
藏酋猴与人类腰椎椎体骨密度对比研究
2021-05-11 03:29:18牟彦郦宗孝容张宇航雍刘军
局解手术学杂志 2021年4期
牟彦郦,宗孝容,张宇航,雍刘军
(1.成都医学院2017级临床医学本科,四川 成都 610500;2.成都医学院人体解剖与组织胚胎学教研室/发育与再生四川省重点实验室,四川 成都 610500)
藏酋猴属于灵长目,猴科,猕猴属[1],为中国特有非人灵长类杂食性哺乳动物,其在灵长类的进化轨迹上与人类的关系仅次于类人猿,在形态结构、生理、代谢功能和生物学等方面与人类高度相似[2-3],是一种潜在的、极具生物医学研究价值的非人灵长类动物[4]。藏酋猴主要以槐树、狗枣子等植物为食[5],喜欢在地面活动,在崖壁缝隙、树枝间等阳光充裕的地方穿梭跳跃,这种肌肉收缩及舒张运动的机械负荷,使其生长期骨钙盐量增加[4-6],可作为骨形态、结构及力学研究的理想实验对象。
骨质疾病是一个发病率高、涉及人群广、致病危险因素复杂、后果严重的普遍存在的健康问题[7]。骨质疾病的发生与骨密度密切相关[8-9],骨密度是机体某一部位组织中标准骨面积内的矿物质含量[10],受饮食习惯、生活方式、运动强度、遗传、地域等多种因素的影响[11]。人类腰椎椎体富含骨松质,导致骨质病变多发,尤其是骨质增生和骨质疏松,故腰椎椎体成为测量骨密度的常用部位[12]。本课题组在研究中发现,藏酋猴腰椎富含较厚骨密质,未发现人类常见病变,可作为人类腰椎椎体疾病研究的理想动物模型,具有巨大的研究价值。
目前对藏酋猴社会关系与行为的研究甚多,但对其椎体骨密度的研究较少。本研究通过观察藏酋猴与人类腰椎椎体的骨密质和骨松质,测量并比较骨密度数值,探讨两者腰椎椎体骨密度的差异、相关性及差异形成的原因,以期为人类腰椎骨质病变的研究提供参考。
1 材料与方法
1.1 实验材料
选取10~15岁健康成年藏酋猴腰椎椎体标本6具,其中雄、雌各3具,由四川省医学科学院动物研究所提供;选择40~60岁(与藏酋猴年龄相当,藏酋猴1岁相当于人类3~4岁)健康成人腰椎椎体标本12具,其中男、女各6具,由成都医学院人体解剖实验室提供。藏酋猴腰椎有7节,人类腰椎有5节,藏酋猴的第6、7腰椎相当于人类的第1、2骶椎,为与人类腰椎相对应,藏酋猴与人类均选用L1~L5进行研究。由于藏酋猴标本数量限制,本研究对象不区分性别。
1.2 实验方法
将6具健康成年藏酋猴及12具健康成年人的腰椎椎体标本以4%多聚甲醛灌注固定2周后,解剖分离腰椎椎体,并行正中冠状切、矢状切和水平切。用手术显微镜(XT-X-4A)观察各切面骨松质、骨密质的分布和骨小梁的排列;数显游标卡尺(单位:mm,精确度:0.01)测量各切面骨松质及骨密质厚度;并采用高精度电子密度计(型号:DK-300A,精度:0.001 g/cm3),通过其溶液补偿功能,以硅油为介质测量各腰椎椎体骨密度。
1.3 统计学方法
采用SPSS 21.0统计软件处理数据。藏酋猴与人类各腰椎椎体骨密度采用独立样本t检验,P<0.05为差异有统计学意义。藏酋猴与人类腰椎椎体骨密度采取Pearson相关性分析(P<0.01,r∈[-1,1]),P<0.01说明两变量存在显著相关关系。
2 结果
2.1 藏酋猴腰椎各切面观察
藏酋猴腰椎椎体各切面骨松质所占面积均大于骨密质。正中冠状面骨松质周围为一层较厚的骨密质,骨小梁较厚且排列紧密、方向杂乱,骨松质的腔隙在中部较大,边缘较小且密集(图1a)。正中矢状面骨松质表面有一层较薄的骨密质,骨松质中骨小梁排列方向较为一致,中部骨小梁数量较周围更少,排列更疏松,骨松质中部的腔隙较大,边缘的腔隙较小且密集(图1b)。正中水平面骨松质周围为一层极厚的骨密质,骨松质中骨小梁较厚且分布较均匀,排列方向杂乱,骨松质的腔隙普遍较小且密集,周围骨密质厚度不均匀,腹侧骨密质最薄、背侧骨密质较厚(图1c)。
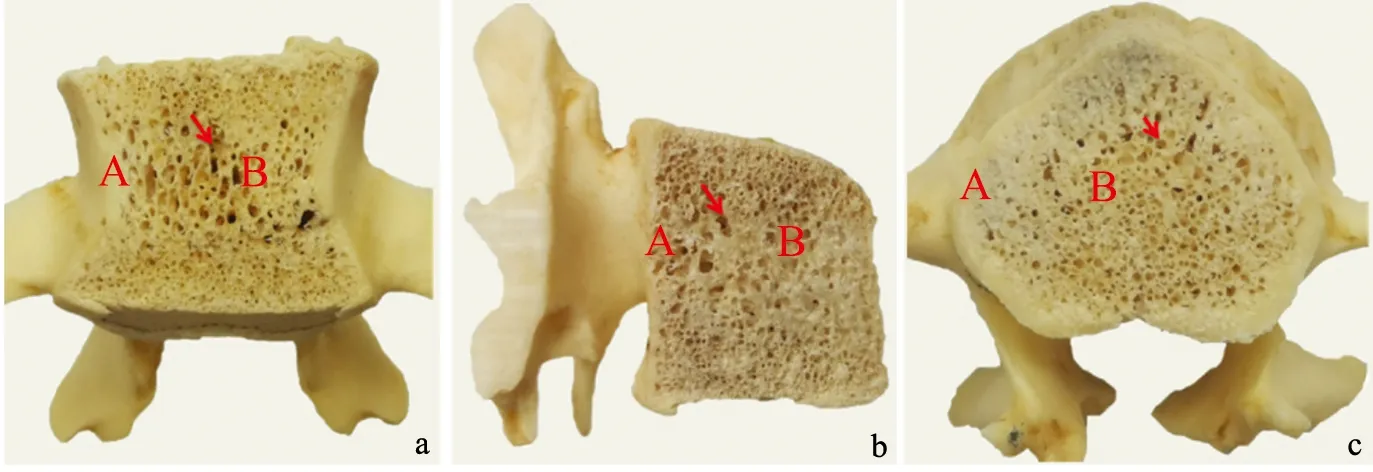
a:冠状切面观;b:矢状切面观;c:水平切面观 A:骨密质;B:骨松质;箭头:骨小梁
2.2 人类腰椎各切面观察
人类腰椎椎体3个切面均以骨松质为主,周边附着一层极薄的骨密质。正中冠状面骨小梁排列较为规则,以纵向为主,骨松质的腔隙在中部较大,边缘较小且密集(图2a)。正中矢状面骨小梁分布均匀,排列规律,水平中部以纵向为主,腔隙较大,排列疏松,边缘以横向为主,排列较中部密集(图2b)。正中水平面骨小梁呈放射状不规则分布,排列逐渐疏松,骨松质的腔隙普遍较大且疏松,周围骨密质厚度不均匀,腹侧骨密质最薄、背侧骨密质较厚(图2c)。

a:冠状切面观;b:矢状切面观;c:水平切面观 A:骨密质;B:骨松质;箭头:骨小梁
2.3 藏酋猴和人类骨密度比较
藏酋猴腰椎正位L1~L5骨密度范围为0.69~0.92 g/cm3,呈逐渐下降趋势,中位数为0.81 g/cm3,均数为0.78 g/cm3,方差为0.050,标准误为0.019,波动范围较小;其腰椎骨密度最大值为L1的(0.89±0.03)g/cm3,最小值为L5的(0.71±0.03)g/cm3。人类腰椎正位L1~L5骨密度范围为0.61~0.81 g/cm3,呈逐渐上升趋势,中位数为0.71 g/cm3,均数为0.66 g/cm3,方差为0.001,标准误为0.008,波动范围较小;其腰椎椎体骨密度最大值为L4的(0.70±0.02)g/cm3,最小值为L1的(0.62±0.01)g/cm3。其中两者L1骨密度差异最大,为(0.31±0.02)g/cm3,L4差异最小,为(0.05±0.01)g/cm3,且藏酋猴的波动范围较人类大,在一定程度上说明藏酋猴腰椎椎体骨密度大于人类。
对藏酋猴和人类L1~L5椎体骨密度分别进行t检验,藏酋猴L1~L3椎体骨密度与人类比较,差异均具有统计学意义(P<0.05);藏酋猴L4、L5椎体骨密度与人类比较,差异无统计学意义(P>0.05),见表1。Pearson相关性分析表明,藏酋猴腰椎椎体骨密度与人类呈显著负相关(r=-0.963,P<0.01)。
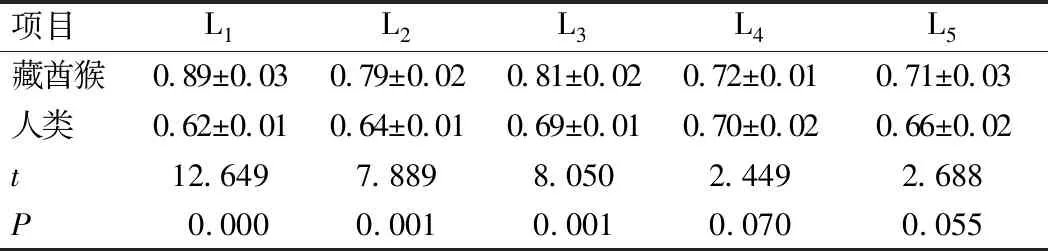
表1 藏酋猴与人类L1~L5椎体骨密度(g/cm3)
3 讨论
本研究中藏酋猴腰椎各切面骨松质所占面积均大于骨密质,周围附着一层较厚的骨密质,这可能与其饮食结构有关。藏酋猴以植物性食物为主,宋蔚[13]的研究表明,植物性食物中含有大量维生素D和植物雌二醇,其中维生素D可促进骨密度的增加,植物雌二醇则是天然的植物化合物,有微弱的雌激素作用,可预防骨质流失。藏酋猴独特的腰椎骨质分布还可能和生活环境有关,其活动位点海拔最低1 120 m,最高2 240 m,主要集中在1 300~1 500 m及1 500~2 000 m 2个高海拔段范围内[5]。随着海拔的升高,光照需要穿过的距离缩短,受大气削弱量也减少,因此光照强度增强[14],这种特殊的生活环境,使藏酋猴能够获得充足的光照,合成大量维生素D3[15],利于骨量的维持,形成紧密排列的骨质。藏酋猴腰椎各切面骨小梁的排列与其运动方式有着一定的相关性[16]。藏酋猴善于穿梭跳跃式冲击负荷运动,这种长时间的肌肉收缩及舒张运动促使其生长期骨钙盐量增加,可抑制破骨细胞骨吸收[17],减少骨质的损失,使得骨小梁较厚、排列紧密,骨松质的腔隙普遍较小且密集。又因藏酋猴可直立行走,椎体受力方向多变,导致骨小梁在正中冠状面、水平面方向杂乱,正中矢状面排列较为一致。
人类骨密度与性别、年龄(女性绝经年限)存在一定的相关性[18],但经定量CT测量腰椎椎体骨密度显示,骨密度受性别、年龄、BMI等的影响较小[12],故本研究不对性别、年龄、BMI等因素进行分析。本研究结果显示,人类腰椎各切面均以骨松质为主,周边附着一层极薄的骨密质。这可能与人类以肉食为主的饮食习惯[19]以及运动频率较小、运动量不足、运动形式单一的生活方式有关。肉制品会产生酸性物质[13],破坏机体的酸碱平衡,故造成其骨含量降低,骨密质含量较少。人类以直立行走和端坐的运动方式为主,其受力主要集中在纵轴方向,从而使得正中冠状面、矢状面骨小梁分布均匀、排列规则,以纵向为主;而四肢着地的运动形式在人类运动方式中少见,造成正中水平面骨小梁呈放射状不规则分布。
藏酋猴腰椎椎体骨密度普遍大于人类,可能与其跳跃式冲击负荷运动有关。藏酋猴以四肢着地的运动为主,脊柱类似于杠杆,同时又受腹腔内容物牵拉作用的影响[20],受力主要集中于脊柱的L1背侧,向下至L5受力逐渐减小,使得腹侧骨密质最薄、背侧骨密质较厚,骨密度呈逐渐下降趋势,是藏酋猴与人类腰椎椎体骨密度呈显著负相关的原因之一。此外,人类直立行走和端坐的运动方式,使得L1~L5椎体受力逐渐增大,是藏酋猴与人类腰椎椎体骨密度呈显著负相关的另一原因。
人类骨质疾病的发生与骨密度存在密切相关性[7-8],骨密度与饮食习惯、运动方式和运动强度存在一定的相关性,科学合理饮食和运动能够有效提高骨密度[6,13]。宋蔚[13]建议,食用富含植物雌二醇的弱碱性食物,如谷物、水果、蔬菜,减少肉制品的摄入,可降低骨质流失,维持骨量,预防骨质疏松[13]。适当地丰富运动形式,保持一定的运动量,可抑制骨吸收和骨重建反应,提高骨量峰值[21],从而增加骨密度。因此,植物性食物和适当的运动,可增加骨密度,抑制骨质疏松等病变的发生。
综上所述,藏酋猴腰椎椎体骨质排列较人类更加紧密,骨密质远厚于人类,其骨密度普遍大于人类,与两者饮食结构、生活方式及生活环境有关。
猜你喜欢
中老年保健(2022年3期)2022-08-24 03:00:58
护理研究(2022年14期)2022-08-06 08:59:48
保健医苑(2021年7期)2021-08-13 08:48:02
中华养生保健(2020年9期)2021-01-18 03:11:50
音乐教育与创作(2020年9期)2021-01-04 01:01:30
中国生殖健康(2019年2期)2019-08-23 08:12:02
中国临床医学影像杂志(2019年4期)2019-06-18 10:54:50
益寿宝典(2018年32期)2018-01-26 19:36:36
今古传奇·故事版(2017年23期)2018-01-12 20:20:13
中国卫生标准管理(2015年10期)2016-01-15 00:48:56
