
Metabolic and biological changes in children with obesity and diabetes
2021-05-29 01:52:36ShirouMatsumotoTomomiNakamuraFusaNagamatsuJunKidoRiekoSakamotoKimotoshiNakamura
World Journal of Meta-Analysis 2021年2期
Shirou Matsumoto, Tomomi Nakamura, Fusa Nagamatsu, Jun Kido, Rieko Sakamoto, Kimotoshi Nakamura
Shirou Matsumoto, Fusa Nagamatsu, Jun Kido, Kimotoshi Nakamura, Department of Pediatrics, Faculty of Medical Sciences, Kumamoto University, Kumamoto 860-8556, Japan
Tomomi Nakamura, Rieko Sakamoto, Department of Perinatal Care Unit, Kumamoto University Hospital, Kumamoto University, Kumamoto 860-8556, Japan
Abstract The World Health Organization has stated that obesity in childhood is one of the most serious public health challenges of the 21st century.Overweightness and obesity in early childhood lead to a higher risk of overweightness and obesity in adulthood, thus conferring an increased risk of chronic inflammatory conditions, including type 2 diabetes mellitus, cardiovascular diseases, non-alcoholic fatty liver disease, and some cancers.Therefore, metabolome analysis, targeted at screening and intervening in childhood obesity, is very important.Recent studies have indicated that amino acid and lipid metabolism could influence metabolic pathways in children with obesity.For this review, we searched clinical data addressing metabolomic profiles and insulin resistance (IR) in children with obesity from inception to February 2021 in Medline, Web of Science, and Scopus.According to our search, branched-chain amino acids (BCAAs), aromatic amino acids, and acylcarnitines have reportedly been associated with IR as biomarkers for diabetes in children.BCAAs, tyrosine, and phenylalanine could be predictors of the future development of diabetes in nondiabetic subjects.In addition, it is well known that insulin regulates BCAA metabolism, and BCAA is a biomarker for IR.To interpret the mechanism behind metabolic changes in obesity, it is very important to understand the pathways and combinations related with amino acid, lipid and glucose metabolism.In this review, we summarize studies on metabolic changes to understand metabolomics in children with obesity.
Key Words: Children with obesity; Diabetes mellitus; Insulin resistance; Amino acids; Acylcarnitine; Metabolomics
INTRODUCTION
The number of overweight children under the age of 5 years in 2016 was estimated to be over 41 million worldwide.Symptomatic pediatric lifestyle-related diseases are present in 5%-15% of children with obesity and increase at onset of adolescence, after the late elementary grade.Although there are many published mechanisms underlying the metabolic profile, those of children with obesity remain only partially understood.Obesity in childhood is a risk factor for hyperlipidemia, hypertension, type 2 diabetes mellitus (T2DM), and cardiovascular disorders in early adulthood.Moreover, severely overweight children develop pediatric lifestyle diseases, such as hyperlipidemia, diabetes, and fatty liver[1,2].Between 1980 and 2014, the number of adults with diabetes increased from 108 million to 422 million[3], with T2DM accounting for > 90% of these cases[4].Early detection and intervention of obesity in childhood is important to prevent both pediatric and adult lifestyle diseases[5].
Recently, investigations have been conducted on early screening and diagnosis using metabolomics, which avails much information and indications.About 141 reliable articles related to metabolomics in obesity have been published[6].Samples were collected from blood (serum and plasma) and urine.Detection methods varied, including liquid chromatography tandem-mass spectrometry, gas chromatography tandem mass spectrometry, time-of-flight mass spectrometry, and nuclear magnetic resonance spectroscopy.Detected metabolites, which are related to T2DM and insulin resistance (IR), included amino acids (upregulated: isoleucine, alanine, proline, glutamate, palmitic acid, 2-aminoadipic acid, and lysine; downregulated: glycine, serine, and citrulline), lipids (palmitic acid), and others (propionyl carnitine, carnitine, and 2-aminoadipic acid).Evidently, all metabolites are related to each other in a complex manner; however, the core factor seems to be amino acids, according to the literature.In this review, we focus on amino acid metabolism in obesity and explain important mechanisms that have been established to date.
SYSTEMATIC REVIEW OF METABOLOMICS IN PEDIATRIC OBESITY
We searched clinical data addressing metabolomic profiles and IR in children with obesity from inception to February 2021 in Medline, Web of Science, and Scopus.The search strategy was based on medical subject headings terms.Variant combinations of the following terms were applied during the search: “childhood obesity,” “obese children,” “pediatric obesity,” “IR,” “metabolomics,” “amino acids,” and “lipids”.Inclusion and exclusion criteria have been described in a previous study (39 Using Metabolomic Profiles as Biomarkers for Insulin Resistance in Childhood Obesity: A Systematic Review).QUADOMICS was used to evaluate the methodological quality of the included studies.We performed literature selection according to the PRISMA protocol shown in Figure 1[7].In this review, we summarize not only systematically reviewed data but also basic studies that help in understanding metabolic changes in children with obesity.We were able to detect 13 reliable clinical studies after systematic selection (Table 1).
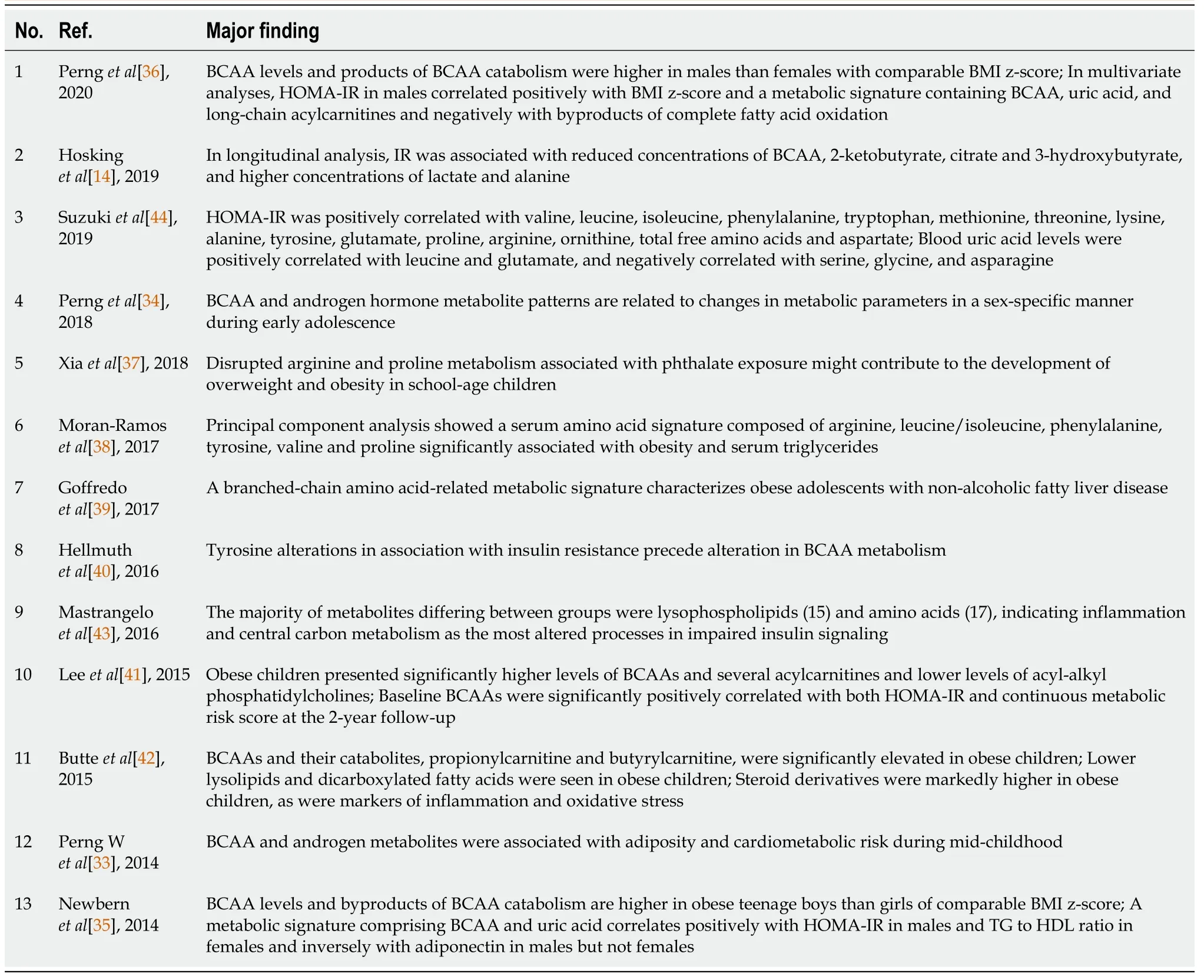
Table 1 Characteristics of study findings and identified metabolomic profiles in 13 clinical studies
AMINO ACID AND INSULIN PROFILES DURING CHILDHOOD IN HEALTHY CHILDREN
In healthy children, total amino acid concentration tends to increase up to 18 years of age due to the growth and construction of their own bodies.Plasma amino acid concentrations during childhood are summarized in Table 2[8].Nine amino acids, namely, alanine, arginine, asparagine, methionine, ornithine, phenylalanine, proline, threonine, and tyrosine, decrease during the first year after birth.After 1 year of age, the concentrations tend to increase throughout childhood and adolescence.Nine other amino acids, namely, cystine, glutamine, glycine, histidine, isoleucine, leucine, lysine, tryptophan, and valine, demonstrate a steady increase throughout infancy, childhood, and adolescence.Five amino acids, namely, aspartic acid, citrulline, glutamic acid, serine, and taurine, show a steady concentration throughout childhood.In children with obesity and without IR, total plasma free amino acids demonstrate lower levels than those in healthy controls.
Insulin exhibits change similar to those of total free amino acids, and puberty strongly influences IR, as described in Table 3[9].During puberty, insulin increase correlates with prepubertal levels.The lower insulin sensitivity observed in prepubertal girls is likely due to a higher growth hormone secretion in pubertal girls or differences[10,11] in insulin peripheral action in both boys and girls[12].Particularly in boys during puberty, it has been demonstrated that IR and cardiovascular risk increase[13].Serum insulin increases in association with age, body mass index (BMI), PS, IGF-1 level, and triglyceride (TG) level[9].In addition, Hoskinget al[14] found IR to be associated with lower concentrations of branched-chain amino acids (BCAAs), 2-ketobutyrate, citrate, and 3-hydroxybutyrate and higher concentrations of lactate and alanine.These data portray the biochemical consequences of IR as rapid metabolism, ketogenesis, and pyruvate oxidation during normal child growth and development.Hence, when children undergo metabolic analysis, these factors, which potentially influence metabolism in the body, should be considered thoroughly and matched sufficiently in evaluating the results.
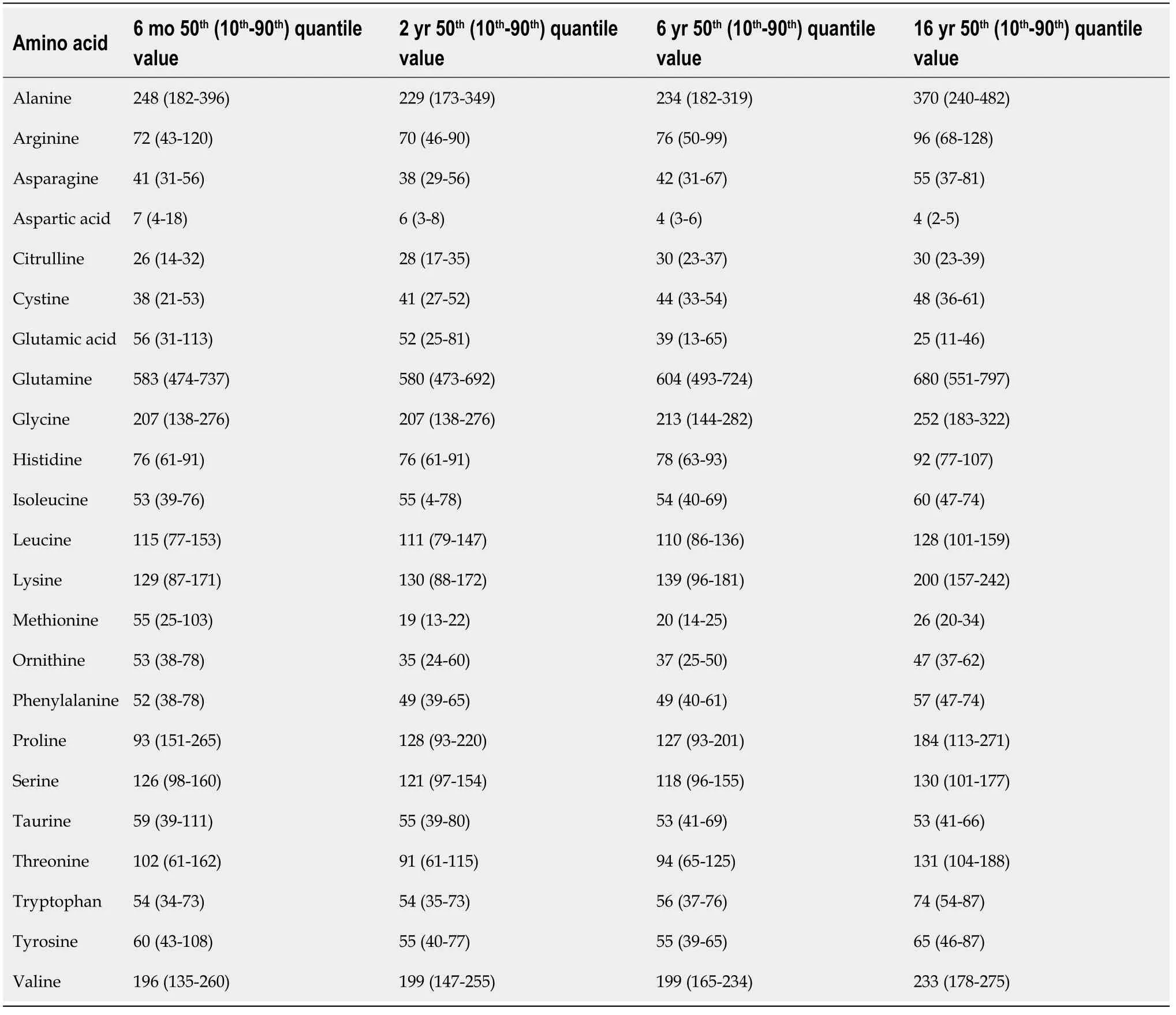
Table 2 Plasma amino acid values (μmol/L) during childhood

Table 3 Insulin, glucose/insulin, and homeostasis model assessment of insulin resistance levels during childhood
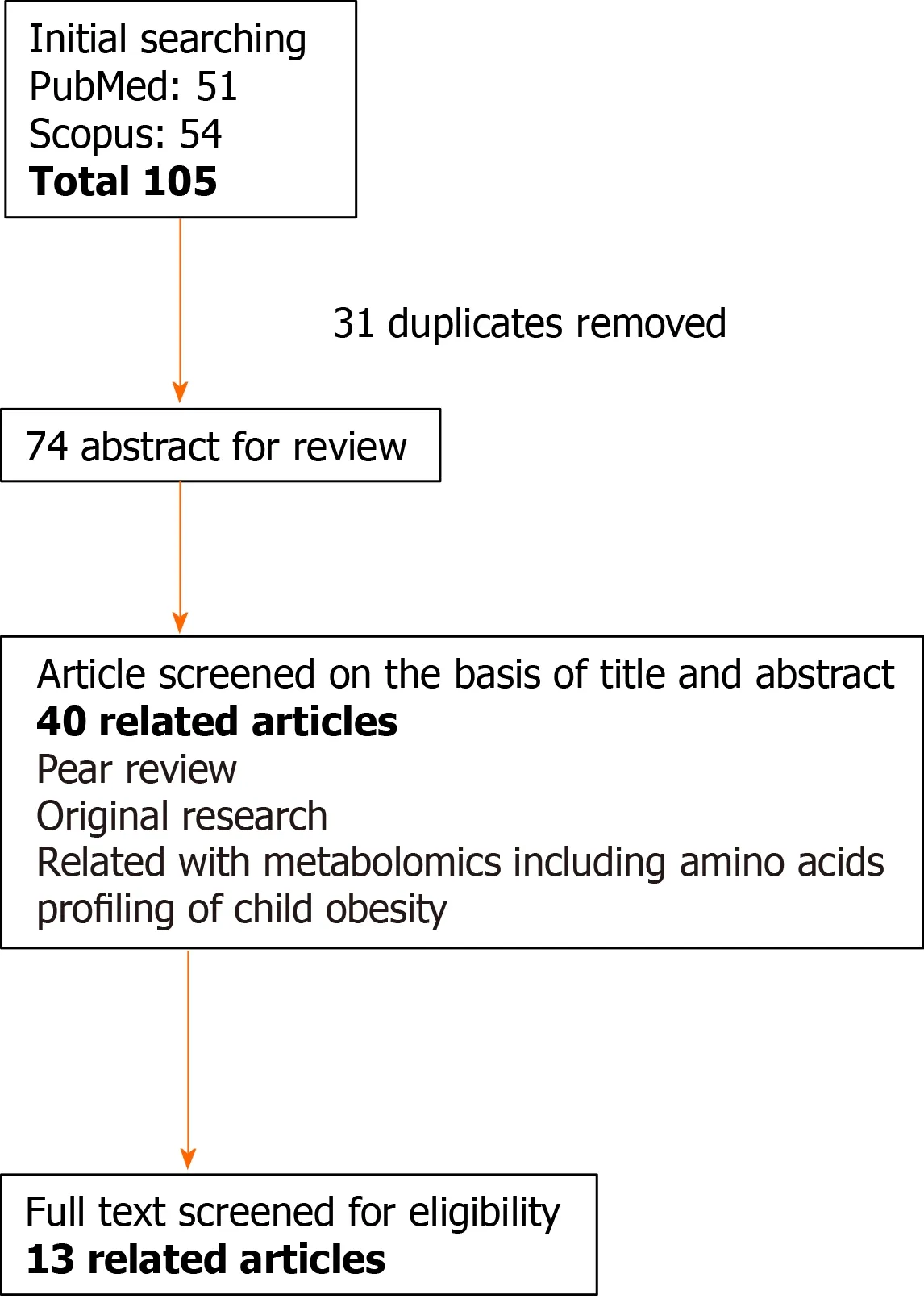
Figure 1 Flow diagram of the literature search and study selection for “metabolic/biological change in children with obesity and diabetes”.
INSULIN REGULATES GLUCOSE METABOLISM AND MODIFIES AMINO ACID METABOLISM
Insulin regulates glucose homeostasis mainly in the liver, muscle, and fat.Insulin inhibits gluconeogenesis and glycogenolysis, leading to induced glycogen storage[15].Insulin promotes the uptake, storage, and use of glucose in muscle and fat.Insulin is responsible for the induction of potassium transport in muscle, differentiation into adipocytes[16], production of androgens by ovaries[17], and retention of sodium by the kidney[18].Insulin performs all these functions by binding with a specific tyrosine kinase-type of transmembrane receptor.The binding of insulin to receptors leads to signal transduction, including insulin-receptor substrate-1 and PI3K, which activates Akt.Akt phosphorylates the Ser9 site of GSK-3b and inhibits its activity, leading to glycogen synthesis (Figure 2).
In addition, insulin, which acts as an inhibitor of protein degradation, reduces essential amino acid concentrations in the post-absorptive state.Insulin infusion leads to hypoaminoacidemia and does not stimulate protein synthesis in skeletal muscle or splanchnic tissues during the post-absorptive state[19].Insulin infusion decreases muscle protein degradation and does not enhance protein synthesis without amino acid intake[10,20,21].In growing children without IR, total plasma free amino acid levels are lower than those of adults (Table 3).
METABOLIC CHANGE IN PREDIABETES AND T2DM
Metabolic change in adults is summarized in Figure 3.BCAAs are known to be associated with obesity, a phenomenon first reported by Feliget al[22] in 1969.The plasma concentrations of BCAAs and aromatic amino acids (ArAAs), such as Phe and Tyr, were increased and that of glycine was decreased in individuals with obesity when compared to those of age- and sex-matched healthy controls.Consequently, Newgardet al[23] reported in 2009 that principal component analysis performed on datasets comprised plasma and urine amino acids, acylcarnitines, organic acids, and fatty acids.They identified a cluster of metabolites comprising BCAAs, ArAAs (Phe and Tyr), Glu/Gln, Met, and C3 and C5 acylcarnitines that are strongly associated with IR, suggesting a fundamental alteration of BCAA metabolism in insulin-resistant states.On the other hand, glycine was negatively associated with IR when measured using the homeostatic model assessment (HOMA) score[24] or by hyperinsulinemiceuglycemic clamp[20].In patients with T2DM, the concentrations of BCAAs can be up to 1.5-fold, or even 2-fold, higher than those in healthy subjects, with a mean change below 1.5-fold (1.2-1.3).These results are reproducible and statistically significant[25-27].
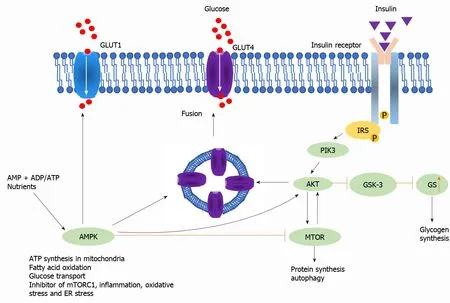
Figure 2 Insulin signaling pathway.

Figure 3 Insulin signaling pathway.B
The strong association of BCAAs with IR has been reported in several studies, including a cross-sectional study of subjects with metabolic syndrome and varying BMI[28], a study on Chinese and Asian-Indian subjects residing in Singapore in which BMI was controlled (average BMI of 24)[29], a study on subjects at extremes of insulin sensitivity in the Insulin Resistance Atherosclerosis Study[30], and studies on the effects of combined aerobic and resistance training in insulin-resistant subjects[31].In all of these studies, the BCAA-related factor was more correlated with IR than any lipid-related factor.
Another candidate metabolite is 2-aminoadipic acid (2-AAA), which is strongly correlated with incident T2DM, with subjects in the top quartile for this metabolite having a > 4-fold increase risk of disease[32].2-AAA proved to be an independent biomarker because its levels were not associated with BCAAs or ArAAs in this study.
The association between cardiometabolic diseases and certain lipid metabolites commonly measured in clinical chemistry laboratories - including TGs, cholesterol, and total nonesterified fatty acids - has long been recognized, leading some to conclude that cardiometabolic diseases are driven by perturbed lipid homeostasis.Application of metabolomics has led to a broader appreciation of metabolites that associate with these maladies and, in some cases, metabolites that predict intervention outcomes and future disease development.
METABOLIC CHANGE IN CHILDREN WITH OBESITY AND IR
Pernget al[33] investigated plasma metabolic profiling in 262 children (6-10 years of age).They found that BCAA-related and androgen-hormone patterns were higher in obese than in lean children.Both patterns were associated with adiposity and worse cardiometabolic profiles.Each increment in the BCAA and androgen pattern scores corresponded with a 6% (95% confidence interval (CI): 1%-13%) higher HOMA-IR.Children of mothers with obesity had higher BCAA scores than in those of mothers without obesity.Pernget al[34] also reported a BCAA pattern corresponding with a 4.82 (95%CI: 0.92-8.71) mg/dL decrease in fasting glucose in boys.In girls, the BCAA pattern was associated with an increase in TGs [4.17 (0.03-8.32) mg/dL].The androgen pattern was associated with decreased leptin [-2.35 (-4.34 to -0.35) ng/dL] and increased C-reactive protein [0.28 (0.03-0.54) mg/dL] in girls.These relationships did not differ by baseline BMI percentile.Sex differences in biomarkers were reported by Newbernet al[35].They investigated metabolome profiles in 82 adolescents with obesity.BCAA levels and byproducts of BCAA catabolism were higher in teenage boys with obesity than in girls of comparable BMI z-scores.A metabolic signature comprising BCAA and uric acid correlates positively with HOMA-IR in males and TGto-high-density lipoprotein ratio in females and inversely with adiponectin in males but not females.
Pernget al[36] also investigated metabolomic profiles of four overweight/obesity and metabolic risk phenotypes among 524 adolescents aged approximately 13 years.Their data demonstrated that long-chain fatty acids, BCAAs, acylcarnitines, diacylglycerols, and steroid hormones differed by weight status and metabolic phenotype.Xiaet al[37] investigated urinary metabolic analysis in 69 children with overweight/obesity and 80 lean children.They discovered that mono-n-butyl phthalate (MnBP) exposure was significantly associated with overweight/obesity in children and the disrupted arginine and proline metabolism might have played an important role in mediating the putative obesogenesis of MnBP.Moran-Ramoset al[38] investigated data from the Obesity Research Study for Mexican Children conducted in Mexico City and included a case control (n= 1120), cross-sectional (n= 554), and longitudinal study (n= 301) of 6-12-year-old children.Principal component analysis presented a serum amino acid profile composed of arginine, leucine/isoleucine, phenylalanine, tyrosine, valine, and proline significantly associated with obesity [odds ratio (OR) = 1.57; 95%CI: 1.45-1.69,P= 3.84 × 10-31) and serum TGs (β = 0.067,P= 4.5 × 10-21).In the longitudinal cohort, the amino acid profile was associated with serum TG and the risk of hypertriglyceridemia after 2 years (OR = 1.19; 95%CI: 1.03–1.39,P= 0.016).They found that an amino acid profile is an independent risk factor for future hypertriglyceridemia in childhood.Goffredoet al[39] investigated 180 metabolites in 78 adolescents with obesity with (n= 30) or without (n= 48) non-alcoholic fatty liver disease (NAFLD) assessed by magnetic resonance imaging.Adolescents with NAFLD had higher plasma levels of valine (P= 0.02), isoleucine (P= 0.03), tryptophan (P= 0.02), and lysine (P= 0.02) after adjustment for confounding factors.BCAAs were negatively correlated with peripheral and hepatic insulin sensitivity.Higher baseline valine levels predicted an increase in hepatic fat content at follow-up (P= 0.01).Hellmuthet al[40] studied 80 children with obesity before and after a 1-year lifestyle intervention program.Only tyrosine was significantly associated with HOMA (P< 0.05) at baseline and end, and with change during the intervention (P< 0.05).Leeet al[41] analyzed 186 plasma metabolites in 109 Korean boys at baseline and after 2 years.Significantly higher levels of BCAAs and several acylcarnitines were observed in children with obesity and as well as lower levels of acyl-alkyl phosphatidylcholines.Baseline BCAAs were significantly positively correlated with both HOMA-IR and continuous metabolic-risk score at the 2-year follow-up.Baseline BCAA concentration was identified as a predictor of future risk of IR and metabolic syndrome.Butteet al[42] analyzed the metabolic profiles of 353 Hispanic children without obesity and 450 with obesity.BCAAs, BCAA-catabolites, propionylcarnitine, and butyrylcarnitine were significantly elevated in children with obesity.

Figure 4 Metabolic change in children with obesity.
Mastrangeloet al[43] reported in 2016 on the metabolic profiles of prepubertal children with obesity.They found that BCAAs, ArAAs, alanine, ketoisocaproic acid, pyruvate, and C3 and C4 acylcarnitines increased in children both with obesity and IR (Figure 4).In addition, free carnitine and 3-hydroxyutyrate decreased in children presenting obesity with IR.Because ArAAs potentially compete with BCAAs for transport into mammalian cells, their elevation in serum may indicate a disruption in their transportation into the brain, leading to an excess of BCAAs.Suzukiet al[44] reported in 2019 about relationship among amino acids, lipids, and glucose metabolism.In this study, HOMA-IR was positively correlated with BCAAs, ArAAs, Ala, and Glu/Gln, with statistical significance[28].The mechanism underlying this profiling has not been determined; however, a hypothesis has been suggested.Pozefskyet al[45] suggested that impaired insulin activity and decreased utilization of amino acids in the muscles increased plasma BCAA levels owing to reduced uptake of BCAA in the muscles in lifestyle-related diseases.Newgard[46] reasoned that the increased circulating blood BCAA in obese and insulin-resistant subjects partly results from a decline in amino acid (AA) catabolism in their adipose tissue.Readily usable glucose and lipid substrates are considered to obviate the need for AA catabolism in adipose tissue by downregulation of the BCAA catabolic enzymes through the suppression of peroxisome proliferator-activated receptor-γ signaling in such metabolic adaptations.Würtzet al[47] reported that gluconeogenesis substrates, including Ala, increased in adults with impaired glucose tolerance.Moreover, Shimizuet al[48] reported that depletion of plasma Ala serves as a cue to increase plasma fibroblast growth factor 21 values and enhance liver-fat communication, resulting in the activation of lipolytic genes in adipose tissues.Increases in circulating BCAAs in obesity result in part from decreased rates of their oxidation in adipose tissue, due to coordinated transcriptional suppression of all the BCAA catabolic enzymes as well as from increased phosphorylation and inactivation of the branched-chain ketoacid dehydrogenase (BCKDH) complex in the liver, such that fewer BCAAs are taken up from the blood.Rodent models of obesity exhibit increased hepatic expression of the inhibitory BCKDH kinase, BDK, and reduced expression of the activating BCKDH phosphatase, PPM1K[49].Transplantation of microbiota from humans with obesity into germ-free mice causes a significant increase in circulating BCAAs and obesity[50].Hence, gut microbiota is a candidate donor of BCAAs.BCAA biosynthetic pathways are induced in the gut microbiota of obese compared to that of lean humans.
CONCLUSION
Obesity leading to several complications during adulthood is a critical issue to be resolved and protected against during childhood.Metabolomics is a common tool for investigating not only adult but also children with obesity, whose data potentially provide clear fundamental mechanisms and evaluations.Nutrient factors, such as glucose, amino acids, and lipids, influence and relate with each other; nevertheless, amino acids are a critical modifier in obesity.Because amino acids reflect the internal metabolic change and intestinal microbiota and act as metabolic modulators, monitoring of amino acids by metabolome analysis may be a useful method to predict metabolic disorders during childhood.
ACKNOWLEDGEMENTS
We thank all the staff at our institution who cooperated in the collection of patient samples as well as those at the Department of Central Clinical Laboratory, Kumamoto University Hospital.
 World Journal of Meta-Analysis2021年2期
World Journal of Meta-Analysis2021年2期
- World Journal of Meta-Analysis的其它文章
- Helicobacter pylori infection and susceptibility to cardiac syndrome X: A systematic review and meta-analysis
- Laboratory hematologic features of COVID-19 associated liver injury:A systematic review
- Therapeutic applications of dental pulp stem cells in regenerating dental, periodontal and oral-related structures
- Nonalcoholic fatty liver disease and cardiovascular concerns: The time for hepatologist and cardiologist close collaboration
- Viral hepatitis: A brief introduction, review of management,advances and challenges
- Ankle injuries in athletes: A review of the literature