
HRAS基因表达对宫颈癌细胞自噬及凋亡作用及机制
2021-05-19 09:18阎臻付晓瑞李新敏崔珺
青岛大学学报(医学版) 2021年2期
阎臻,付晓瑞,李新敏,崔珺
(1 郑州市妇幼保健院妇产科,河南 郑州 450052; 2 郑州大学第一附属医院肿瘤科; 3 郑州市妇幼保健院病理科)
宫颈癌是与乳癌具有相似病死率的妇科恶性肿瘤,严重危害女性生活质量[1]。其目前可用治疗方式(外科手术为主、放化疗为辅)和治疗效果均值得肯定[2-3]。然而,上述疗法仍然存在一定的局限性,肿瘤复发率高、放化疗毒副作用大[4]。目前的研究重点已转向新型分子靶向药物的研发[5]。国内外相关研究认为,肿瘤发展由多因素共同作用,其中遗传学因素发挥重要作用[6-8]。在多种肿瘤中,RAS基因家族的基因突变使其激活,从而成为具有致癌活性的癌基因[9]。研究结果显示,Harvery鼠肉瘤病毒癌基因(HRAS)隶属于RAS基因家族,在胃癌、乳癌中异常表达[10-11]。同时,PTEN基因为多种肿瘤抑制基因,其适度表达可促进细胞凋亡,抑制细胞生长、浸润和转移[12-14]。本研究通过生物学预测网站发现HRAS和PTEN信号通路在宫颈癌中具有显著作用,本文探究HRAS基因表达对宫颈癌细胞自噬及凋亡的作用及其机制。
1 材料与方法
1.1 实验材料
人宫颈癌细胞株Hela细胞(中国科学院细胞库,上海);人宫颈癌标本(郑州市妇幼保健院手术切除标本);无特定病原体级雌性BALB/c小鼠(中国协和医科大学实验动物科学部,饲养于本院实验室);HRAS沉默质粒(si-HRAS)、HRAS过表达质粒(HRASvector)等质粒(上海吉凯基因化学技术有限公司,北京百奥莱博科技有限公司);PCR扩增引物(生工生物工程(上海)股份有限公司);实时定量聚合酶链反应(qRT-PCR)试剂盒(ABI公司,美国);ABI 7500荧光定量PCR仪(北京京科瑞达科技有限公司);RIPA细胞裂解液(上海图赫实业有限公司);Western blot检测抗体(上海博谷生物科技有限公司);BCA试剂盒(上海炎熙生物科技有限公司);ECL荧光检测试剂盒(苏州宇恒生物科技有限公司);Bio-Rad图像分析系统(伯乐生命医学产品(上海)有限公司);Annexin V-FITC/PI细胞凋亡检测试剂盒(碧云天生物技术研究所,江苏);流式细胞仪(BD公司,美国)。
1.2 实验方法
1.2.1宫颈癌小鼠模型的构建及组织处理 取BALB/c小鼠,每5只合并一笼饲养,12 h-12 h昼夜交替光照,能自由获取食物和水。采用数字表法将18只小鼠随机分成3组:正常组(A组),假手术组(B组),模型组(C组),每组6只小鼠。在无菌操作条件下,将人宫颈癌新鲜肿瘤标本经处理清除坏死组织和血块后,剪成约2 mm3的组织块备用。将模型组小鼠经乙醚吸入麻醉后固定于操作台,腹部消毒,做长0.5 cm切口,将制备标本植于腹部皮下。假手术组小鼠经腹腔注射1 mL纯生理盐水。手术全程在无菌条件下完成。正常组小鼠不做任何处理。于超净工作台打开小鼠腹腔,直接用肉眼观察荷瘤病灶形态。取模型组小鼠肿瘤组织分为两部分:一部分用于苏木精-伊红(HE)染色;一部分在液氮中速冻,然后移至-70 ℃保存,用于后续检测。
1.2.2细胞分组及转染 将宫颈癌Hela细胞分为4组,空白对照组(A组:不转染任何序列)、阴性对照组(B组:转染空载体质粒)、HRASvector组(C组:转染HRAS过表达质粒)、si-HRAS组(D组:转染HRAS沉默质粒siRNA)。将对数生长期Hela细胞接种于6孔板,细胞生长至30%~50%融合时行转染,室温孵育后加入细胞培养孔,于37 ℃、体积分数0.05的CO2条件下培养6~8 h后,换完全培养基,培养24~48 h后,进行后续实验。
1.2.3qRT-PCR检测 取各组小鼠组织标本以及各分组转染细胞,提取组织和细胞中总RNA。组织水平上检测HRAS和PTEN的表达量;细胞水平上检测HRAS、PTEN、ATG7、LC3、Beclin1、bax、Fas和E-cadherin的表达量。取反应液进行荧光定量PCR,采用ABI 7500荧光定量PCR仪检测。采用2-ΔΔCt计算mRNA表达量。实验重复3次。
1.2.4Western blot检测 转染各组细胞裂解后,离心弃除沉淀,取上清为细胞总蛋白。取上清液采用BCA试剂盒测定各样品的蛋白浓度。滴加一抗兔多克隆抗体HRAS、PTEN、ATG7、LC3、Beclin1、bcl-2、bax、Fas和E-cadherin,4 ℃孵育过夜,次日加HRP标记山羊抗兔IgG二抗室温下孵育。以化学发光凝胶成像仪进行曝光。实验重复3次。
1.2.5流式细胞术检查 采用Annexin V-FITC/PI细胞凋亡检测试剂盒检测各组细胞染色后凋亡变化。将各组细胞置37 ℃、体积分数0.05的CO2培养箱中培养48 h,收集细胞,PBS洗涤后将细胞重悬,加入Annexin V-FITC和PI轻轻混匀,避光室温反应15 min。使用流式细胞仪在激发波长488 nm处检测细胞凋亡水平。实验重复3次。
1.3 统计学方法
2 结 果
2.1 宫颈癌小鼠模型构建
建模21 d后,小鼠全部成活,模型组小鼠均荷瘤成功,成瘤率为100%。肉眼观察和HE染色结果显示,正常组和假手术组小鼠行动和外观相对正常、无体质量的明显变化,均未见肿瘤病灶的形成,组织切面无异常变化,细胞结构完整;模型组小鼠精神萎靡、行动迟缓、体质量明显下降,有半球形伴破溃和出血的肿瘤病灶形成,组织切面灰白、质硬、血管丰富,癌细胞呈巢状、胞核增大、核仁不明显,表明建模成功。见图1。
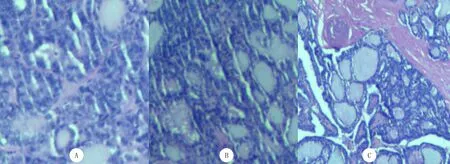
A:正常组,B:假手术组C:模型组。HE染色,200倍。
2.2 组织HRAS和PTEN的mRNA和蛋白表达
qRT-PCR和Western blot检测结果显示,正常组和假手术组小鼠组织中的HRAS和PTEN的mRNA和蛋白表达量均无明显差异(t=1.180~1.821,P>0.05)。与正常组相比较,模型组小鼠组织中HRASmRNA和蛋白的表达量均显著增加,PTENmRNA和蛋白的表达量下降(t=9.450~14.260,P<0.05)。见图2。
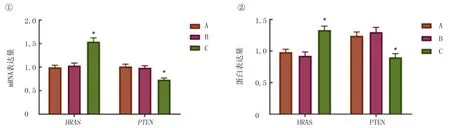
①qRT-PCR检测;②Western blot检测。A:正常组,B:假手术组,C:模型组。与正常组小鼠组织相比,*P<0.05。
2.3 HRAS基因表达对PTEN信号通路的影响
qRT-PCR和Western blot检测结果显示,空白对照组HRASmRNA和蛋白表达与阴性对照组比较差异无显著性(t=0.186~1.088;均P>0.05);与空白对照组和阴性对照组比较,HRASvector组HRAS的mRNA和蛋白表达水平明显上升(F=19.600、23.270,P<0.05),PTEN的表达水平则明显下降(F=16.760、35.130,P<0.05)。si-HRAS组HRAS的mRNA以及蛋白表达水平明显下降(F=8.127、13.160,P<0.05),PTEN的表达水平明显上升(F=9.456、22.940,P<0.05),差异均有统计学意义。见图3。
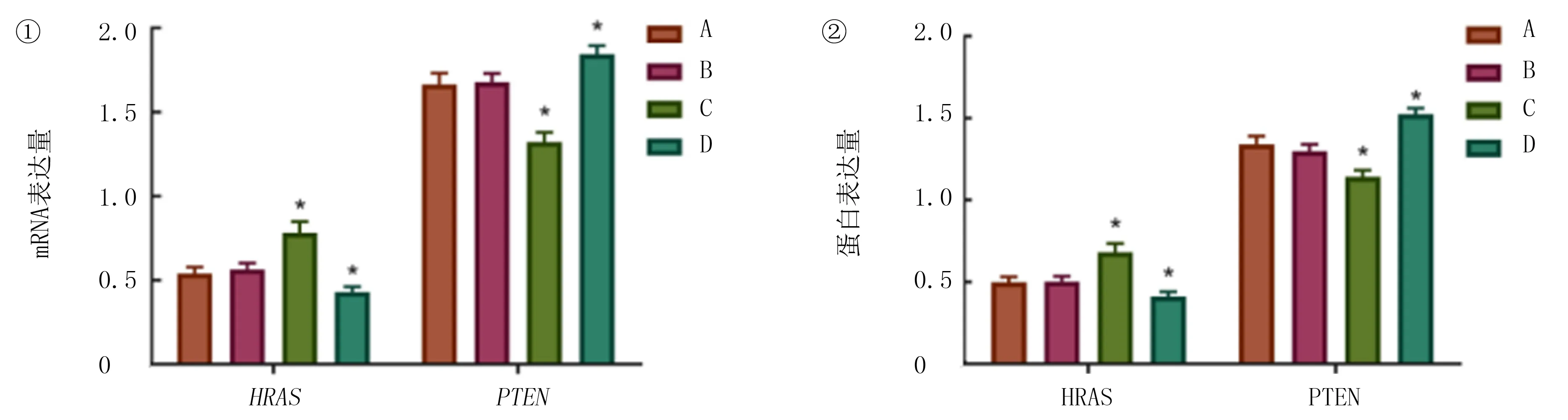
①qRT-PCR检测;②Western blot检测。A:空白对照组,B:阴性对照组,C:HRASvector组,D:si-HRAS组。与空白对照组和阴性对照组相比,*P<0.05。
2.4 HRAS基因表达对上皮间质转化的影响
qRT-PCR和Western blot检测上皮间质转化因子E-cadherin表达的结果显示,E-cadherin的mRNA和蛋白表达在空白对照组与阴性对照组间差异无统计学意义(t=0.069、0.686,P>0.05);与空白对照组和阴性对照组比较,HRASvector组E-cadherin mRNA和蛋白表达水平明显下降(F=4.220、8.110,P<0.05)。si-HRAS组E-cadherin蛋白表达水平明显上升(F=12.600、18.280,P<0.05)。见图4。
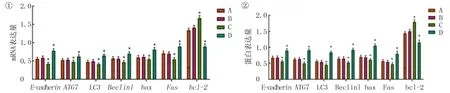
①qRT-PCR检测;②Western blot检测。A:空白对照组,B:阴性对照组,C:HRASvector组,D:si-HRAS组。与空白对照组和阴性对照组相比,*P<0.05。
2.5 HRAS基因表达对细胞自噬的影响
qRT-PCR和Western blot检测细胞自噬相关因子ATG7、LC3、Beclin1表达结果显示,空白对照组与阴性对照组间差异无统计学意义(t=0.130~0.470,P>0.05);与空白对照组和阴性对照组比较,HRASvector组ATG7、LC3、Beclin1 mRNA和蛋白表达均明显下降(F=2.742~7.310,P<0.05)。si-HRAS组ATG7、LC3、Beclin1 mRNA和蛋白表达明显上升(F=3.160~30.970,P<0.05)。见图4。
2.6 HRAS基因表达对细胞凋亡相关因子和凋亡率的影响
qRT-PCR和Western blot检测bax、Fas、bcl-2结果显示,空白对照组和阴性对照组比较差异均无统计学意义(t=0.245~1.459,P>0.05);与空白对照组和阴性对照组相比较,HRASvector组bax和FasmRNA及蛋白表达水平明显下降(F=1.011~7.314,P<0.05),bcl-2mRNA和蛋白表达水平则明显上升(F=177.300、220.400,P<0.05)。si-HRAS组bax及FasmRNA和蛋白表达水平明显上升(F=11.200~41.180,P<0.05),bcl-2mRNA和蛋白表达则明显下降(F=113.600、129.800,P<0.05)。流式细胞术的检测结果表明,空白对照组和阴性对照组比较差异均无统计学意义(t=2.020,P>0.05)。与空白对照组和阴性对照组相比较,HRASvector组细胞凋亡率明显下降,差异有统计学意义(F=31.020,P<0.05);si-HRAS组细胞凋亡则明显增加,差异具有统计学意义(F=622.800,P<0.05)。见图4、5。
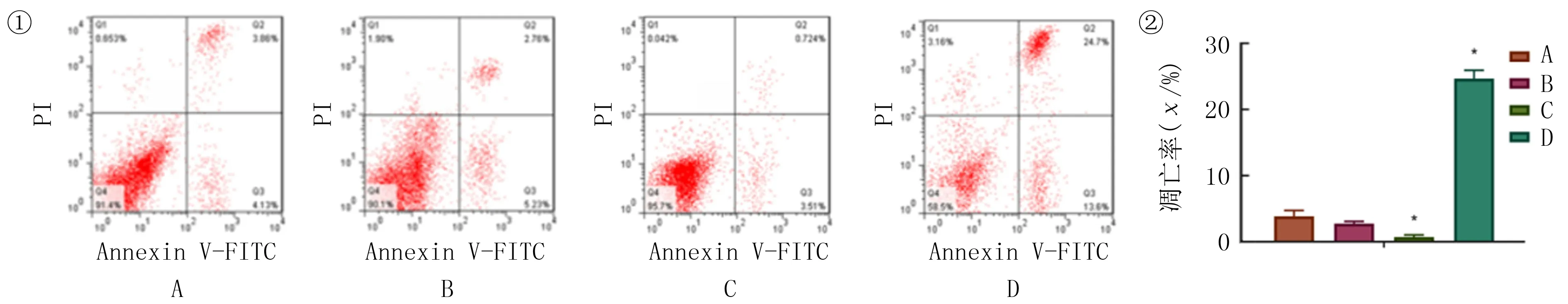
①细胞转染后各组细胞凋亡图;②统计分析图。A:空白对照组,B:阴性对照组,C:HRASvector组,D:si-HRAS组。与空白对照组和阴性对照组相比,*P<0.05。
3 讨 论
基因治疗通过基因上调或沉默,介导特定作用通路,发挥对人类肿瘤的治疗作用[15]。宫颈癌为女性常见妇科肿瘤之一,其发病率呈年轻化趋势[16]。人类乳头状瘤病毒(HPV)感染已被证实为宫颈癌的主要诱因,且既往研究已证实HPV感染可通过刺激或抑制一系列复杂信号通路,进而诱导宫颈上皮细胞的瘤变或癌变[17-18]。上皮间质转化是人类恶性肿瘤发生转移的主要原因之一,其可改变肿瘤细胞特性或肿瘤细胞微环境,促进肿瘤生长、诱导癌细胞增殖和转移[19-20]。PTEN是最早发现的抑癌基因之一,已被证实在人类诸多癌症中发生基因突变或高频缺失,如生殖系统肿瘤、消化道肿瘤、呼吸系统肿瘤等[21-22]。PTEN可以调控多条信号通路,如PIK/AKT通路,诱导细胞凋亡并抑制细胞侵袭转移。例如,SONG等[23]就曾报道microRNA-126可通过靶向PIK3R2基因,进而调控PTEN/PI3K/AKT信号转导途径抑制非小细胞肺癌A549细胞的增殖、迁移和侵袭。国内亦有相似研究报道,如蒋立峰等[24]报道肺癌病人外周血Treg细胞计数明显升高,且其机制可能与PTEN表达下调激活PI3K-AKT信号通路相关;孙吉春等[25]发现抑制MiR-21表达可能通过调控PTEN/AKT信号通路抑制胰腺癌细胞AS-PC-1增殖和侵袭。目前,通过靶向激活PTEN信号通路从而促进上皮间质转化、促进细胞增殖侵袭、自噬和凋亡等细胞生物学行为,已成为肿瘤基因治疗的一新型热点方向。
RNA干扰是通过关闭特定基因序列表达从而实现目标基因表达沉默的过程。基因沉默在基因功能研究以及关联信号通路的研究中发挥着重要作用,可用于探究肿瘤生长机制,为肿瘤的基因治疗提供潜在的治疗途径[26]。鉴于此,本研究初步推测:HRAS基因沉默可能通过介导PTEN信号通路参与宫颈癌发病过程。
本文HE染色结果显示,宫颈癌小鼠造模成功。造模后HRAS在宫颈癌小鼠中呈高表达,PTEN表达被抑制,过表达HRAS基因抑制PTEN表达,表明上调HRAS基因可能抑制PTEN信号通路的激活。同时,结合PCR、Western blot检测细胞上皮间质转化、自噬和凋亡相关因子及凋亡率的结果表明,上调HRAS表达可导致bcl-2mRNA和蛋白表达水平明显上升,PTEN、ATG7、LC3、Beclin1、bax、Fas和E-cadherin呈低表达,细胞凋亡率明显下降;与此同时,低表达HRAS基因可激活PTEN信号通路活性,导致PTEN表达的上升,并下调bcl-2的表达,上调ATG7、LC3、Beclin1、bax、Fas和E-cadherin的表达,促进细胞凋亡。上述研究结果说明,HRAS基因沉默可诱导PTEN信号通路的激活,促进上皮间质转化,进而促进细胞自噬和细胞凋亡。ATG7、LC3、Beclin1是细胞自噬的关键蛋白[27];bax和Fas为促凋亡蛋白,而bcl-2为抑制凋亡蛋白[28]。细胞自噬为继细胞凋亡的另一种细胞程序性死亡,其和细胞凋亡均在肿瘤细胞生存中扮演重要角色;上皮间质转化同样对肿瘤细胞转移意义重大。作者推测外源性HRAS基因沉默通过促进PTEN信号通路的激活,发挥PTEN抑制细胞增殖分裂、肿瘤浸润转移及促肿瘤细胞凋亡效应;诱导细胞自噬相关蛋白ATG7、LC3和Beclin1表达,促凋亡蛋白bax和Fas表达、上皮间质转化特征性蛋白E-cadherin表达,并下调抑制凋亡蛋白bcl-2表达,提示上皮间质转化、细胞自噬和凋亡均参与了宫颈癌细胞死亡过程。本实验结果符合既往研究有关HRAS基因表达和PTEN信号通路作用的报道。例如,SUN等[29]发现,HRAS异常甲基化诱导该促癌基因的激活可能是膀胱肿瘤发生的早期事件,并可能进一步作为膀胱组织肿瘤生物标志物用于早期诊断和作为潜在的治疗靶点。WU等[30]对吉西他滨在乳癌中的耐药性进行研究,结果提示miR-21可通过PTEN/AKT信号通路诱导上皮间质转化,其结果可用于预测最佳乳癌治疗并为逆转吉西他滨耐药性提供潜在治疗靶点。
总而言之,本研究显示HRAS基因表达下调可促进PTEN信号通路的激活,促进上皮间质转化、小鼠宫颈癌细胞自噬和凋亡,从而抑制宫颈癌的发生。本研究结果有助于深入探究宫颈癌发生相关分子机制,从而为PTEN作为预防和改善宫颈癌治疗方案潜在功能分子提供理论依据。然而,本文尚存在部分问题有待进一步研究,例如下调HRAS基因表达如何激活PTEN信号通路、上皮间质转化及自噬和凋亡等,HRAS基因表达下调介导PTEN信号通路激活在动物模型中的作用验证,等等。这些问题均有助于我们更充分地理解HRAS基因表达通过PTEN信号通路介导宫颈癌发生的作用机制,为该肿瘤的治疗提供充分的理论证据。
猜你喜欢
新农业(2022年21期)2022-11-18
中日友好医院学报(2022年4期)2022-10-15
昆明医科大学学报(2022年3期)2022-04-19
中老年保健(2021年12期)2021-08-24
现代临床医学(2021年4期)2021-07-31
云南医药(2021年3期)2021-07-21
天津医科大学学报(2021年1期)2021-01-26
中国生殖健康(2020年7期)2021-01-18
云南医药(2020年5期)2020-10-27
看世界·学术下半月(2020年7期)2020-09-10
