elovl 5蛋白在GO数据库中的注释信息体现出:elovl 5蛋白影响脂肪酸和雌激素合成,且在分子功能上与多种酶活性有关(表3)。由此推测,ELO蛋白除了合成脂肪酸,可能还影响雌激素合成,与srbd1等10种蛋白质相互作用。
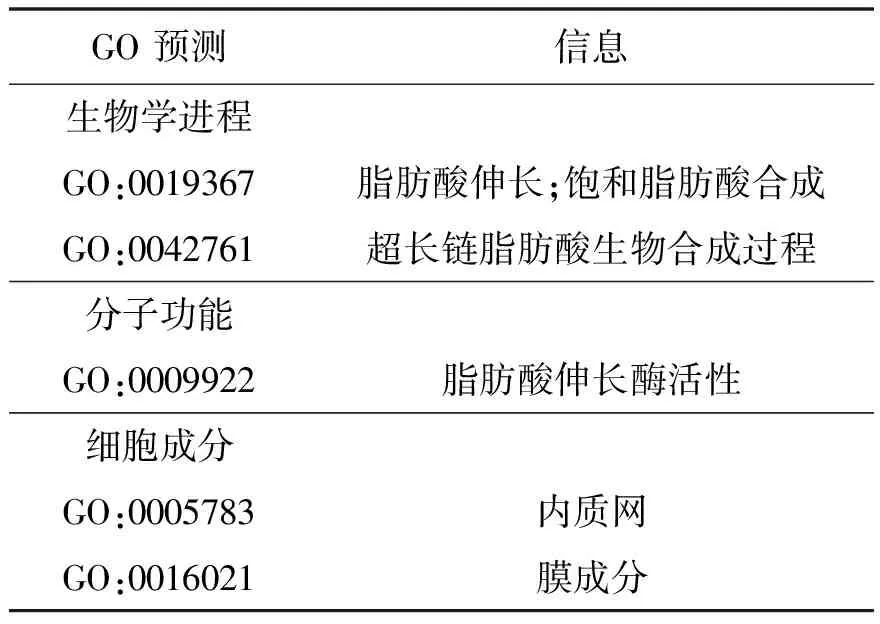
表2 ELO蛋白质在GO数据库中的注释信息

表3 Elovl 5蛋白质在GO数据库中的注释信息
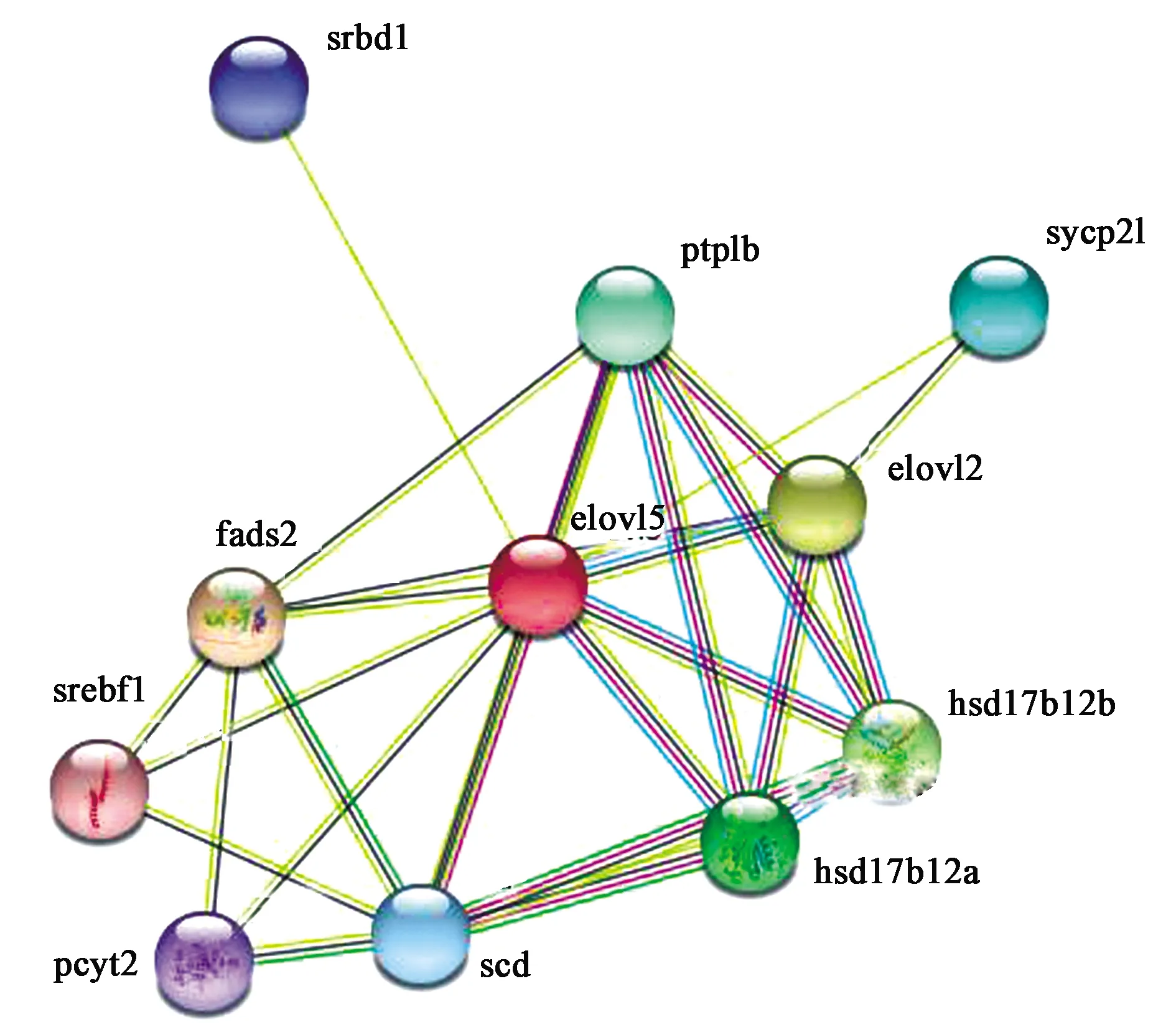
图10 STRING数据库斑马鱼Elovl 5蛋白相互作用关系
2.9 ELO蛋白同源建模及三级结构预测
一般预测蛋白质三维结构有3种方法: 同源建模法(homology modeling)、线串法(threading)和重头预测法(abinitio)。其中同源建模是使用最广泛的一种方法,该方法的优点在于:待测蛋白与模板序列的同源性越高,预测结果越准,当待测蛋白与模板序列同源性高于60%时,预测结果与实验实际测得结果非常接近。
利用Phyre2在线工具中Normol模式对黑鲷ELO进行同源建模和三维结构预测,预测出黑鲷ELO蛋白的三维结构如图11所示,该三维模型以c5xpdA为模板,覆盖范围(同源性)为31%,略高于30%,可信度为31.6%,一定程度上反映了黑鲷ELO蛋白结构。
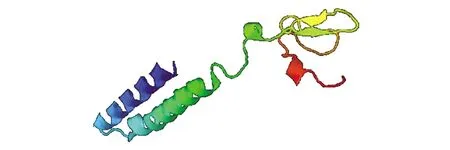
图11 黑鲷ELO基因编码蛋白的三级结构
3 讨论
黑鲷ELO氨基酸序列与其他鱼类同源性均较高,推测不同物种中该基因编码的蛋白质活性相似。本研究表明,与金头鲷、鳜鱼、鲤鱼、斜带石斑鱼4种不同生境的鱼类相比较,同源性聚类分析及系统树分析均显示黑鲷与金头鲷亲缘关系最近,与鲤鱼亲缘关系最远,这与已有研究结论一致[24]。金头鲷与黑鲷同属鲷科鱼类,在亲缘关系上比其他3种鱼类更近,该结论与传统分类学相吻合,但有学者认为鱼类的生活环境可能影响ELO的进化[3]。不同物种间同种蛋白质的同源性高,其在功能上也具有相似性[25]。这可能是由于不同物种间ELO蛋白结构的高稳定性和序列的强保守性所致。本课题组在通过对黑鲷、真鲷及其杂交后代CDS序列的基因密码子偏好性进行聚类时发现4种鱼基因编码区在密码子使用上呈现高度一致性[26]。
黑鲷因其低温耐受性较差,在长江以北无法在室外自然越冬,每年开展室内越冬又耗时耗力[2]。有研究发现鱼类耐低温可能与ELO基因表达相关[27]。本文运用生物信息学工具对黑鲷ELO蛋白理化性质、结构、分子功能及蛋白质相互作用等进行了分析。结果表明:黑鲷ELO基因编码蛋白由294个氨基酸组成,亮氨酸含量最高,存在多种二级结构,其中以α-螺旋为主,与斜带石斑鱼、拟南芥等分析结果一致[3,8]。ELO蛋白在拟南芥中亚细胞定位至细胞质或内质网,其在黑鲷亚细胞定位至细胞质的概率最高,且无信号肽,推测该蛋白质为一种存在于细胞质的非分泌蛋白。总体来说,ELO蛋白为碱性、小分子、稳定性蛋白质,存在1个ELO超家族结构域,与HUFA 合成的另1个关键酶脂肪酸去饱和酶(fatty acid desaturase,FAD)存在显著差异[3]。分析表明,ELO蛋白分子中存在多个磷酸化作用位点,证明其具有合成高度不饱和氨基酸功能,可以促进细胞生长;该蛋白分子中也存在多个赖氨酸糖化位点,可用于ELO蛋白的翻译后修饰,通过与别的分子相互作用,形成一种新的功能,赖氨酸糖化对于维持蛋白质的结构和功能都起到重要的作用。此外,本研究通过对黑鲷ELO结构及功能预测分析,为探讨该基因在黑鲷低温耐受机制方面提供了相关理论基础。蛋白质相互作用关系分析中,以斑马鱼中Elovl 5基因为模板,推测srbd1等10个蛋白质与ELO可能存在直接的相互作用关系,有可能影响雌激素合成,并在分子功能上与多种酶活性有关[28]。
本研究通过生物信息学方法对不同鱼类ELO基因编码蛋白序列进行进化聚类分析,以及对黑鲷该蛋白理化性质、亚细胞定位、跨膜区、信号肽、酶切位点、蛋白磷酸化、糖基化、二级结构、结构域及三维结构等方面进行预测与生物信息学分析,推测该蛋白质功能与低温耐受性相关。下一步将继续开展该基因在不同组织和不同发育时期差异表达分析,以确定其靶标组织和关键的表达时期,了解黑鲷中脂肪酸合成的途径和机制;进一步开展黑鲷抗寒性状相关的基因表达调控研究,最终为黑鲷耐低温新品种的培育工作提供理论依据。
猜你喜欢
河北科技大学学报(2023年5期)2023-11-09 01:44:44
上海金属(2021年6期)2021-12-02 10:47:20
昆明医科大学学报(2021年3期)2021-07-22 07:40:04
中国洗涤用品工业(2019年4期)2019-05-11 09:27:18
生物学通报(2019年3期)2019-02-17 18:03:58
中成药(2018年1期)2018-02-02 07:20:05
浙江农业学报(2017年3期)2017-04-08 02:39:02
动物医学进展(2015年10期)2015-12-07 05:46:19
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:23
食品工业科技(2014年7期)2014-03-11 18:15:07
